植物受高温胁迫机理及对玉米生长影响的研究进展
2021-04-01卫晓轶洪德峰马俊峰王稼苜
卫晓轶,洪德峰,马俊峰,马 毅,王稼苜,魏 锋
(河南省新乡市农业科学院,河南 新乡 453002)
随着全球工业化的不断加快,煤炭、石油以及各种燃料使用产生的气体和温室效应的加剧,全球气候变暖的程度越来越明显[1-4]。全球气候的改变主要表现在平均气温的升高和大气层CO2浓度的增加,这些对植物的生长和产量产生了很大影响[5]。有推测表明,到21世纪中期,极端年日最高气温将会增长1~3 ℃[6]。随着气候的改变,高温已成为影响主要粮食作物生长发育的重要因素[4,7]。在植物生长季节,极端高温灾害下平均气温每增加1 ℃,其产量估计将损失6%~7%[8]。植物已经进化出了应对复杂多变的环境温度气候条件以及对生长发育至关重要的相互联系的信号途径[9-10]。了解植物对高温胁迫反应的分子机制对提高作物对高温的耐受性具有重要意义。
1 热害胁迫的机理研究
1.1 热害胁迫对植物的影响
研究认为,开花期高温胁迫会导致花粉成熟及花药开裂、花粉管的伸长受阻、花粉萌发受到影响,造成不育,导致结实率降低[4,11]。Prasad等[12]认为,高温胁迫下代谢过程会受到严重影响,花粉活力及萌发能力明显下降。水稻在抽穗扬花期对高温极为敏感,高温胁迫极易导致花粉败育和子房受精受阻,空粒率提高,结实率下降,从而造成严重减产[13-17]。小麦在灌浆初期,在高温胁迫条件下,胚乳细胞数量减少,从而使粒重减少。在灌浆后期,高温胁迫条件下,通过限制籽粒同化物供应或直接影响籽粒中淀粉的合成,从而使淀粉的合成受到损害,进而影响产量[18]。
1.2 植物响应热害胁迫的分子机理
高温容易引起蛋白质变性,导致内质网(ER)中错误折叠的蛋白质积累,从而引发植物未折叠蛋白反应(UPR)。Liu等[19]研究发现,膜结合转录因子OsNTL3是水稻苗期耐高温胁迫所必需,OsNTL3的功能缺失突变体对高温胁迫敏感,而无跨膜结构域的截短形式的OsNTL3的诱导表达提高了水稻幼苗的耐热性。Li 等[20]报道,褪黑素(MT)可能会增强玉米苗期的耐热性,主要通过调节抗氧化防御,MG解毒和渗透调节系统。Shi等[21]和Xu等[22]研究认为,外源褪黑素可能引起热激因子和热激蛋白的表达,从而增强拟南芥和番茄苗期的耐热性。