海洋植物谱系地理模式与遗传连通性研究进展*
2021-03-30胡自民杜玉群梁延硕钟凯乐
胡自民 杜玉群 梁延硕 钟凯乐 张 杰①
海洋植物谱系地理模式与遗传连通性研究进展*
胡自民1, 2杜玉群1, 3梁延硕1, 3钟凯乐1, 3张 杰1, 2①
(1. 中国科学院海洋研究所 实验海洋生物学重点实验室 中国科学院海洋大科学研究中心 青岛 266071; 2. 青岛海洋科学与技术试点国家实验室 海洋生物学与生物技术功能实验室 青岛 266237; 3. 中国科学院大学 北京 100049)
海洋植物谱系地理学虽然在国内受到的关注比较有限, 但近年来的研究进展对了解环境变动如何影响物种的多样性结构、生态适应和进化潜力等亦提供了重要信息。本文首先对刊发海洋植物谱系地理研究最具代表性的两本国际期刊《》和《》进行搜索和筛选, 从研究团队的地域分布和研究对象的阶元等角度纵览全球范围内近20年间海洋植物谱系地理学研究的现状和特征。其后, 本文从洋流如何驱动海洋植物谱系地理结构和多样性、入侵物种的种群遗传变异特征和路径重建、冰期避难所如何影响现今海洋植物遗传结构和地理变动、克隆繁殖和谱系多样化等四个方面概述了这一领域的主要成果进展。最后, 本文还从岛屿地理模型、共生物种的谱系协同进化和高通量数据应用等三个方面对海洋植物谱系地理学的发展趋势进行了展望。
海藻; 海草; 气候变化; 冰期避难所; 入侵遗传学; 遗传多样性保护
1987年, 加州大学尔湾分校的John C. Avise教授在《Annual Review of Ecology and Systematics》上撰文首次提出‘Phylogeography’的概念(Avise, 1987), 标志着谱系地理学这门学科的正式诞生。在随后的20—30年间, 该学科旨在利用蛋白质和DNA序列数据研究物种内遗传谱系的亲缘关系、地理分布格局特征和形成机制(Avise, 2000)。时至今日, 随着分子数据获取的日益便捷(如高通量测序)和相关领域研究理论与技术的快速发展(如统计遗传学和溯祖理论), 谱系地理学被广泛用来研究生态适应、新种形成、物种互作和气候变化等全球热点问题(Pereyra, 2009; Jormalainen, 2017; Wernberg, 2018)。
谱系地理学从诞生伊始就定位为连接宏进化(如新种形成、物种互作、群落和生态系统遗传学)和微进化(如瓶颈效应、环境选择、基因渐渗和漂变)之间的桥梁(图1)(Avise, 1987), 涉及种群遗传学、生态学、古地质学、分子进化和生物地理学等多学科的交叉融合。不仅如此, 谱系地理学还对许多相关学科的发展产生了重要影响, 特别是进化遗传学、生物地理学、保护生物学和分类学(图1)(Beheregaray, 2008)。以谱系地理学为中心还衍生了一些新的学科与研究方向, 如统计谱系地理学和基于溯祖理论的多学科交叉等(图1)。特别是比较谱系地理学, 它通过对某一地区拥有不同生态位的分类单元的谱系多样性特征、遗传结构、时空变动历史和相关驱动因素进行整合分析(Meta-analysis), 不仅可以挖掘该地区物种进化历史的共性规律和过程, 而且为物种多样性保护和生态系统功能维护提供了重要参考(Hoareau, 2013)。
