基于生物能量学模型的尖吻鲈精准投喂管理辅助决策系统构建
2021-04-01刘晓娟沙宗尧李大鹏王春芳
刘晓娟 沙宗尧 李大鹏 王春芳
(1.华中农业大学水产学院, 长江经济带大宗水生生物产业绿色发展教育部工程研究中心, 池塘健康养殖湖北省工程实验室, 武汉 430070; 2.武汉大学遥感信息工程学院, 武汉 430079)
养殖成本、养殖环境和水产品质量安全是当前水产养殖业转型升级发展面临的主要压力与问题[1]。传统投料方式不精准、不均匀和劳动力成本高等问题, 是导致养殖成本上升、养殖环境污染加剧和养殖动物疾病暴发的主要原因[2]。不同养殖品种、养殖环境与生长阶段, 其摄食量及摄食效果具有很大的差异。精准投喂是根据养殖对象的生物习性和生长规律, 通过改变养殖对象的投喂方式和投喂量, 最大限度地提高饲料的利用效率, 减少饲料浪费, 降低过量投喂而造成的水体污染, 改善产品的品质, 从而促进养殖从业者的养殖效益最大化[3]。随着互联网技术的高速发展, 5G技术的日渐成熟, 渔业规模化、集约化、工业化和机械化的不断进步, 我国水产业的发展进入了“新拐点”。探索更标准、更智能、更精准的投喂管理策略, 实现新型标准化、数据化、信息化、智能化、产业化的智慧渔业将会是水产养殖业由量变转到质变的重要途径[1]。
目前, 已有一些依托于互联网的设备和智能平台应用于农业生产过程中, 为实现农业的精准和智能化养殖做出了极大的贡献, 如养殖环境监控、产品质量溯源、生产记录分析和物流跟踪管理等平台已被广泛的地应用于实际养殖中[4], 使水产养殖装备工程化、生产规模化、技术精准化和管理智能化水平大大提高。在德国, 可在计算机控制下完成畜禽饲料精准投喂和奶牛数字化挤奶等多项工作[5]。在挪威, 使用声加速度传输器可以在计算机后台有效监测循环水养殖系统(RAS)中大西洋鲑(Salmo salarL.)的各项活动, 通过数据分析为大西洋鲑提供精准饲料投喂策略[6]。上海奉贤区及无锡地区的对虾智能管理系统和物联网智能控制管理系统的应用, 有效地提高了水产养殖的经济效益、产品质量和管理效率[7]。
随着国家对智慧渔业的不断探索和推广, 对鱼类生产数据分析应用的重要性将日趋凸显。生产管理平台利用互联网信息技术, 对生产过程信息化监测, 采集了大量的渔业生产基本数据。另外, 近年来, 国内外学者已经对水产养殖进行了大量的研究, 积累了海量的数据和模型[2,3,8—13]。利用大数据技术进行数学建模, 通过采集养殖过程中的生产数据, 将数据加以分类、挖掘和分析, 最终把有价值的信息提取出来, 将有用的结果以图表的形式直观地呈现给水产养殖生产管理者和决策者, 是解决水产养殖品种多样、养殖模式各异、养殖环境复杂和影响因子过多而导致的精准监测、检测和控制难等问题的最优途径[14]。
在水产养殖的研究中, 一些学者在水产动物的营养代谢和能量收支研究的基础上, 构建了许多数学模型。一些模型已经被应用到了养殖过程中, 为水产养殖业提供精准投喂策略, 成为养殖管理的重要工具[15—17]。然而, 它们大多都是依靠以往的经验和短期的室内试验完成, 其适用范围较窄, 此外, 这些传统的数学模型之间较为分散, 系统性不强, 且在实际应用中, 需手动计算, 计算步骤烦琐, 分析过程复杂, 大大增加了人工劳动成本。假设能够将渔场中养殖生产的数据和已有数据库及文献库中积累的大量数据收集, 进行处理分析, 在此基础上构建一套系统的、标准化的、针对养殖鱼类生长表现来进行评估的模型, 并利用互联网信息技术, 实现数据的后台输入、采集、更新、储存、校正和管理, 自动形成丰富的分析图表, 不仅有助于养殖生产人员根据模型提供的模拟数据有效地进行养殖生产管理, 还可实现养殖过程全跟踪, 使传统的水产养殖生产管理方式向精准化和数字化发展, 养殖信息和生产方式由经验主义向模型标准化转变[12],进一步降低养殖成本和环境成本, 提高养殖效率和经济效益。因此, 我们需要进一步采集养殖生产中的海量数据, 利用大数据技术和互联网平台, 构建更加实用有效、简单快捷和精准智能的投喂管理模型。
尖吻鲈(Lates calcarifer)隶属于鲈形目(Perciformes), 尖吻鲈科, 尖吻鲈属。俗称盲鰽, 亦称尼罗河鲈, 具有适应能力强、食量大、生长快、病害少、肉质鲜美和营养价值高等优点, 是我国南方地区及东南亚地区主要的养殖鱼类。本文以尖吻鲈为研究对象[17], 通过挖掘养殖场和文献中的大量生产数据, 基于生物能量学原理探讨不同养殖环境条件下尖吻鲈的能量需求, 对不同生长阶段的尖吻鲈生长规律进行研究, 构建出尖吻鲈生长、投喂和污染排放模型, 能够有效预测某一时期尖吻鲈的生长情况, 对饲料的需求量及氮、磷等污染物的排放量,有助于鱼类的差异化上市、减少饲料浪费、降低养殖成本、提高水体透明度和养殖效益。将此模型植入计算机网络端, 构建精准投喂管理辅助决策系统, 通过该系统将互联网等信息技术与水产养殖投喂管理紧密结合, 以水产养殖水体监测、鱼类生长、鱼体营养成分、饲料营养成分及饲料投喂数据采集和处理分析为基础, 以数据统计和综合评价为核心, 从投放鱼苗到成鱼上市进行信息化管理,实现大数据与传统水产养殖技术的融合创新; 为水产养殖生产管理者和决策者提供方便有效的投喂管理工具, 为数字化渔业提供一定的技术支持。
1 材料与方法
1.1 生长阶段的划分
尖吻鲈生长数据主要来自中山市强晟水产养殖有限公司。养殖时间从2018年6月至2019年5月,共计11个月, 在养殖过程中分别记录水温、鱼体体重和饲料投喂量等数据(表1), 本研究选用了4种最常用的养殖动物生长预测模型, 其计算公式如下:
特定增长率(SGR, %/d)=100×(lnWt-lnW0)/t
日增长系数(DGC, g1/3/d)=100×()/t
日均增重(ADG, g/d)=(Wt-W0)/t
热积温系数[TGC, g(1-b)/(d×℃)]=100×
式中,W0为初始体重,Wt为终末体重,T为养殖水温,t为养殖天数。根据上述公式, 尖吻鲈的生长预测公式如下:
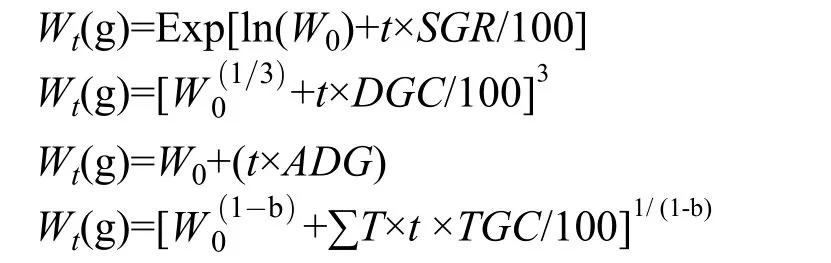
在TGC的计算中, 指数(1-b)先设定为1/3, 随后采用模型生长预测值与实际养殖观测值的最小残差平方和RSS法校正[18], 使RSS达到最小值,RSS采用迭代法计算, 工具采用Microsoft Excel 2016[19]。RSS公式计算如下:

