米糠的营养价值及酸败机制研究进展
2021-04-13高亚楠石宝明赵倩倩
高亚楠 石宝明 赵倩倩 方 静
(东北农业大学动物营养研究所,哈尔滨150030)
水稻是全球第二大粮食作物,2019年我国水稻总产量达到20 961万t[1]。米糠是稻谷加工过程中的副产物,占稻谷总重量的5%~7%,由外果皮、中果皮、交朕层、种皮及糊粉层组成,集中了64%的稻米营养素及90%以上的人体必需元素[2],包括丰富的脂质、蛋白质、膳食纤维以及谷维素、植物甾醇、生育酚等抗氧化物质[3]。稻谷原料和加工技术的差异直接导致米糠中各种成分的含量变化,米糠根据其加工方式可分为普通米糠、脱脂米糠和细米糠。米糠是一种很好的能量饲料,但因其油脂含量较高,在米糠高活性脂解酶的作用下产生大量挥发性脂肪酸,这些产物又在脂氧合酶(lipoxygenases,LOX)的作用下发生氧化酸败变质,限制了其在饲料生产中的应用。为提高对米糠营养价值、酸败机制及解决方式的系统认知,本文结合国内外相关研究资料,从米糠的营养价值、米糠酸败危害、脂解酶和LOX的作用机制、国内外稳定米糠的方法等方面进行综述,为米糠在饲料工业中的高效利用提供一定的理论依据。
1 米糠的营养价值
1.1 米糠中营养成分及限制因素
米糠富含粗蛋白质、粗脂肪、维生素E等营养物质,相比较玉米,氨基酸构成更合理,米糠脱脂后的蛋白质含量高达15.1%[4],是一种丰富而价廉的植物蛋白质资源。米糠蛋白中约70%为可溶性蛋白质,与大豆蛋白相似,必需氨基酸种类齐全,具有低过敏性及良好的溶解性,蛋白质效率比为2.0~2.5,蛋白质消化率大于90%[5]。经胃肠道消化的米糠蛋白可产生抗氧化肽和游离多酚,是一种潜在的抗氧化剂来源[6]。但是,米糠在脂解酶和LOX的作用下产生的脂质氢过氧化物可诱导米糠蛋白氧化,影响营养价值[7]。米糠油中含有35%~45%的油酸、30%~35%的亚油酸、1%~2%的亚麻酸以及单不饱和脂肪酸、多不饱和脂肪酸、棕榈酸、饱和脂肪酸等,其中油酸、亚油酸含量均比玉米高2~6倍[8],然而米糠油完全利用受到游离脂肪酸大量积累的影响(约占总油脂的80%),这归因于米糠中脂解酶的存在;米糠油中不皂化物含量(3.0%~4.5%)相比较其他普通植物油(1%)具有独特优势,不皂化物含有独特的天然抗氧化物复合物,如植物甾醇(1.8%)、γ-谷维素(1.6%)和生育酚等[9]。但是在传统精炼过程中,谷维素、甾醇会随着米糠油加工所得的副产物皂脚被带走[10]。米糠含有200~350 g/kg的膳食纤维,主要是由粗纤维素、木质素和果胶组成,在调节宿主代谢、免疫系统和细胞增殖中起着重要作用[11-12]。