《》和《》是两本聚焦于生态、进化、多样性和生物地理过程等学术前沿的国际著名期刊, 分别创刊于1992年和1974年, 迄今已发表大量主题为谱系地理学的文章。其中, 《》自1993年以来发表的仅题目中包含“phylogeography”一词的论文就有440篇。海洋植物(本文仅指海藻seaweed和海草seagrass)是植物界或海洋生物中很小的一个分支, 全球范围内从事海洋植物谱系地理学研究的团队更是屈指可数。尽管如此, 海洋植物谱系地理结构和遗传变异研究在过去20年间依然取得了重要进展, 许多成果就发表在《》和《》上, 极大地推进了我们对海洋生物多样性变动、环境驱动的适应进化和物种形成等问题的认知。本文旨在对近20年间发表在《》和《》上海洋植物谱系地理学的代表性成果进行汇总, 帮助国内外同行了解这一领域的研究现状和挑战, 同时对一些值得探究的重要科学问题进行展望, 以期推动该领域在未来的长足发展。
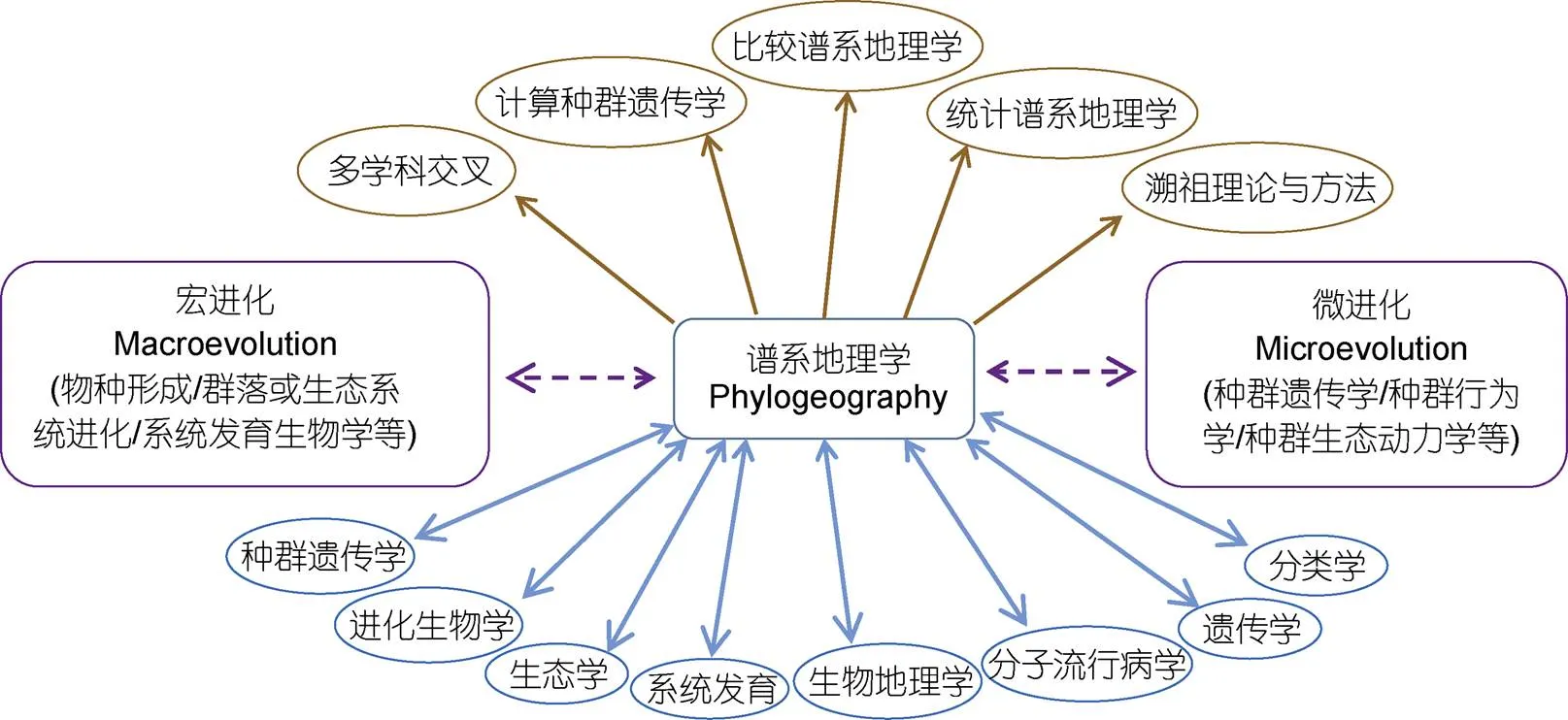
图1 谱系地理学在连接宏-微进化及其在不同学科领域的互作示意图
1 海洋植物谱系地理学研究(2000—2019) ——概览
海洋植物谱系地理学研究的历史可追溯至20世纪90年代, 其中最早的海藻种群遗传结构和多样性研究由荷兰格罗宁根大学生态与进化研究中心的Jeanine Olsen团队于1994年发表于美国藻类学会会刊()(Hu, 2016a)。为了解近20年间海洋植物谱系地理学研究的大致现状, 我们在Web of Science核心数据库(SCI)中进行搜索, 年限设为“2000—2019”。剔除与淡水种类相关的研究后, 当标题为“alga”+主题为“phylogeography”时检索到34篇论文(图2a); 当标题为“seaweed”+主题为“phylogeography”时检索到24篇论文(图2b); 当标题为“seagrass”+主题为“phylogeography”时检索到12篇论文(图2c)。刊登海洋植物谱系地理学研究的期刊虽有20多种, 但大多数期刊发表的海洋植物谱系地理学论文数量有限, 其中《》、《》和《》的发文量排名前三(图2)。同《》和《》一样, 《》也属于专业性极强的藻类学期刊, 它们发表的谱系地理研究的主题较为单一, 主要限于种群遗传变异、谱系分化和多样性分布。相反, 《》和《》上刊登的海洋植物谱系地理学论文不仅涵盖这一基本主题, 而且包括气候变化介导的谱系地理变动、适应性遗传变异和宿主(海藻)-寄主(如细菌)协同进化等诸多前沿课题。《》和《》这两本期刊或许不能全面反映近20年间海洋植物谱系地理学研究的完整现状, 但它们涵盖了当前该领域的主要研究进展、学科热点和发展趋势, 以其作为分析样板, 亦可起到“管中窥豹”的效果。