由L-DES产生的内源H2S至少部分参与了热害胁迫下,烟草悬浮细胞培养物中Ca2+和CaM的诱导表达[23]。小热休克蛋白(sHSPs)是迄今为止最复杂的一组热休克蛋白,由于其异常的丰富性和多样性,它们的功能是在热害胁迫下,保护植物中几乎所有的细胞区隔[24]。Yang等[25]研究认为,高等植物在热害胁迫下,叶绿体LeHSP 100/ClpB参与表达。乙烯介导的信号转导参与了氧化损伤的减少和叶绿素含量的维持,并且使热胁迫下水稻幼苗耐热性增强。此外,研究发现,热休克因子和乙烯信号相关基因参与了水稻幼苗耐热性的复杂网络调控[26]。谢晓金等[27]、陈仁天等[28]研究表明,在高温胁迫条件下,水稻花药中的脯氨酸(Pro)、游离氨基酸、可溶性蛋白质和还原型谷胱甘肽(GSH)含量均显著降低,且随气温升高和高温胁迫时间延长,其花粉活力和萌发率、结实率和产量均有不同程度下降。张桂莲等[29]研究认为,高温胁迫下,水稻花药保持较高的抗氧化酶活性、较好的花粉散落特性和花粉萌发特性及较低的膜透性和丙二醛(MDA)含量,是品种耐高温的生理基础。
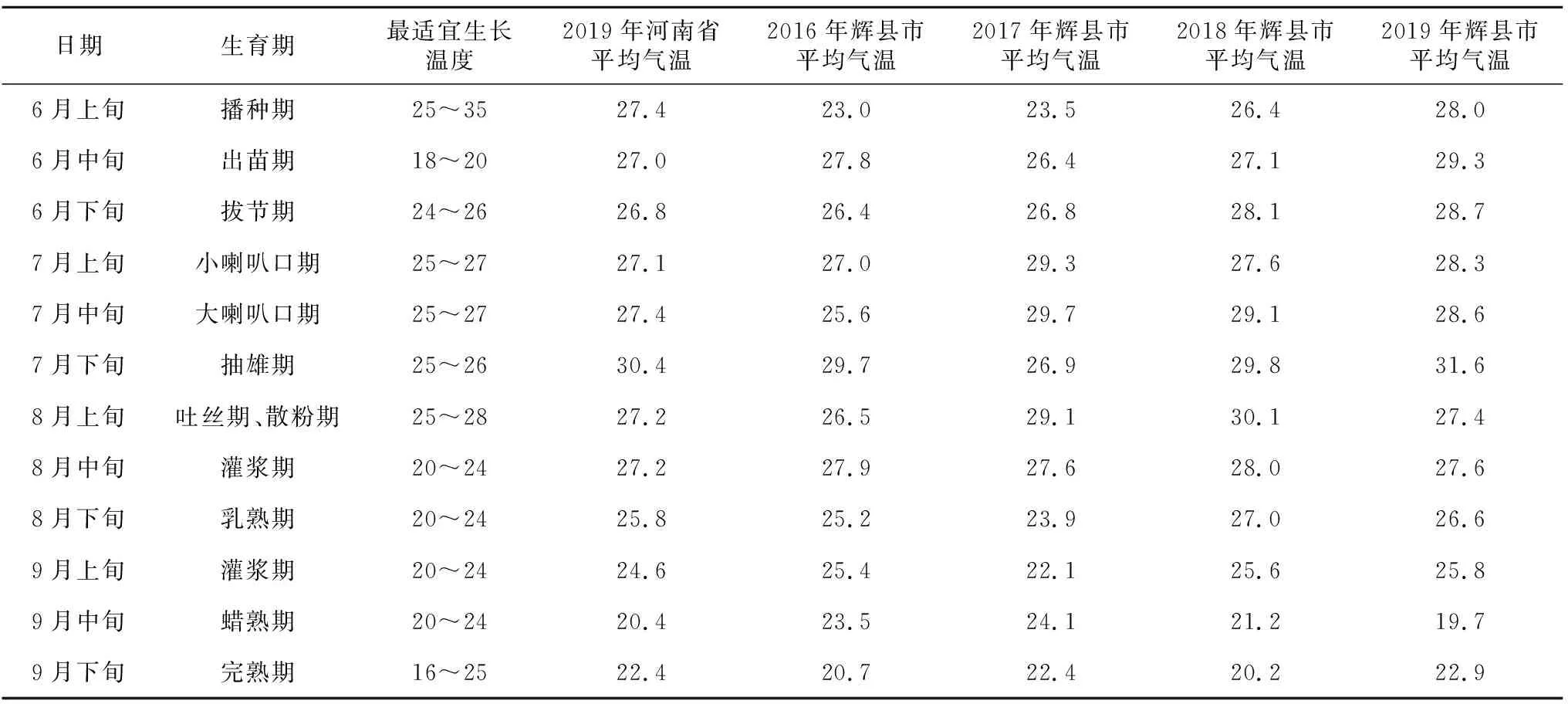
表1 玉米生长各生育期最适宜生长温度 单位:℃
1.3 热害胁迫对植物的损伤类型
热害(高于最佳生长温度5℃以上)通常导致植物至少四种以上类型的损伤。
1) 活性氧(ROS)引起的氧化应激,从而导致蛋白质变性,膜脂质过氧化反应、核酸(DNA和RNA)损害。
2) 甲基乙二醛(MG)引发的二羰基应激,甲基乙二醛(MG)能迅速与生物大分子(蛋白、脂肪、核酸)发生反应。
3) 由于水分缺失引起的干旱造成细胞内失水,导致渗透胁迫。
4) 生物膜完整性缺失[30-32]。因此,热害是一种主要的非生物胁迫因素,影响细胞的新陈代谢,植株的生长发育全过程。植物本身已经进化出相应的有效机制来应对初级和次级胁迫,这些机制涵盖抗氧化防御机制、MG解毒系统、渗透调节系统和应激蛋白等。抗氧化防御机制包括抗氧化酶(愈创木酚过氧化物酶,GPX;谷胱甘肽还原酶,GR;过氧化氢酶,CAT;超氧化物歧化酶,SOD;抗坏血酸过氧化物酶,ATX)和抗氧化物(抗坏血酸,AsA;谷胱甘肽,GSH)[30-35]。MG解毒系统包括乙二醛酶Ⅰ,GlyⅠ;乙二醛酶Ⅱ,GlyⅡ[36-38]。渗透调节系统包括脯氨酸,Pro;甜菜碱,GB;海藻糖,Tre;可溶性总糖,TSS[34,35,39-41]。应激蛋白包括热激蛋白,HSPs;胚胎发育晚期丰富蛋白质,LEA[31,33-35]。