但是米糠中可溶性膳食纤维只占米糠膳食纤维的1%~2%,不溶性膳食纤维难以被单胃动物内源酶消化吸收。米糠含有丰富的B族维生素及维生素E,但维生素A、维生素D、维生素C含量则较少;钙磷比例不平衡,磷含量较高,但属于植酸磷,锰、钾、镁、硅含量也较多;米糠还含有抗胰蛋白酶抑制因子、糜蛋白抑制剂、植酸、过氧化酶和脂解酶等抗营养因子[13]。
1.2 米糠在动物饲粮中的应用
玉米和豆粕是提供能量和蛋白质的主要原料,然而玉米和豆粕价格每年都在波动,为了经济上的优势,非常规饲料原料已被用来替代传统饲料原料。米糠是较好的能量饲料,营养水平见表1,米糠中粗蛋白质含量比麸皮低但比玉米高;必需氨基酸中赖氨酸、蛋氨酸含量均高于玉米和麸皮;粗脂肪含量比玉米、麸皮都高;粗纤维含量低于小麦麸但高于玉米,且价格低于玉米和小麦麸,在配合饲料中可以部分或大部分替代玉米使用。

表1 米糠营养水平
1.2.1 米糠在猪饲粮中的应用
氨基酸是猪新陈代谢的重要营养物质,Lee等[14]报道米糠中粗蛋白质、赖氨酸、蛋氨酸的标准化回肠消化率分别为74.4%、78.5%、85.3%,而且米糠中的粗脂肪含量可以提高猪对氨基酸的消化率。适宜的饲粮粗纤维水平不会降低猪的生长性能,有的甚至有提高作用。在猪小肠中,饲粮中粗纤维不能被内源性酶消化,在后段肠道被微生物发酵产生短链脂肪酸,不仅为机体供能,还使肠道pH降低[15]。这个过程促进了肠道有益菌增殖成为优势菌,抑制大肠杆菌等有害菌增殖[16]。伴随后段肠道微生物的快速生长繁殖,其对氮的需求增加,从而减少猪氮排放[17]。郝帅帅[18]用34.8%的米糠替代部分玉米饲喂苏淮猪的研究发现,米糠对日增重、屠宰性能和肉品质指标等与对照组均无显著差异。龙际飞等[19]以48头宁乡猪为试验对象,发现饲喂在肥育前期米糠替代量为6.00%、中期为5.50%、后期为7.00%的碎米-米糠型饲粮,对胴体品质和肉品质未产生不良影响。添加34.8%米糠的高粗纤维水平饲粮还可提高苏淮猪的大肠占整个肠道质量的比重,同时有助于苏淮猪的十二指肠和空肠绒毛生长,加深空肠隐窝深度[20]。Pu等[21]报道,当饲喂膳食纤维水平为19.10%的脱脂米糠时,与对照组相比表观消化率没有显著变化,但是增加了远端肠道微生物群的多样性和代谢能力,促进粮纤维降解菌数量的增加,提高了挥发性脂肪酸的含量。Herfel等[22]在断奶仔猪饲粮添加10%稳定化米糠可提高仔猪饲料转化率,还伴随着结肠双歧杆菌数量的增加。