为准确获悉《》和《》在过去20年间发表的海洋植物谱系地理学研究的论文数量, 我们在其官网的高级搜索界面‘abstract’中先后输入关键词‘alga’或‘seaweed’以及‘seagrass’分别进行搜索, 时限设为2000年1月—2019年12月。经过筛选和核对, 我们发现除了2000年和2006年, 其余年份都有海洋植物谱系地理学研究的论文发表(图3)。总体上, 每年发表的论文数量在2—4篇之间, 其中2003年只有1篇, 而2016年数量最多达到9篇(图3)。在过去20年间共发表74篇论文, 其中, 《》贡献57篇(海藻42篇+海草15篇), 《》贡献17篇(海藻14篇+海草3篇)。以海藻为研究主体的论文占比76%, 以海草为研究主体的论文占比24%(图3)。在海草谱系地理学研究中, 葡萄牙学者的贡献最大, 共有4篇; 澳大利亚和意大利学者的贡献居次, 各有3篇(附录)。

图2 Web of Science核心数据库搜索结果(期刊名称+论文篇数)(2000—2019)
注: a. 标题“alga”+主题“phylogeography”; b. 标题“seaweed”+主题“phylogeography”; c. 标题“seagrass”+主题“phylogeography”
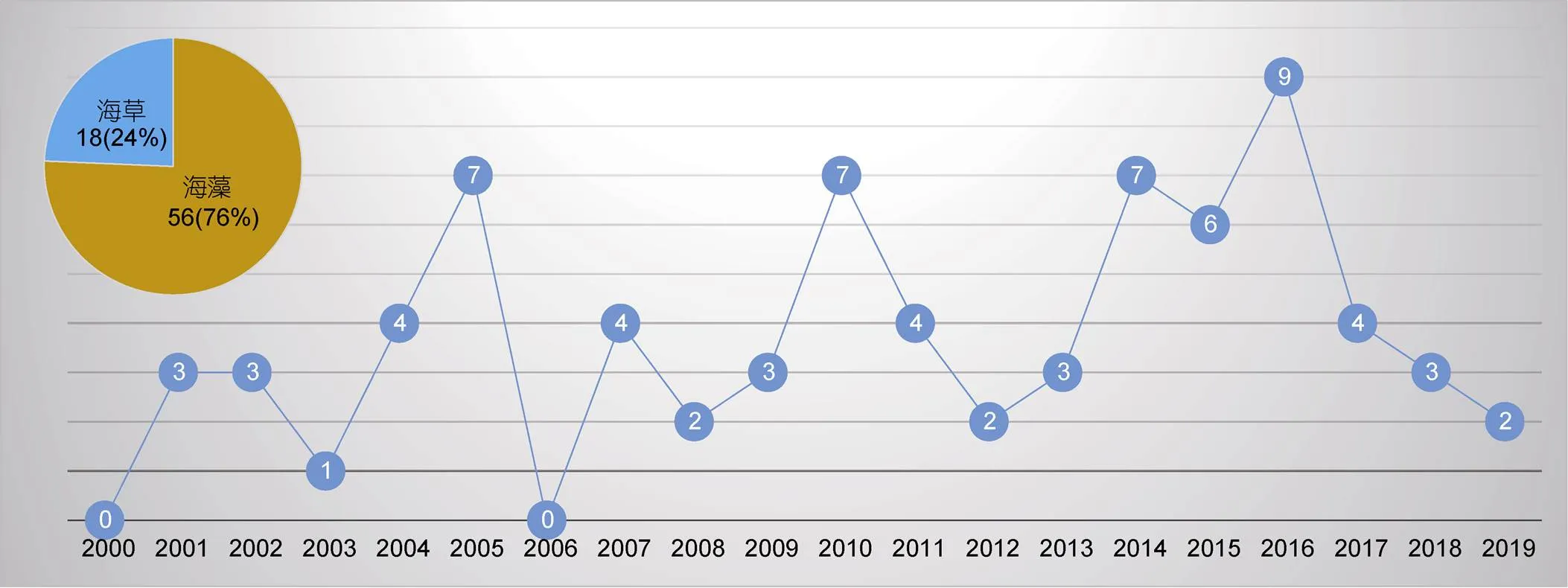
图3 2000—2019年间《Molecular Ecology》和《Journal of Biogeography》发表的海洋植物(海藻和海草)谱系地理学研究论文数量
在56篇海藻谱系地理学研究论文中, 有47篇聚焦大藻(macroalgae), 9篇聚焦小藻(microalgae)(图4)。若依研究主题对47篇大藻研究进行划分, 8篇为入侵海藻, 30篇为谱系地理结构和多样性, 6篇涉及海藻倍性和繁殖模式(图4)。若依第一作者所在机构进行划分, 葡萄牙阿尔加夫大学海洋科学研究中心(CCMAR)的Ester Serrão团队贡献最大, 共计11篇。中国学者贡献8篇论文, 其中, 中国科学院海洋研究所实验海洋生物学重点实验室胡自民研究员所属团队贡献了7篇, 格罗宁根大学生态与进化研究中心的Jeanine Olsen团队累计贡献6篇, 法国(罗斯科夫海洋生物实验站的Valero Myriam)、美国和澳大利亚的学者分别贡献4篇(图4)。若依研究材料所属门类对47篇论文进行划分, 褐藻占比最高, 共计31篇, 其次为红藻(11篇)和绿藻(5篇)(图4)。在31篇褐藻谱系地理学研究论文中, 墨角藻属共计有10篇, 马尾藻属共计有4篇, 巨藻共计有3篇, 其他褐藻种类累计14篇(图4)。若依研究对象的分布地域来划分, 超过70%(33篇)的材料分布在北大西洋, 仅有23%(11篇)的材料在太平洋(包括澳大利亚和新西兰)(图4)。