1.4 玉米对热害胁迫响应机理的研究
玉米不仅是重要的粮食作物,也用于饲料、燃料、工业等。近年来,玉米的基因组测序已经完成,许多重要基因的生理学功能已得到注释,玉米已成为一个新的模式作物。在玉米苗期,植株对高温胁迫十分敏感,从而也影响着玉米的产量和品质[42]。Li等[43]报道,丙酮醛可能会增强玉米苗期的耐热性,部分通过H2S调节。高温条件下,在玉米的营养生长阶段,K+和Ca2+的吸收显著增加,N、S、Mg2+和Na+的吸收显著降低[44]。在玉米苗期,高温胁迫增加了植株中可溶性糖和蔗糖的含量[45]。在玉米的开花期,高温胁迫会直接影响玉米花粉活力和花粉管的生长[46]。Ottaviano等[47]报道,细胞膜的稳定性作为一个生理学指标,已广泛应用于评价植物的耐热性。过氧化氢(H2O2)和一氧化氮(NO)在植物细胞中起信号分子的作用。近年来,硫化氢(H2S)也被发现具有类似的功能,但它们在植物受热害条件下,相互作用机理仍不明确。Li等[48]研究表明,H2O2处理可以提高玉米幼苗的耐热性,H2O2可能参与了NO与H2S之间的下游信号互作,从而提高了耐热性。Li 等[49]认为,在热激信号转导途径中,植物热激转录因子起重要的调节作用。在玉米受热害胁迫下,ZmHsf12可能参与了信号响应表达。
2 高温热害对玉米生长的影响
表1为河南省玉米各生育时期最适宜生长温度[50],及玉米生育期内2019年河南省平均气温,并以豫北一个玉米试验点河南省辉县市为例,列举了2016—2019年以来,玉米生育期内辉县市平均气温,与河南夏玉米最适宜生长温度进行比较。可以看出,从出苗期至灌浆期,2019年河南省、2016—2019年辉县市平均气温多数高于玉米最适宜生长温度,尤其从7月中下旬至8月中下旬,2019年河南省及2016—2019年辉县市平均气温高于玉米最适宜生长温度0.4~5.6 ℃。
图1为2016年—2019年辉县市6月—9月平均气温。从图1可以看出,从2016—2019年,河南省辉县市每年7月中下旬到8月下旬的平均气温均高于玉米最适宜生长上限温度。此时河南省夏玉米正经历穗分化期、开花期和灌浆期,因此应重点关注高温胁迫对这个阶段的玉米造成的影响。