Fan等[23]研究表明,饲粮中用脱脂米糠替代玉米降低了血清中的炎症生物标记物含量;并通过上调结肠中黏蛋白2(MUC2)基因表达水平,下调结肠中核因子E2相关因子2(Nrf2)、醌烟酰胺腺嘌呤二核苷酸磷酸脱氢酶1(NQO1)和血红素氧合酶1(HO-1)基因表达水平显示出调节肠屏障的潜在功能;但是,添加脱脂米糠水平与肠道健康之间并不总是呈正相关。高芳芳等[24]在脱脂米糠等量替代饲粮中部分玉米的研究中发现,饲喂含28%脱脂米糠饲粮可提高苏淮猪血清和肝脏中抗氧化酶活性。
1.2.2 米糠在家禽饲粮中的应用
米糠中植酸盐与结构性碳水化合物会影响家禽对矿物质和粗蛋白质的消化吸收,在家禽饲粮中用米糠替代玉米的饲养效果会随着米糠替代水平的增加而降低,但可通过添加复合酶制剂的方法提高其利用率。米糠油中的不饱和脂肪酸和生物活性物质有改善家禽生长性能、免疫应答等功能的作用。Sun等[25]研究发现,饲喂28日龄四川鹅含18%全脂米糠的饲粮会抑制28~42日龄鹅的采食量,但在生长后期出现补偿性生长,对57~70日龄四川鹅生长性能有积极影响;此外,饲喂含18%全脂米糠的饲粮可降低鹅肉皮下脂肪含量,增加多不饱和脂肪酸含量,提高肉质营养价值。试验表明,当饲粮脱脂米糠水平超过20%就会影响扬州鹅生长性能、屠宰性能和内脏发育[26]。Gallardo等[27]的研究发现,在30%全脂米糠饲粮中添加复合多糖酶(200 mg/kg)和植酸酶(50 mg/kg)时,肉鸡对氮、能量和氨基酸的利用效率会得到提升。Kang等[28]在雏鸡基础饲粮分别添加不同水平(5、10、20 g/kg)米糠油的研究发现,随着米糠油添加水平的增加,雏鸡的日增重(线性和二次曲线)、饲料转化率(线性)、血清中免疫球蛋白G(lgG)含量(线性)显著提高,胆固醇含量(线性)显著降低。
1.2.3 米糠在反刍动物饲粮中的应用
在反刍动物饲粮中加入脂肪可提高能量利用效率,还可防止瘤胃酸中毒,促进脂溶性营养的吸收。γ-谷维素是米糠油中主要的生物活性物质,已经证明对动物健康有许多益处。赵倩明[29]采用瘤胃尼龙袋法和饲养试验相结合,分析了在泌乳奶牛饲粮中添加玉米胚芽粕和全脂米糠对奶牛生长性能的影响,结果显示在泌乳中期饲粮中加入6.80%玉米胚芽粕和3.40%米糠,有利于降低饲料成本,提高奶牛产奶量。Vargas等[30]发现饲粮中添加22.5%的全脂米糠对羔羊干物质和养分的表观消化率、氮平衡、瘤胃液氨氮浓度和pH均无显著影响。在评估澳大利亚初生羔羊对添加多不饱和脂肪酸强化颗粒饲料反应的研究中发现,添加米糠油可改善羔羊的生长性能和胴体品质[31]。Bhatt等[32]研究发现,在马尔普拉羔羊的精料中添加40 g/kg钙皂形式米糠油(脂肪酸钙盐)可提高断奶羔羊的胴体性状。Escobar等[33]在每日补充33 mg/BW0.75的γ-谷维素对公羊阴囊隔离引起的睾丸变性的研究中发现,米糠中的γ-谷维素可通过降低氧化应激保护精液和睾丸,但是补充γ-谷维素也会导致精子有更多的异常形态。