9篇小藻研究中澳大利亚学者贡献3篇, 美国和法国学者各自贡献2篇。其中, 虫黄藻是小藻中的重点研究对象, 共有4篇论文(附录)。这些数据表明, 海洋植物谱系地理学研究具有鲜明的材料和地域偏倚性, 这可能与不同地区研究团队的数量、分布和兴趣爱好有关。即使是褐藻, 研究对象也仅限于少量种属。因此, 培养与海洋植物谱系地理学研究相关的人才梯队, 结合不同地区的地质气候变动历史发掘特征研究材料和学科方向, 应是后期研究工作的重点。
2 海洋植物谱系地理学研究进展(2000—2019)
2.1 洋流与谱系结构及分化
对具有长距离扩散能力的海洋生物而言, 洋流是其个体迁徙和基因交换的媒介, 可在广阔空间内产生具有高度均质性的遗传谱系(Mitarai, 2009)。与此同时, 洋流也能构成外在或无形的物理屏障, 导致在狭小空间内相邻种群的遗传异质性(Palumbi, 1994)。海洋植物(特别是海藻)在其生活史中会频繁释放单倍体的孢子和二倍体的合子, 这些微观的繁殖体(propagule)存活时间虽短, 但受潮汐和海浪作用可在微小地理尺度内逐步扩散与迁移。某些海藻(如石莼科和马尾藻科)即使漂浮在海上亦可进行生长与繁殖, 洋流的驱动使其进行长距离扩散成为可能。
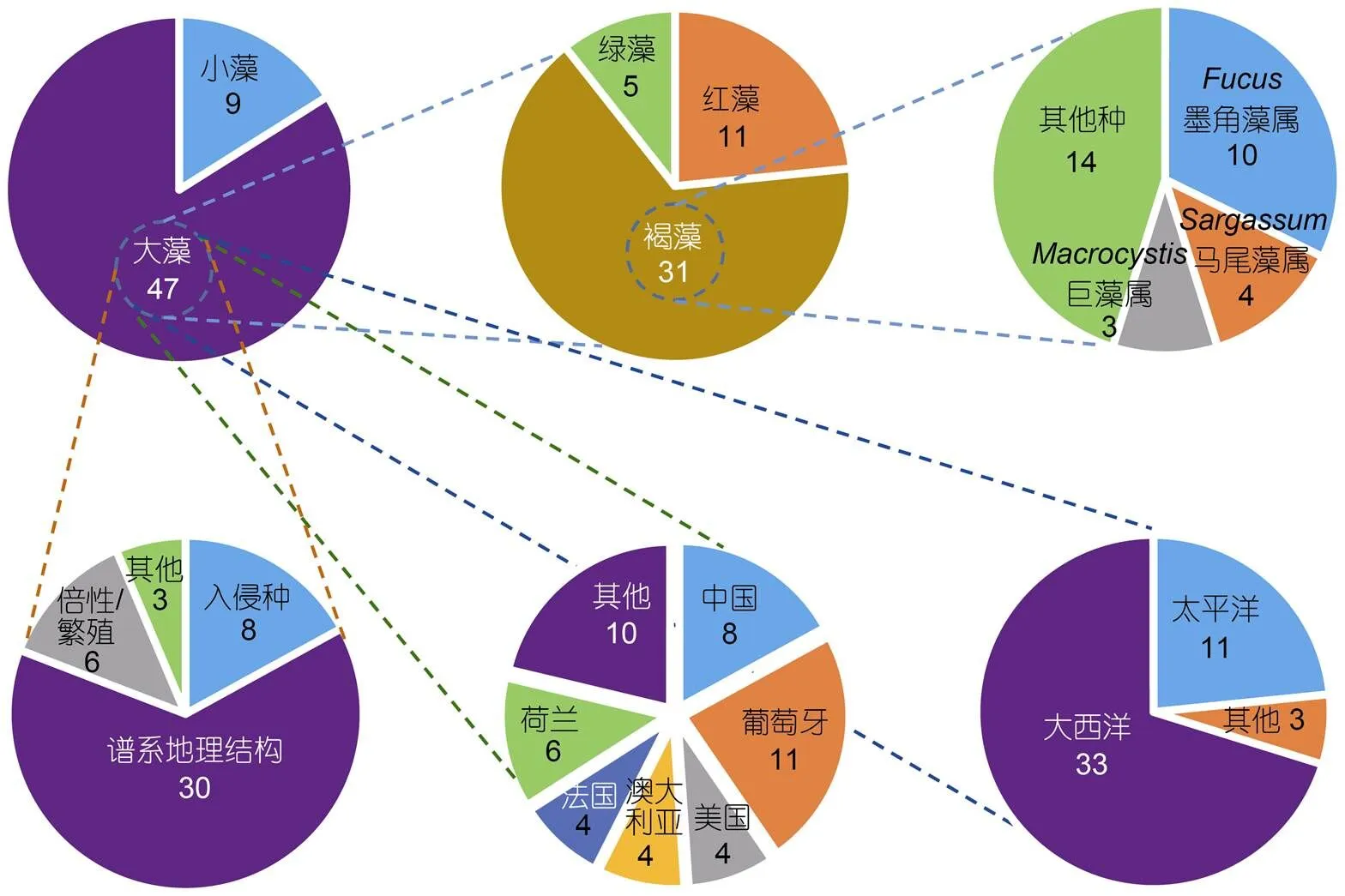
图4 基于研究对象、地区和主题等对2000—2019年间在《Molecular Ecology》和《Journal of Biogeography》上发表的海洋植物(海藻)谱系地理学研究详细划分
有研究发现, 因生活史、物候、形态和栖息地差异, 生长在北美东部太平洋沿岸(多年生)和加利福利亚湾(一年生)的大叶藻()已产生高度遗传分化(Capello-Pasini, 2003, 2004)。太平洋沿岸的厄尔尼诺-南方涛动(ENSO)使得局部大叶藻种群间发生对称基因交流, 种群遗传结构均质化, 符合典型的基石扩散模型(Stepping-stone dispersal model) (Muñiz-Salazar, 2005)。同样, 在北美大西洋的缅因州, 墨角藻()的繁殖体(如配子/合子)受持续近岸洋流的驱动进行扩散, 导致种群遗传分化模式与传统的距离隔离模型(Isolation by distance, IBD)不相匹配(Muhlin, 2008)。在西北太平洋, 黑潮及近岸洋流的流向与鼠尾藻()种群间基因流的方向基本一致, 但并不存在IBD模式(Li, 2017)。受黑潮、韩国沿岸流和利曼寒流的驱动, 济州岛成为鼠尾藻迁移的中转站, 进而汇聚了较高的遗传多样性; 在日本-太平洋沿岸, 从南向北的黑潮和从北向南的津轻暖流使得种群间双向的基因流都较大; 在中国沿岸, 从南向北的南海暖流和从北向南的中国沿岸流促进了南北种群的交流, 导致鼠尾藻种群结构呈现均质性(Li, 2017)。