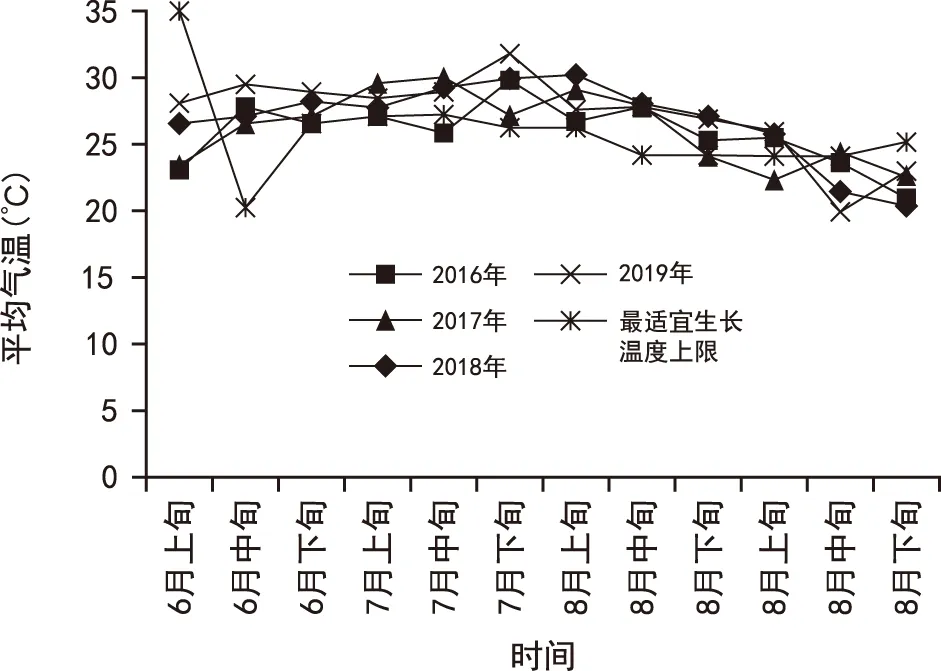
图1 2016年—2019年辉县市6月-9月平均气温
玉米在不同阶段受高温胁迫会使玉米减产程度不同[51],玉米在开花期对高温较为敏感,在高温胁迫下,淀粉合成受到抑制,导致授粉率降低[4]。而且,玉米开花后籽粒的生长发育受高温胁迫的影响也较大[52],高温胁迫会导致籽粒发育不正常,造成减产[53]。赵龙飞等[17]研究表明,开花后受高温胁迫对玉米叶片光合速率和抗氧化能力及产量等产生的影响大于开花前受高温胁迫。在玉米穗分化时期、散粉吐丝期,高温胁迫会影响花粉花丝的形态结构,比如穗分化时期高温胁迫会造成雄穗分枝减少、一个穗位节有多个果穗共生的现象[54]。在散粉吐丝期高温胁迫会有两种表现形式:一是花粉发育不正常或者败育,导致花粉数量减少、生活力减弱,受精能力减弱或丧失;二是花粉粒分裂异常,形成畸形花粉粒。导致花粉数量和活力降低,从而影响受精结实[55]。研究表明,高温影响玉米雌穗发育,造成雌穗吐丝困难,延缓雌穗吐丝或吐丝不协调,从而使授粉受到影响。高温还容易导致雌穗吐丝后花丝失水,花丝表面无粘液,生活力丧失,影响花丝寿命,使花粉在柱头上萌发受阻,影响玉米授粉过程,且气温越高,花丝寿命越短[4,56-59]。Yang等[60]研究认为,当温度高于38 ℃时,会降低玉米花粉的萌发能力和花粉管的伸长能力,导致结实率平均降低21%。花后前期高温胁迫下,不同灌浆时期玉米籽粒的淀粉合成相关酶活性降低,从而使淀粉含量降低,且籽粒中的 3-吲哚乙酸(IAA)和玉米素核苷(ZR)含量均显著下降[59,61-62]。Engelen-Eigles等[63]研究表明,授粉后6 d,35 ℃高温处理玉米胚乳细胞有丝分裂周期4 d,会抑制有丝分裂细胞进入核内复制周期。
3 采取措施
热害胁迫使植物的生长、光合作用、矿质营养和产量性状均受到一定的影响。热胁迫引起植物不同部位的超微结构改变和氧化应激。植物可以通过保持膜的稳定性、调节抗氧化剂和可溶性物质以及清除活性氧来承受一定程度的热害胁迫。植物耐热性可通过选择耐热性品种、基因工程、以及渗透液、微生物、矿质营养、土壤改良剂的外源施用和适当的农业措施来提高[64]。