式中,Yj为模型生长预测值,yj为实际生长观测值。
1.2 生化成分分析
用于生化分析的尖吻鲈为养殖场采样所得, 体重从21—1128 g共计10个不同体重组, 并采用AOAC[20]标准测定法对尖吻鲈的生化成分进行测定。采用尖吻鲈体重和其水分、蛋白、脂肪、灰分和能值含量之间的回归分析对尖吻鲈的各生化成分进行预测。
1.3 各能量的估算
尖吻鲈各能量计算数据主要来自已发表的文献[17, 21—30], 基础代谢能(HeE)采用呼吸实验中耗氧量来计算(氧热系数为13.6 J/mg)。鱼类基础代谢能和其体重之间关系可用HeE=A×WB描述, 其中,鱼类的指数B值为0.8, 故基础代谢能的计算公式如下[3]:

式中, a、b为常数,T为水温,W为体重。
储积能(RE)是通过计算初末鱼体能值之差来估算, 排泄能(UE+ZE)为鱼体鳃和尿液排出的能量,通过公式24.9×(UN+ZN)来计算[3],UN和ZN分别为鱼体鳃和尿液排出的氮的含量。摄食热增能(HiE)主要为鱼类摄食过程中消耗的能量, 计算公式如下:

式中,IE为摄食能,FE为粪能,GEt和GE0分别为终末和初始鱼体能值,UN和ZN分别为鱼体鳃和尿液排出的氮的含量。不同生长时期鱼类消化能需求量可通过计算储积能(RE)、基础代谢能(HeE)、摄食热增能(HiE)及排泄能(UE+ZE)之和来估算[3], 故尖吻鲈消化能需求量计算公式如下:

1.4 饲料需求量估算
在确定饲料中各营养成分含量及其消化率的基础上, 可计算出饲料消化能(DE)含量, 计算公式如下[30]:
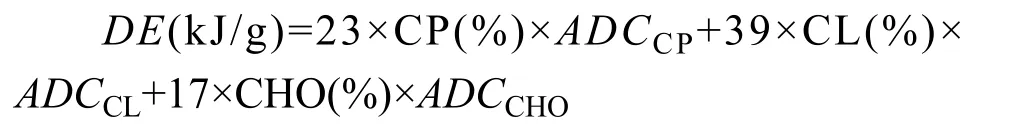
式中, CP、CL和CHO分别为饲料蛋白质、脂肪和碳水化合物,ADC为表观消化率。饲料需求量计算公式如下:
饲料需求量(Feedreq)=消化能需求量(DEreq)/饲料消化能含量(DEof feed)
1.5 废物排放量的估计
水产养殖过程中所排放的污染物(TW)主要分为固态污染物(SW)和溶解态污染物(DW), 其预测公式如下:

式中, DM为干物质,ADC为表观消化率, 以上式子同样可运用于估算N、P废物的排放量, 具体计算公式如下:
固态氮污染物=摄入的氮(Nintake)×(1-ADCN)
固态磷污染物=摄入的磷(Pintake)×(1-ADCP)
溶解态氮污染物=消化的氮(Ndigested)×ADCN-储积的氮(RN)
溶解态磷污染物=消化的磷(Ndigested)×ADCP-储积的磷(RP)
1.6 验证试验
尖吻鲈生长和饲料需求模型的模型验证数据主要来自中山市强晟水产养殖有限公司。对养殖场中用于养殖不同生长阶段尖吻鲈的饲料营养成分进行分析(表2), 尖吻鲈生长模型用模型预测体重与实际观测体重比较来验证。饲料需求模型通过比较饲料转化率(FCR)的实际观测值和模型预测值来验证。

表1 各池塘水温(℃)、尖吻鲈初末体重(g)及饲料投喂量Tab.1 Water temperature (℃), interval time (days), initial body weight (g), final body weight (g), and feed consumption (g/kg fish) of Lates calcarifer in different ponds (g/fish)
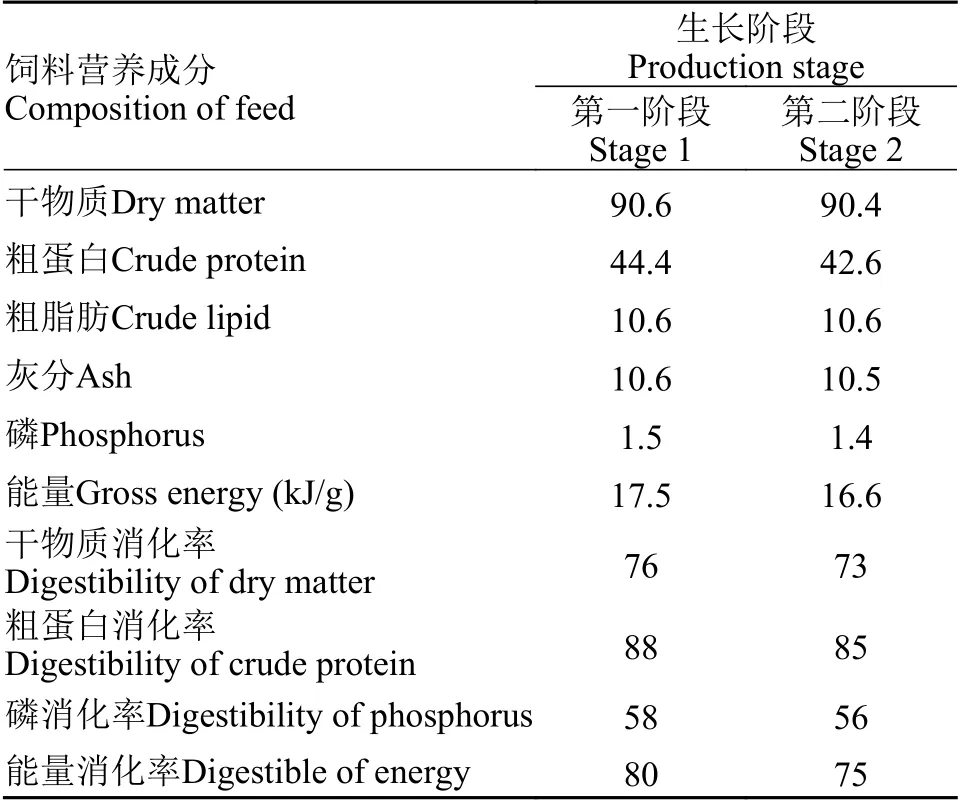
表2 尖吻鲈不同生长阶段饲料营养成分及其消化率Tab.2 Feed composition and digestible energy in different growth stages of Lates calcarifer (%)
1.7 软件系统开发
软件系统包括三层架构, 分别为界面层(UI)、应用层和数据访问层(RL, 图1)。界面层是基于HTML5/CSS集成开发, 支持任何浏览器和移动设备的访问, 可为软件使用人员提供交互式操作的可视化界面。应用层是基于Java Server-Pages (JPSs)开发, 负责执行算法数据分析, 及处理软件系统的业务逻辑, 提供各种后端服务(如鱼体质量计算、饲料需求量计算和污染物排放量计算等)来响应来自UI层的命令。数据访问层用于存储与养殖品种相关的所有模型的参数, 以及运行软件包所需的其他必要数据。为了使该工具更好地应用于实际养殖中, 本研究中采用的apache tomcat(v8.0)是基于Java技术实现的, 可同时用作应用服务器和Web服务器, 它的优势主要在于它的源代码开放, 支持跨平台的应用, 使开发的软件包可以运行在几乎所有的(包括Windows和Linux系统)平台上。
2 结果
2.1 生长模型的选择