事实上, 偏离IBD模式在海洋生物谱系遗传研究中很常见, 主要原因是物种的强扩散能力导致种群间的低遗传分化(如随机交配种群), 或种群的漂变-迁移平衡被破坏(如侵殖事件), 或种群间遗传分化值因欧氏距离(Euclidean distance)以外的因素所驱动(Manel, 2003)。但是, 以欧氏距离为基础的IBD模型并不能准确反应种群的遗传结构。White等(2010)将种群间的派生海洋学距离(Derived oceanographic distance, DOD)整合到IBD模型中, 发现IBD模型不能解释小尺度的空间遗传结构, 而DOD则检测到南加州圣芭芭拉海峡不同种群间50%以上的遗传变异。因此, 与基于欧氏距离的IBD模型相比, DOD在揭示种群遗传结构模式方面前景广阔。基于此, Alberto等(2011)通过高分辨率的海洋学距离模拟发现, 在南加州的圣芭芭拉, 当6月份洋流变强时, 巨藻()产生和释放孢子的效率也提升, 其揭示的遗传变异是IBD或栖息地连续模型的2倍, 凸显海洋学距离在塑造巨藻种群遗传结构(6月份最为显著)中的作用。另外, 洋流驱动的孢子平均运输时间是预测巨藻种群分化的最佳因子。因此, 将海洋环流动力学、遥感监测、数学模拟和地形地貌特征等整合到海洋植物谱系地理学研究中, 有望深入揭示海洋植物的多样性演化模式、地理扩散机制和时空变动过程, 从而精确解读海洋植物的种群进化动力学机理。
2.2 海洋植物入侵与谱系连通性
外来种入侵在世界各地时有发生, 但在入侵种对当地产生明显的生态和环境影响之前, 它们很少受到关注, 原因之一是入侵种与本地的同属种(姊妹种)的形态特征较为相似, 非专业人士很难察觉。运用谱系地理学的研究方法与技术可有效的解决这一科学难题, 包括其中潜藏的多次隐秘入侵、扩散路径反演和谱系杂交后代的可育性等。在北大西洋, 新管藻(先前在分类上被称作多管藻)广泛分布于北美大西洋西岸, 在19世纪中叶前这种红藻在英国尚未见报道, 当前分布在大西洋东岸的种群被视为入侵种。目前, 关于新管藻入侵到英国有三个假设: (1) 北美本地种群通过跨大西洋入侵到英国和爱尔兰; (2) 日本或西北太平洋的祖先种群首先入侵到北美, 通过级联引入扩散到英国; (3) 太平洋的祖先种群先入侵到大西洋的东、西两岸, 在东岸从法国入侵到英国, 随后向北和向南分别扩散至挪威和西班牙。McIvor等(2001)利用叶绿体L谱系地理数据和杂交实验证实日本是该物种分布和谱系多样性的起源中心, 欧洲的入侵种群内至少存在两个不同的遗传世系(即两次入侵), 欧洲和加拿大新斯科舍的基因型可能来自日本的北海道, 而新西兰、美国加利福尼亚州和北卡罗来纳州的基因型可能来自日本的本州, 这两次独立入侵的谱系之间完全杂交可育。因此, 谱系地理结构总体上支持假设2和假设3。无独有偶, Provan等(2005)发现, 虽然刺松藻(ssp.)在起源地日本显示出极低的遗传变异, 但在入侵地基因型分布具有明显的地理结构, 其中一个基因型仅分布于地中海, 另一个基因型则分布在西北大西洋、北欧和南太平洋, 显示刺松藻从太平洋出发经历了两次独立的入侵事件。类似的研究策略也应用于红藻海门冬入侵路线反演和隐存多样性挖掘(Andreakis, 2007, 2009)。