2 米糠酸败危害
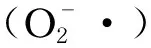
3 造成米糠酸败的酶类
3.1 脂解酶
脂解酶是一种能够催化酯类物质水解合成物质的总称,主要包括酯酶(EC3.1.1.1)和脂肪酶(EC3.1.1.3)[39],尽管二者序列差异很大,但大多数都含有1个以丝氨酸(Ser)为活性位点的保守催化三联体[Ser-组氨酸(His)-天冬氨酸(Asp)/谷氨酸(Glu)],都具有1个特殊的序列-G X1S X2G-(其中X是任何氨基酸)[40]。根据形成含氧阴离子洞(能稳定反应中间体的活性位点的氨基酸)和催化三联体的结构和残基,微生物脂肪酶、酯酶和相关蛋白质可分为38个超家族和112个同源家族[41]。酯酶催化小分子酯类,而脂肪酶催化三酰基甘油脂水解为甘油和脂肪酸;酯酶倾向于水解少于10个碳的短链脂肪酸的酯键,而脂肪酶水解多于10个碳的长链脂肪酸酯键;酯酶能够水解水溶性底物,而脂肪酶能够水解油水界面上的底物;脂肪酶具有界面活性而酯酶没有[42-43]。
米糠中除了脂解酶以外还含有3种类型的磷脂酶,分别是磷脂酶A(A1和A2)以及磷脂酶C、磷脂酶D。这4种类型酶的相对活性比为100∶24∶35∶39,磷脂酶作用于磷脂单分子层,是种子储存脂质萌发动员的关键酶[44]。脂肪酶-Ⅱ是活性最高的脂解酶,它是一种碱性蛋白质,催化三联体为Ser175-Asp231-His292[45-47],近些年报道的米糠中的脂解酶见表2。

表2 米糠中的脂解酶
3.2 LOX
LOX是不饱和脂肪酸代谢的关键酶类,可将花生四烯酸、亚油酸及其他多不饱和脂肪酸转变为有生物活性的代谢产物,从而影响细胞结构、代谢及信号转导[52]。稻谷中LOX主要存在种胚和种皮中,含量很低。稻谷中的亚油酸、α-亚麻酸和花生四烯酸是LOX催化氧化反应的底物,产生共轭双烯过氧化氢,这种由LOX催化的脂解过程称为脂质氢过氧化。水稻基因组编码14个LOX[53],其中5个已被克隆和鉴定,包括3种LOX同工酶(LOX-1、LOX-2和LOX-3)[54-55]和HI-LOX[56]。LOX-1、LOX-2和LOX-3在水稻种子寿命中具有不同的功能,而其他的则参与生物胁迫反应。