图1 基于web系统的鱼类精准投喂管理模型辅助决策系统的软件设计框架Fig.1 Conceptual framework of web-based precision feeding models
尖吻鲈生长数据来自中山市强晟水产养殖有限公司, 共计7个池塘104个数据(表1), 并根据上述公式计算尖吻鲈不同生长时期各生长模型(SGR、DGC、ADG和TGC)的值。结果显示, 不同生长时期尖吻鲈TGC的值呈现一定的规律, 当尖吻鲈体重为5—1000 g时,TGC的值中出现1个异速生长点, 根据该点的位置(两条线段的交点), 尖吻鲈的生长周期可分为2个生长时期(图2): 5—296 g(第一生长期)和>296 g(第二生长期)。
尖吻鲈在不同生长时期的各个子模型的最适体重指数(1-b)存在显著差异。无论是在第一生长期还是第二生长期, 指数调整后的TGC模型的残差平方和都显著小于SGR、DGC、TGC和ADG等生长模型(P<0.05, 表3)。因而, 当尖吻鲈体重在1—1000 g时, 相较于其他生长模型, 优化后的TGC模型对尖吻鲈生长的预测效果更好(图3a、3b)。
2.2 鱼体生化成分
随着尖吻鲈体重的变化, 其体生化成分如水分、蛋白质、脂肪和灰分相对含量变化趋于平稳(图4)。因此, 简单的线性方程可以有效估计尖吻鲈蛋白质(图5a)、脂肪(图5b)、灰分(图5c)和能量(图5d)含量。本研究分别选用了线性方程y=aX+b和指数方程y=aXb对不同体重尖吻鲈的体生化成分进行预测, 结果发现, 线性模型预测尖吻鲈体内蛋白质和能量含量的效果比指数方程好, 而在脂肪和灰分的预测上, 则指数方程优于线性方程(表4)。当对尖吻鲈鱼体磷含量分析时, 同样发现, 线性模型对鱼体磷含量的预测效果好于指数方程(表5)
2.3 各能量的估算
尖吻鲈生化成分组成、基础代谢能、摄食热增能、排泄能及消化能需求量的计算公式见表5。当水温在15—35℃时, 尖吻鲈的HeE随水温的增加先增加, 当达到最适温度后, 则逐渐开始下降。另外, 研究还发现, 尖吻鲈摄食热增能与储积能和基础代谢能之间呈正相关关系(图6a); 排泄能则与摄食能和消化能密切相关, 为7.37%的储积能、基础代谢能和摄食热增能之和(图6b)。
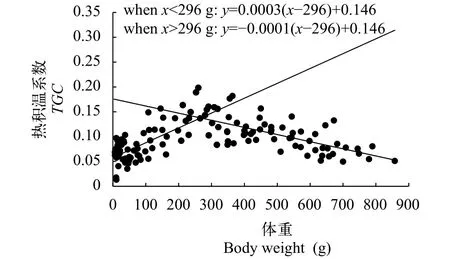
图2 不同生长阶段尖吻鲈积温系数Fig.2 Thermal-unit growth coefficient (TGC) as a function of body weight of Lates calcarifer

表3 尖吻鲈在不同生长时期SGR、DGC、ADG和TGC及校正后的TGC模型平均值、体重指数和残差平方和(RSS)Tab.3 The regression coefficients, body weight (BW) exponents and RSS of SGR, DGC, ADG, TGC and revised TGC models for different stages of Lates calcarifer
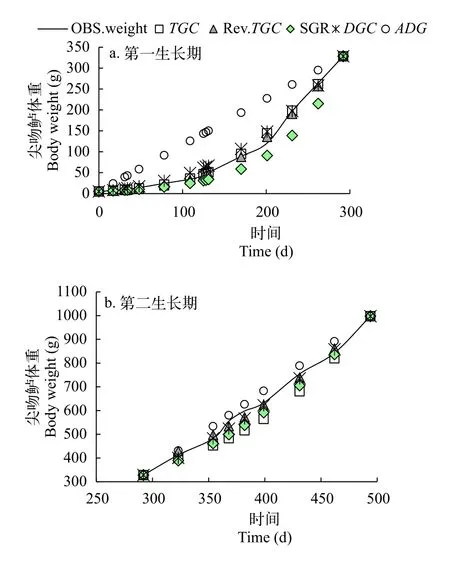
图3 不同生长模型的比较-分别用SGR、DGC、ADG、TGC和Rev.TGC生长模型来描述尖吻鲈的生长Fig.3 Comparison of growth models- DGC, SGR, ADG, TGC and Rev.TGC models to describe the growth of Lates calcarifer
2.4 饲料需求量、污染物排放量和模型验证
尖吻鲈消化能需求量、饲料需求量见表6, 各污染物排放量预测结果见表7。经估算, 当尖吻鲈体重从1 g增长到1000 g时, 每条鱼消化能需求量为2.33×104kJ, 饲料需求量约为1.57 kg, 每生产1000 kg鱼, 总固态污染物排放量为543 kg, 可溶性含氮污染物为56.6 kg, 可溶性含磷污染物为6.1 kg。当尖吻鲈体重在1—1000 g内, 每消耗1000 kg饲料, 总固态污染物排放量为349 kg, 可溶性含氮污染物为36.3 kg,可溶性含磷污染物为3.9 kg。
在模型的验证中, 本研究分别采用尖吻鲈体重和饲料转化率(FCR)的模型预测值与实际观测值之间的拟合度来验证, 结果发现尖吻鲈体重实际观测值与模型预测值之间的拟合度较好(R2=0.97, 图7),而FCR的实际观测值与模型预测值之间的拟合度相对较差(R2为0.57, 图8b), 且饲料需求模型对FCR的预测值明显小于实际观测值(图8a、8b)。
2.5 软件包的开发