外来种入侵的后果之一是与本地种杂交, 发生基因渗入则造成不同基因库的混合, 对本地种造成“基因污染”。小种群入侵后若与本地种发生杂交, 也会对后者产生显著影响; 一旦生境发生变化或受到扰动, 则会导致本地种适合度的丢失, 面临较高的灭绝风险。日本新管藻()是一种入侵到美国西北大西洋的北卡罗来纳州、西班牙、澳大利亚和新西兰等地的红藻。Savoie等(2015)利用核糖体内转录间隔区(Internal transcribed spacer, ITS)检测到其中一个遗传谱系含有日本新管藻与当地新管藻的重组ITS序列, 存在核DNA从到的非定向渗入, 这可能是在其杂交后减数分裂期基因交换或重组所致。入侵种和本地种的杂交与渗入在陆地植物和褐藻中已有报道, 但这在红藻中则是首次报道, 这对于解读红藻种间杂交和新种形成的机制具有重要启示。
在众多解释引入种何以成为入侵种的物种互作机制方面, “敌害释放假说”(Enemy release hypothesis, ERH)最为知名, 它假定当捕食者、寄生虫或病原体不能与引入种共同适应(Co-adaptation)时, 前者就不能对引入种造成危害, 引入种就可分配更多资源在引入地进行生长和繁殖, 进而在进化时间尺度上形成“竞争力的提升进化”(Evolution of Improved Competitive Ability, EICA; Blossey, 1995)。杉叶蕨藻()是一种在欧洲广泛肆虐的入侵绿藻, 它的原产地为加勒比海、北印度洋、南中国海、日本和澳大利亚等热带水域。1980年, 水族人员利用澳大利亚的杉叶蕨藻为德国斯图加特Wilhelma动物园选育出一个冷水品种。1984年, 该品种被摩纳哥海洋博物馆意外释放到地中海, 即所谓“水族逃逸假说”。与此同时, 不同地区杉叶蕨藻遗传谱系的连通性显示其扩散可能还存在“海流运输假说”, 即当前的地中海谱系与水族馆有关的谱系之间遗传关系紧密。杉叶蕨藻上附生着与其永久关联的微生物区系, 它们同宿主一样保留着独特的谱系地理特征, 解析与杉叶蕨藻相关联的细菌谱系有望为追溯宿主地理起源提供独特的信息来源。Meusnier等(2001)研究了不同地区杉叶蕨藻nrDNA ITS和附生细菌的16S rDNA序列, 两组数据均显示澳大利亚的杉叶蕨藻谱系与地中海-水族馆进化支呈姊妹关系, 即地中海西部引入的杉叶蕨藻起源于澳大利亚东部。这种海藻-细菌协同进化分析为探索海洋植物入侵提供了广泛可用的鉴定范式, 若能在时空层面将其量化(Spatial and temporal quantification),则可将其发展成研究多样性地理演化和物种适应进化的重要工具。随后, Meusnier等(2002)利用PCR-SSCP技术进一步证实地中海和其他地区的入侵杉叶蕨藻谱系起源于澳大利亚的观点, 而近海岛屿和近海大陆杉叶蕨藻种群之间的生态型差异则与其所处的生境/环境条件(尤其是海水温度和浊度)有关。