其中,LOX-1和LOX-2是影响种子寿命的关键基因[57],LOX-3是影响水稻抗虫性的关键基因[58]。在3种LOX中,LOX-3占总LOX活性的80%[59]。LOX-1和LOX-2氧化不饱和脂肪酸主要产物是13-氢过氧化物(HPOD),米糠变质过程中产生的挥发性产物的主要成分是13-HPOD的降解产物1-己醛,但是挥发性的13-HPOD及其裂解物1-己醛对害虫是有毒的;LOX-3氧化不饱和脂肪酸主要产物9-HPOD,LOX-3的存在可以将LOX-1、LOX-2产生的13-HPOD转化为不利于13-HPOD裂解酶的物质,还可能会与LOX-1和LOX-2竞争脂肪酸,从而减少13-HPOD的产生[58],因此,LOX-1、LOX-2含量少的米糠可提高耐储性,LOX-3含量少的米糠可提高抗虫害性。
4 米糠酸败机制
糙米中油脂含量相对稳定,因为在完整的谷粒内脂解酶主要位于种皮的交叉细胞中,而大部分油脂储存在糊粉层和胚芽中,经过研磨操作,将这种相互隔离稳定状态破坏[59]。稻谷中的脂质主要集中在种胚中,其次是种皮糊粉层,所以脂质主要分布在由种胚和种皮构成的米糠中。这些脂质在米糠中活性较强的脂肪酶、酯酶、LOX的作用下[60],发生水解、氧化等反应形成自由基和挥发性羰基化合物,使米糠酸价迅速升高,未酯化的脂肪酸赋予食物苦味和霉味,而氧化化合物会产生腐臭的异味[61],米糠酸败机制如图1。此外,米糠酸败速度还与微生物作用、稻谷品种以及总酚类物质、总黄酮和总花青素含量有关[62]。
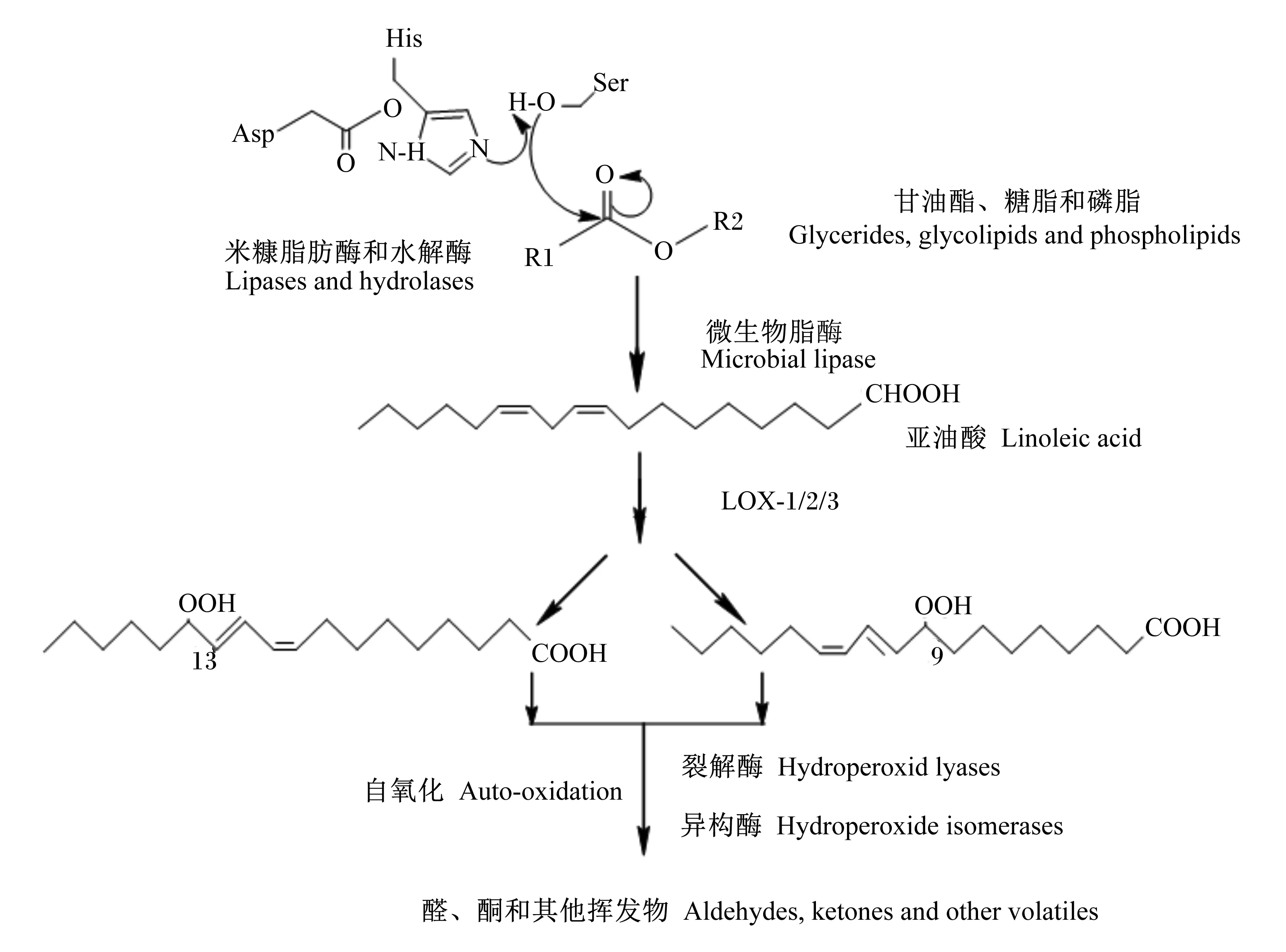
图1 米糠酸败机制
4.1 米糠水解酸败
碾米后,当脂解酶和油水界面接触,在疏水残基和亲水残基作用下,α-螺旋盖打开,酶的活性中心暴露并与底物接触,米糠中脂解酶活性最高的脂肪酶-Ⅱ,它的结合位点是1个浅碗结构,底物可从顶部进入与活性中心Ser结合,在形成的氢键与脂肪酶疏水作用的稳定下发生酰化反应[64]。在活性中心His残基参与下,Ser被激活,羟基上的质子氢转移到His残基的咪唑环上,Ser羟基残基随后增加了亲核性,攻击底物的羰基形成四面体中间复合物,在羰基的氧上带负电荷,氧阴离子洞通过形成稳定的电荷分布,降低了四面体中间复合物态能,然后发生脱酰化步骤,亲核试剂攻击酯键的碳原子,使得酯键断裂,His咪唑环再度将质子氢转移给Ser的阴离子氧,从而释放出大量的挥发性脂肪酸和甘油[65]。米糠脂解酶催化效率与酶的结合位点、Ser羟基与裂解甘油三酯酯键的距离有关[66]。
4.2 米糠氧化酸败