图4 尖吻鲈鱼体生化组成与体重的关系Fig.4 Whole body chemical composition of Lates calcarifer and its relation with body weight

图5 尖吻鲈体重和鱼体蛋白质(a和e)、脂肪(b和f)、灰分(c和g)和能值(d和h)之间的等距和对数回归分析Fig.5 Isometric and allometric relationship between whole body mass and body protein (a and e), lipid (b and f), ash (c and g) and gross energy (d and h) of Lates calcarifer
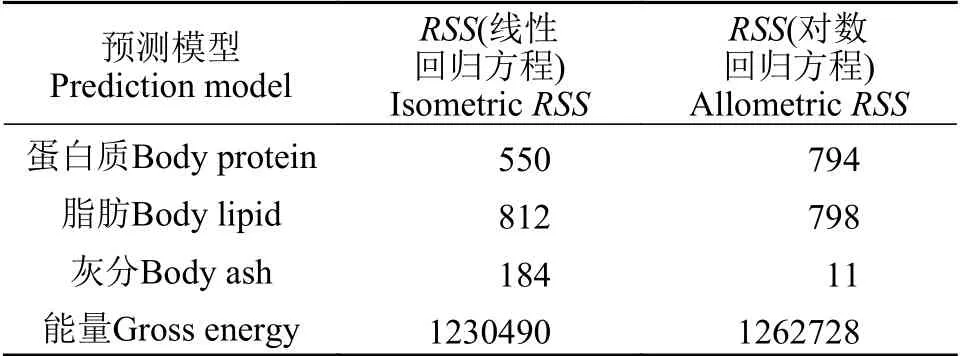
表4 线性方程和对数方程(体重与BP、BL、BA、GE)预测值与实际观测值的残差平方和(RSS)Tab.4 Residual sum squares (RSS) of the isometric and allometric regression analysis of body weight with body protein, lipid, ash and gross energy as predictors of Lates calcarifer
为实现该模型的可视化, 本研究开发了模型在线软件包。在数据库方面, 设计了10个表来存储软件包的模型参数和辅助信息(图9)。鱼类管理表是用来存储不同品种鱼类的相关信息, 包括整个系统中鱼类的唯一编码、鱼类的名称及用于计算积温的最低温度。鱼类生长阶段表具有唯一的鱼类生长阶段编码, 该编码定义为表中特定鱼类的生长阶段。个人信息表包含系统中所有注册的用户。对于任何用户, 在用户模型表中建立的一些模型均可以与该用户关联。饲料管理表用于存储不同饲料产品的相关参数, 这些参数(如干物质、粗蛋白、粗脂肪、能值、磷及其消化率等)是预测鱼类饲料需求量的污染排放量所必需的。每个模型中都存储了与模型相关的参数, 用户可在需要时检索这些参数(例如, 鱼类品种、鱼类生长阶段、饲料参数和其他模型参数等)。此外, 用户还可根据其自身生产数据自定义某些参数, 例如, 用户可以通过输入其养殖对象以往周年的生长数据, 使后台自动生成更加匹配其生长环境的生长模型参数, 在预测生长的时候选择该新生成的参数。
应用层可实现模型所有功能的可视化。例如上述已建立模型的鱼类体重预测、饲料需求预测及污染排放预测功能。此外, 该层还可提供诸如用户注册和用户登录之类的其他辅助功能(图10)。图10展示了用户界面, 已注册的用户登录到系统后, 显示主菜单, 从中可以访问开发的各项预测功能。用户界面允许用户与系统交互,TGC可以来自理论值或采用用户自定义参数。UI层是基于html5/css标准开发的, 任何浏览器和移动设备均可进行访问, 可为用户切换访问设备时提供更好的用户体验。
3 讨论
3.1 生长模型
一些研究发现, 控制肌肉纤维聚集和肥大的基因的表达可能会影响鱼类的生长速度, 导致鱼类在其生长周期中存在生长异速点[6,8,19]。根据该点的位置可将鱼类整个生命周期分为几个不同的生长阶段。本研究通过计算不同体重鱼类的TGC值, 同样发现尖吻鲈在其生长周期中存在生长异速点, 但与虹鳟(Oncorhynchus mykiss)[19]和罗非鱼(Oreo-chromisspp.)[3]的生长周期中含有两个异速生长点不一致的是, 本研究中尖吻鲈仅存在一个异速生长点。一些研究表明鱼类在其性成熟时期对营养物质的利用效率会发生改变, 从而导致生长速度发生改变[19]。尖吻鲈是一种雄性先成熟的雌雄同体的鱼类, 雄性性腺先发育成熟, 而后转变为雌性性腺发育成熟, 亚洲尖吻鲈幼鱼达到雄性成熟需要3—4年, 体重一般为2 kg以上, 而达到雌性成熟则需要6年, 体重一般为4 kg以上[31]。而本研究主要针对商品规格的鱼类(>400 g即可上市), 研究对象体重小于1 kg, 故还需要收集更多较大规格(>2 kg)的尖吻鲈生长数据, 才能确定尖吻鲈在其生长周期是否还存在更多的异速生长点。