海洋植物可进行二倍体的有性繁殖和单倍体的无性与营养繁殖, 繁殖方式的选择能影响物种的交配系统和长距离扩散, 进而通过改变漂变-迁移平衡来影响物种的遗传结构。Baker定律认为在远距离定殖事件后, 物种的无性繁殖能力越强, 其在入侵地成功建立种群的可能性就越大。在过去几十年间, 红藻真江蓠()已经从西北太平洋扩散到欧洲和北美河口的几乎所有高盐与温带地区。最新研究发现, 真江蓠在北美和欧洲的长距离定殖和随后的种群扩张与其较高的单性繁殖比例有关。在起源地西北太平洋, 几乎所有真江蓠的叶状体都是通过固着器附着在硬基质上, 而在入侵地大部分是自由漂浮无固着器的叶状体。这种起源地的硬基质和入侵地的软基质之间的单倍体-二倍体比率差异非常明显: 起源地种群二倍体比例平均为58%, 而在入侵地则为81%, 明显高于起源地(Krueger-Hadfield, 2016)。这可能是因为真江蓠的二倍叶状体比单倍体生长速度更快, 对环境胁迫的耐受力更强, 有助于在新的栖息地实现种群的快速建立。
2.3 冰期避难所与谱系时空演化
第四纪冰期和间冰期交替(始于约240万年前)引发的气候变化和海平面升降, 不仅改变了沿海的生态景观构造, 而且对生物多样性组成、丰度和分布模式等产生了重要影响(Hofreiter, 2009; Hu, 2016b)。末次盛冰期(the Last Glacial Maximum, 约1.8万—2.0万a前)是距今最近的一次冰期, 对现存物种地理分布和遗传格局影响较大。该时期北半球主要有三个大陆冰川中心: 欧亚大陆冰盖、劳伦泰德和格陵兰大陆冰盖以及西伯利亚大陆冰盖(Charbit, 2007)。与陆地生物相比, 冰盖的形成和消退对海洋生物地理分布的影响长期未得到足够的重视, 但越来越多的证据表明冰川活动对后者的影响程度超出了我们的认知(Maggs, 2008; Ni, 2014)。在冰盛期, 大陆冰盖一直延伸至海岸线, 造成海平面下降, 进而引起潮间带海洋生物大量灭绝。某些物种会被隔离并存留在特殊的区域即避难所内(如海沟、麻坑)。这些避难所不仅是生物在冰期逃避劫难的场所, 也是冰期后物种扩散和重新分布的起点与源库。受全球气候变暖的影响, 潮间带生物将逐步失去一些边缘的栖息地, 丢失许多独特的基因型和种质库(Provan,2012; Jueterbock, 2013; Zhang, 2019)。拓展与冰期避难所相关的研究主题(如避难所种群对环境变动的生态响应过程与机制, 冰期避难所种群的保护遗传学)不仅可丰富我们对种群适应力和生态恢复力的认知, 而且可提升物种对未来环境变化的反馈和保护潜力(Hampe, 2013; Hu, 2016b)。