米糠水解酸败产生的挥发性脂肪酸、亚油酸和亚麻酸等不饱和脂肪酸,作为氧化酸败的底物进一步被LOX催化。LOX催化含有1,4异戊二烯结构的多不饱和脂肪酸特定位置并定向双加氧,生成具有共扼双键的脂氢过氧化合物再被脂氢过氧化物裂解酶和脂氢过氧化物异构酶降解为具有挥发性的己醛、戊醛和戊醇等羰基类低分子化合物,从而产生酸败味[59,63]。不饱和脂肪酸也是自氧化反应的主要反应物,自氧化是一种由光、热、氧、水分、金属和类金属等各种环境因素作用发生的非酶脂质氧化过程[67],米糠中的脂质过氧自由基不断攻击新的脂质分子发生链式反应,直到因氢源不可用而终止。不饱和程度决定了脂肪酸的氧化稳定性,3个双键的亚麻酸去除氢原子所需的活化能最小,更容易氧化,据报道,亚麻酸、亚油酸、油酸和硬脂酸的氧化速率比为2 500∶1 200∶100∶1[68]。
4.3 微生物作用
微生物作用也是米糠变质的重要原因。米糠富含蛋白质和油脂等营养物质,极易受湿、热、氧、霉等影响而变质。饲料霉变的主要因素有:饲料水分含量高(或水活度大)、热量的产生、遭雨水淋湿、保管不当等[69]。而且米糠中脂肪含量较高,稻谷中的优势霉菌如黄曲霉、黑曲霉、烟曲霉快速增殖,产生大量微生物源脂肪酶等其他酶类与米糠自身酶类共同作用分解米糠中脂质、淀粉、蛋白质,加速米糠酸败[70]。有报道称,米糠在50 ℃、相对湿度70%的环境下储存6 d引起的米糠发热霉变会造成米糠油色泽明显加深[71]。米糠被霉菌污染后,除了会引起霉变变质外,更为严重的是这些霉菌可以产生具有致癌、致畸、致突变效应的霉菌毒素,而且其中具有相当强耐受力的毒素,在饲料加工过程中无法去除。
5 解决米糠酸败的研究进展及建议
脂解酶比LOX具有更高的热稳定性,而且储存的谷物水分含量较低时,LOX活性较低[72]。因此,针对米糠脂解酶活性设计稳定方法,是延长米糠货架期的科学方法。本文比较了近5年国内外10种抑制脂解酶效果较好的稳定米糠方法(表3),主要包括了热处理(红外辐射、挤压膨化、流动微波辐射)、非热处理(紫外线、低温、有机酸、磁性固定化、木瓜蛋白酶)和基因法(酶缺陷、底物缺乏、增加天然抗氧化剂含量)。热处理中红外辐射、挤压膨化、流动微波辐射都可用于工业中的大规模加工,微波和挤压处理的便利高效性更加适合工业化生产。但是,有报道称热处理分解了一些膜结构,从而使一些极性脂质暴露于降解酶下以及使热不稳定抗氧化剂失活,从而产生更高水平的挥发性氧化产物[61]。