表5 尖吻鲈生化成分组成、基础代谢、增值热、排泄代谢能及消化能需求量方程Tab.5 The equations for the estimation of body chemical composition, basal metabolism, heat requirement of feeding, urinary-branchial excretion and digestible energy requirement

图6 尖吻鲈热增能(HiE)和排泄能(UE+ZE)与其他各能量之关系Fig.6 The relationship between heat increment of feeding (HiE), urine and gills energy (UE+ZE) and other energy of Lates calcarifer

表6 尖吻鲈不同生长时期消化能需求量和饲料需求量Tab.6 Digestible energy and feed requirement of Lates calcariferat different stages
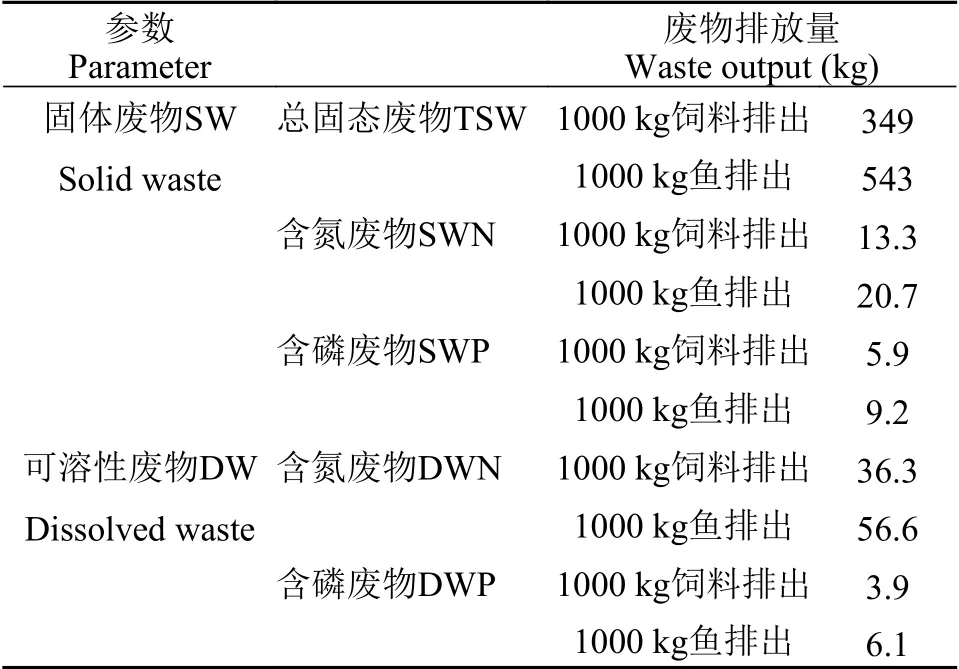
表7 尖吻鲈固态废物和可溶性废物排放量Tab.7 Output of solid and dissolved wastes of Lates calcarifer
目前, 国内外最常用来描述鱼类生长的模型主要为SGR、DGC、ADG和TGC模型[3]。其中,SGR模型由于在恒温条件下对幼鱼的生长预测效果较好, 因而经常被一些研究者用于鱼类生长实验中,来探索更适合鱼类生长的饲料配方和养殖模式[3]。但由于鱼类为变温动物, 鱼类的生长速度会随着水温的变化而发生改变,TGC模型由于考虑到了水温对鱼类生长的影响, 因此在变温条件下, 其预测效果优于其他模型[15]。一些研究表明, 不同生长时期的鱼类其生长速率存在差异, 鱼类在其生长周期中存在生长异速点[3,8,19], 因此, 采用单一系数的模型来预测不同生长时期的鱼类的生长速度会存在较大的偏差。Chowdhury等[3]的研究结果表明当罗非鱼体重小于30 g时, 生长指数为1/3的TGC模型才具有较好的预测效果; Dumas等[19]对虹鳟的研究同样发现, 当生长指数为1/3时,TGC模型只适用于体重在20—500 g内的虹鳟, 且TGC模型的最适生长指数(1-b)随着鱼体质量的不断上升逐渐增加, 最终的理论数值会趋近1。因此, 在构建鱼类生长模型时, 需要考虑到这一因素并对生长模型的参数进行优化以提高生长模型预测的准确性。本文研究发现, 根据生长异速点将尖吻鲈的生长周期划分为两个生长阶段后, 优化后的TGC模型的预测效果明显优于其他的模型, 这与罗非鱼[6]、虹鳟[19]、异育银鲫(Carassius auratus gibelio)[8]和草鱼(Ctenopharyngodon idellus)[10]的研究结果相似, 但由于鱼类品种不同, 其生长规律和生长潜力具有较大的差异, 在一定程度上直接影响生长模型的最适参数值, 因而本研究构建的TGC模型的最适生长指数与其他鱼类均存在较大的差异。
3.2 尖吻鲈鱼体成分和能量收支
鱼类体重的增长是饲料中各营养成分在鱼体中沉积的结果, 因此, 在探讨鱼类的生长和对饲料的需求时, 需要了解鱼类对饲料中各种营养物质的需求范围和利用效率。一些研究表明, 运用数学模型可以较好反映鱼类体重与鱼体中各营养物质的关系[16,32—34]。本研究分别对尖吻鲈体重与其体内各营养成分之间的关系进行了分析。结果显示, 在不同的鱼类体重、养殖水温、饲料组成和投喂水平的情况下, 线性等速方程y=a×W+b和指数方程y=a×Wb均可以用来预测不同体重鱼类各体成分的含量。通过对这2种模型的预测结果进行比较分析,研究发现线性等速模型在预测鱼体蛋白质和能值的效果比指数模型更好, 这与罗非鱼[6]、虹鳟[19]和异育银鲫[9]的研究结果一致。但在对尖吻鲈体内脂肪和灰分的预测中, 指数模型的预测效果比等速模型效果更好, 这一研究结果仅在对虹鳟脂肪的预测中发现, 而其他鱼类体内脂肪和灰分均为等速增加,这可能与鱼类品种差异和饲料营养成分不同有关,但具体原因还要进一步研究。