非热处理中低温(-80 ℃)处理并没有使脂肪酶失活,只是一种暂时性小规模稳定方法;紫外线照射处理引起蛋白质变性从而使酶失去催化活性,而且与热处理方法相比能耗更低,对酸值、过氧化值以及γ-谷维素、生育酚含量影响不大,有成为适用于大规模低成本稳定方法的潜力;经磁性固定化处理的木瓜蛋白酶重复利用性得到提高,拓宽蛋白酶的pH范围及降低了对温度的敏感性;从基因角度培育脂肪酶缺陷/底物缺乏/含天然抗氧化剂的稻谷品种无疑是最经济的方法,但是还需要进一步深入研究。
虽然,以上稳定方法对脂解酶活性抑制都取得了不错效果,但当前米糠的利用率仍然极低,这是因为在发展中国家的主要大米加工厂缺乏稳定设备,存在稳定成本过高、稳定后米糠品质下降等问题。从以上的报道可以看出,木瓜蛋白酶抑制脂肪酶活性效果最好,但实际生产中常因酶的高昂价格、温度和pH的限制影响其进一步大规模推广。而生产蛋白酶的另一种生物技术——微生物发酵法,其在生产蛋白酶时不仅不受原料约束,且营养需求低、提取工艺快速简单,蛋白酶回收率高,还可以降低饲料中抗营养因子,改善饲料品质,调控畜禽肠道健康,增强畜禽机体免疫力,提高动物的生长性能,更容易实现大规模自动化生产[83]。目前,国内外对发酵米糠的研究主要集中在提取生物活性物质、富集米糠蛋白、提高米糠中酚类成分和清除自由基等方面[84-85]。而利用益生菌中丰富的蛋白酶系抑制米糠脂解酶活性的研究却尚未报道。因此,寻找可以分解米糠脂解酶的益生菌,利用发酵工程技术或许是一种能有效稳定米糠酶活性的新方法。除此之外,在发酵过程中益生菌大量繁殖,还可抑制由于微生物作用引起的米糠酸败。
6 小结与展望
米糠富含优质蛋白质、膳食纤维,具有独特的谷维素、生育酚、生育三烯醇等抗氧化剂。近年来,米糠在功能食品中的应用呈增长趋势,但米糠的稳定是其有效利用的前提。综上所述,为了进一步推广米糠的有效利用,首先必须充分了解脂质过氧化过程中涉及的关键基因和酶;其次所选择的稳定方法必须经济可行,便于实施,且有效抑制脂解酶活性;最后要尽量减少对米糠中蛋白质、淀粉、活性物质营养成分的影响,保持米糠的功能性。目前关于米糠稳定化的研究较多,但不同稳定化方法对米糠的营养成分、风味和气味的影响研究相对较少,因此有必要在动物模型上评价稳定后米糠的营养价值与安全性。在全面禁抗大背景下,发酵饲料是一种很有应用前景的抗生素替代品,是一种创造高附加值产品的同时高效转化低价值资源的可行方法,还可以生产出许多生物活性成分。但是利用益生菌中丰富的酶系实现抑制米糠脂解酶活性的同时提高其附加值的最佳发酵工艺有待进一步深入研究。
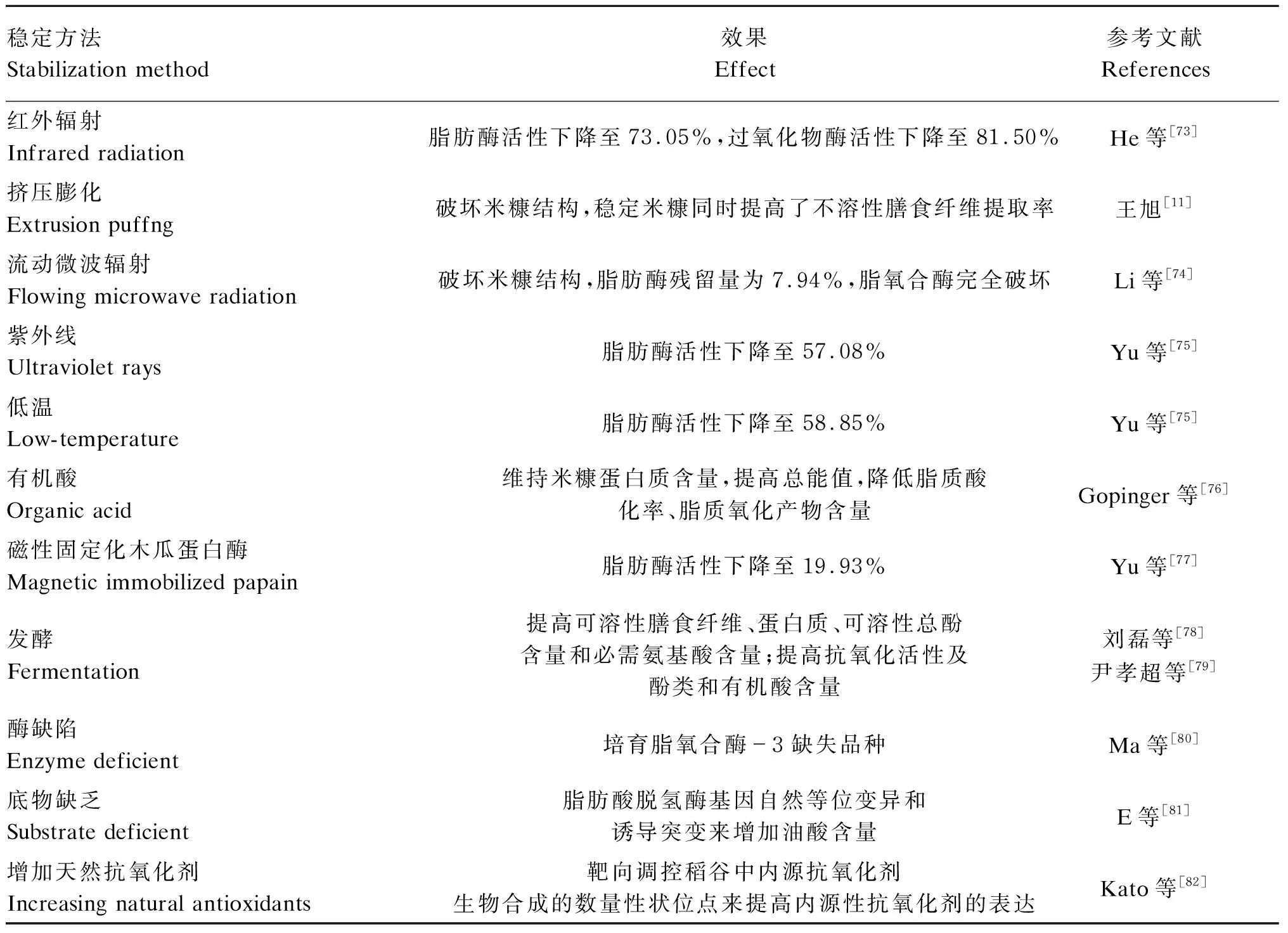
表3 近5年国内外稳定米糠的方法