图7 尖吻鲈体重的实际观测值和模型预测值及其回归关系Fig.7 Observed body weight and predicted body weight of Lates calcarifer

图8 尖吻鲈饲料系数(FCR)的实际观测值和模型预测值及其回归关系Fig.8 Observed and predicted feed conversion ratio of Lates calcarifer
基础代谢能(HeE)是动物用来维持生命所有器官活动所需要的最低能量。一些研究发现, 同种鱼类在不同体重和不同养殖水温情况下, 其单位体重消耗的基础代谢能具有很大差异[12], 通过对鱼类能量学的进一步研究发现, 采用指数方程HeE=(a+bT)×BW0.8可以反映水温和鱼体质量对鱼类HeE之间的关系[3,12,8]。在本研究中, 鱼类基础代谢随水温呈先上升后下降的趋势, 这与虹鳟[12]的结果相一致, 原因可能与鱼体内有关基础代谢的酶的活性相关, 具体原因还需进一步研究。鱼类代谢活动中所损耗的能量除基础代谢能外, 在摄食活动中还会有一部分能量消耗, 这部分能量被称为摄食热增能(HiE),一些研究发现,HiE与HeE和RE之间具有高度相关关系。在本研究中, 分别将尖吻鲈的HiE与HeE和RE进行回归分析, 结果发现HiE和RE+HeE的相关度最高, 这与罗非鱼[3]和异育银鲫[8]的研究结果一致。UE+ZE为鱼类鳃和尿液排泄过程中所损失的能量。有研究表明,UE+ZE与摄食能(IE)具有一定的关系[3], 本文对尖吻鲈的研究发现,UE+ZE与RE+HeE+HiE之间呈现线性回归关系, 此结果与罗非鱼[3]和草鱼[10]的研究结果一致, 但回归系数差异较大, 原因可能与养殖品种、养殖环境及饲料营养成分相关。

图9 数据库端设计中的关键表Fig.9 Key tables in the design of the database side
3.3 废物排放和模型验证
一些研究表明, 水产养殖的污染排放和营养负荷同样可采用数学模型有效预测[35]。本研究分别对不同生长阶段鱼类摄食的饲料各营养成分、饲料各营养成分消化率及不同质量鱼体中氮和磷含量进行了分析, 通过营养物质平衡法估算尖吻鲈养殖过程中的污染排放量。结果显示, 当生产1000 kg尖吻鲈或者投喂1000 kg饲料时, 总固态污染物排放量为543和349 kg, 其结果明显比草鱼[10]的排放量(分别为632和440 kg/1000 kg)低, 但显著高于虹鳟[35](分别为250和220 kg/1000 kg)和异育银鲫[8]的排放量(分别为419和282 kg/1000 kg)。原因可能与不同鱼类和不同的饲料对饲料营养成分的消化吸收效果具有差异相关。
饲料系数(FCR)是评价饲料报酬的一个重要指标[30], 在动物养殖中, 通常被用来反映鱼体对饲料的利用效率。有研究显示, 由于鱼类生长与饲料需求具有直接的关系, 而FCR作为衡量鱼类对饲料转化为体重的效果的主要指标, 同样可应用于鱼类饲料需求模型的验证中[3,12]。本文通过对模型鱼体体重及FCR的模型预测值和实际观测值进行比较, 结果发现, 本模型对尖吻鲈体重的预测值和观测值的拟合度较好(R2=0.97), 而FCR的预测值和挂测值的拟合度相对较差(R2=0.57), 且FCR观测值显著大于预测值, 表明该模型对尖吻鲈生长的预测效果较好,而对饲料需求量的预测中与实际投喂量有一定偏差, 然而, 采用同样的方法, Chowdhury等[3]对罗非鱼FCR的预测效果较佳, 原因可能在于罗非鱼的研究在实验室进行, 投喂量较为精确, 而本研究投喂数据来自养殖场, 在实际投喂过程中投喂过量所致。
3.4 软件系统的开发
在水产养殖业中, 饲料投喂量对水产养殖业的生产成本、废物排放和水产品质量具有决定性的影响[3]。因此, 精确投喂被认为是提高饲料利用率、降低饲养成本和污染排放的重要途径。然而,由于水产养殖品种众多, 养殖环境多变, 养殖模式复杂, 所谓的鱼类精准投喂仍处于探索期, 虽有一些尝试使用自动投饵机、传感器监测设备和NBIoT无线通信技术来设计水产养殖智能管理系统[36],但因为覆盖低、成本较高, 且易受环境变化干扰等原因而无法大量投入使用。此外, 这些系统只针对各种传感器监测数据做日常监测, 并未将所有数据如饲料投喂数据、水质监测数据及鱼体生长数据关联起来综合分析, 各数据之间职能单一, 相互封闭, 在实际养殖过程中无法对生产进行有效的管理。以互联网、大数据为核心的智慧渔业是一个全新的产业模式, 虽没有现成的经验和技术作为参考, 但水产养殖的精准化、数字化、智能化的重要需求及5G技术的快速发展态势为未来水产养殖智能信息管理平台提供了良好的发展环境。
图10 所开发软件包的前端用户登录界面和预测结果界面(UI)Fig.10 Front-end user interface (UI) of the developed software package
在大数据的基础上构建模型, 能够推进大数据技术与水产养殖产业的深度融合与应用。当数据样本数大幅增大的同时, 庞大的数据可以显著提升模型的精确度, 且在一定程度上降低对数据的要求[37]。基于互联网技术的软件平台是实现水产养殖智能管理最有效的技术手段之一。本研究分析大量水产养殖生产数据的基础上构建模型, 并为养殖户及管理者建立了一个线上投喂管理辅助决策系统, 以使养殖户能够更加方便快捷的使用本研究所提出的模型预测服务。该在线系统具有以下功能: (1)海量数据记录与储存, 用户注册或登录系统后, 可将所养殖鱼类名称、水温、体重和饲料各营养成分输入, 系统将会自动储存这些数据, 以方便对养殖场以往生产数据和现有数据进行对比分析;(2)生长预测: 用户可根据上传养殖场的生产数据,如水温、初始体重和养殖时间, 使用当前研究计算出的默认TGC生长率预测鱼的体重。也可以通过上传自己以往的年生长和水温变化等生产数据, 计算自己的TGC生长速率, 做进一步的生长预测;(3)饲料需求量预测: 用户可以将养殖场不同生长阶段所用的饲料各营养成分输入系统, 系统结合鱼类生长预测结果可自动估计饲料需求量; (4)氮、磷污染物排出量预测: 系统结合鱼类生长预测和饲料需求量预测结果, 可自动估计该鱼塘在养殖期间总氮、总磷的排放量。所有结果将自动以表格或图表形式显示, 任何用户都可以对养殖情况一目了然。所有养殖户或渔业管理人员都可以通过任何浏览器和移动设备访问这个网上水产养殖智能精准养殖预测服务平台来进行鱼类生长、饲料需求量和污染排放量的预测。
在软件的开发和使用过程中, 本研究实现了电脑(PC)端网页的开发, 但因为随着智能手机的普及,及部分渔民对电脑的使用存在障碍, 后期的研究将会开发手机移动端, 并将此养殖预测服务平台与水质监测设备、自动预警报警系统及一些自动化设备(如自动增氧机、自动投喂机)进行耦合, 使渔民不仅能够随时随地的在手机上直观的观察鱼塘的水质变化情况, 预测鱼类的生长、饲料需求和氮磷排放, 还可以通过自动预警报警系统对鱼塘的异常情况进行及时反应, 并通过自动投喂机显示的实际投喂量对模型预测投喂量不断的优化, 进一步提高模型对应用鱼塘的预测准确度, 从而实现针对不同渔民养殖鱼塘的精准养殖预测功能。
4 结论
本研究构建的精准投喂管理辅助决策系统耦合了体重和各营养载体、能量之间的数据模型, 及尖吻鲈不同生长时期TGC生长模型的应用, 验证实验中尖吻鲈体重的模型预测值与实际观测值之间显著相关, 饲料系数(FCR)模型观测值大于实际预测值。这表明本实验中构建的精准投喂管理模型可以有效地估计养殖中鱼体生长情况, 但由于实际养殖中饲料投喂多为过量投喂, 可能导致实际投喂量大于养殖需求量, 进而导致污染排放量的增加。本研究中开发的精准投喂管理辅助决策系统软件,为养殖投喂管理精准化、信息化奠定了一定的理论和技术基础。
由于模型预测准确性取决于高质量的大量数据, 当有效数据量越多, 模型的准确性越高。本模型验证的数据主要来自某一海鲈、尖吻鲈渔场, 因此在预测其他渔场某种鱼类的生长和饲料需求时,还需要采集大量该养殖场的生产数据, 并将数据输入此平台中, 通过对数据的处理和分析, 平台将自动调整模型参数, 以使该模型预测结果更加精确。