在全球气候变化背景下, 确定和保护冰期避难所已成为指导生物多样性保护的首要任务(Keppel, 2012)。冰期避难所的数目与位置、冰期后物种的扩张时间与路径, 被认为是了解物种形成过程、成种事件和多样性保护的重要课题(Hewitt, 2004; Richardson, 2010)。化石、孢粉、古气候和古地质等数据在一定程度上有助于获悉动植物的潜在避难所和定居路线, 但对化石证据有限或缺乏的物种(如海洋植物)而言, 确定其避难所则比较困难。谱系地理学研究则为追溯这类生物的避难所形成和再扩散过程提供了强有力的工具。
当冰期来临时随着冰盖延伸, 北半球大多数海洋生物整体呈现由北向南迁移的趋势, 有些物种或种群会被隔离在避难所内, 长期的地理隔离会造成种间或种内的谱系分化。冰盖消退后, 避难所中的残存种群会向外扩散, 通常伴随着遗传漂变或奠基者效应, 使得重新定居种群的遗传多样性较低或基因型偏少。通常, 冰期避难所种群往往比重新定居的种群具有更长的进化历史, 因而积累了更高的遗传多样性和独特的基因型(Maggs, 2008)。因此, 遗传多样性是判断冰期避难所的一个重要指标。此外, 不同避难所种群由于地理隔离、祖先多态性的随机保留和杂交区(Hybridization zone)/接触区(Contact zone)出现等因素, 基因型的分布也会存在较大差异。因此, 基因型的空间分布模式是判断避难所的重要依据之一。
海洋植物是近岸生态系统的重要组成部分, 它们营固着生长, 迁移能力较弱, 在经历剧烈的环境变动后也能在当地保留相对完整的遗传信息。因此, 即使没有化石证据, 谱系地理学研究和古地质环境数据也可帮助推测海洋植物冰期避难所的位置。目前,海洋植物冰期避难所研究主要集中在北大西洋、西北太平洋、东南亚等热点地区(图5)。在北大西洋东岸, 海洋植物的潜在冰期避难所包括: 伊比利亚半岛、英吉利海峡(布列塔尼地区-Hurd deep海沟)、爱尔兰西南部、挪威沿岸、法罗群岛、非洲西北部(毛里塔尼亚和摩洛哥)、地中海(图5)。在北大西洋西岸, 海洋植物避难所主要位于加拿大沿岸(如新斯科舍和纽芬兰)。在西北太平洋, 目前报道了五个零散的冰期避难所: 日本海、日本-太平洋沿岸、东海(冲绳海槽)、韩国南部(济州岛)和台湾岛北部(图5)。值得提及的是, Hu等(2018)对在东南亚和中国南海广泛分布的匍枝马尾藻()进行了深入的谱系地理学研究, 首次提出中国海南岛西南部(莺歌海盆地的中央凹陷)存在一个以前未被认知的冰期避难所, 印度尼西亚的巴厘岛周围可能存在二级避难所(图5)。诚然谱系地理学为我们探讨冰期存留物种的进化历史提供了新的视角, 但单一证据不足以解释复杂的生物演化过程, 利用多源证据的综合方法(如物种分布模型), 整合多种学科的研究优势(如基于物理海洋学模型预测种群间扩散和基因流模式)将是未来的发展方向(Brennan, 2014; Assis, 2018)。
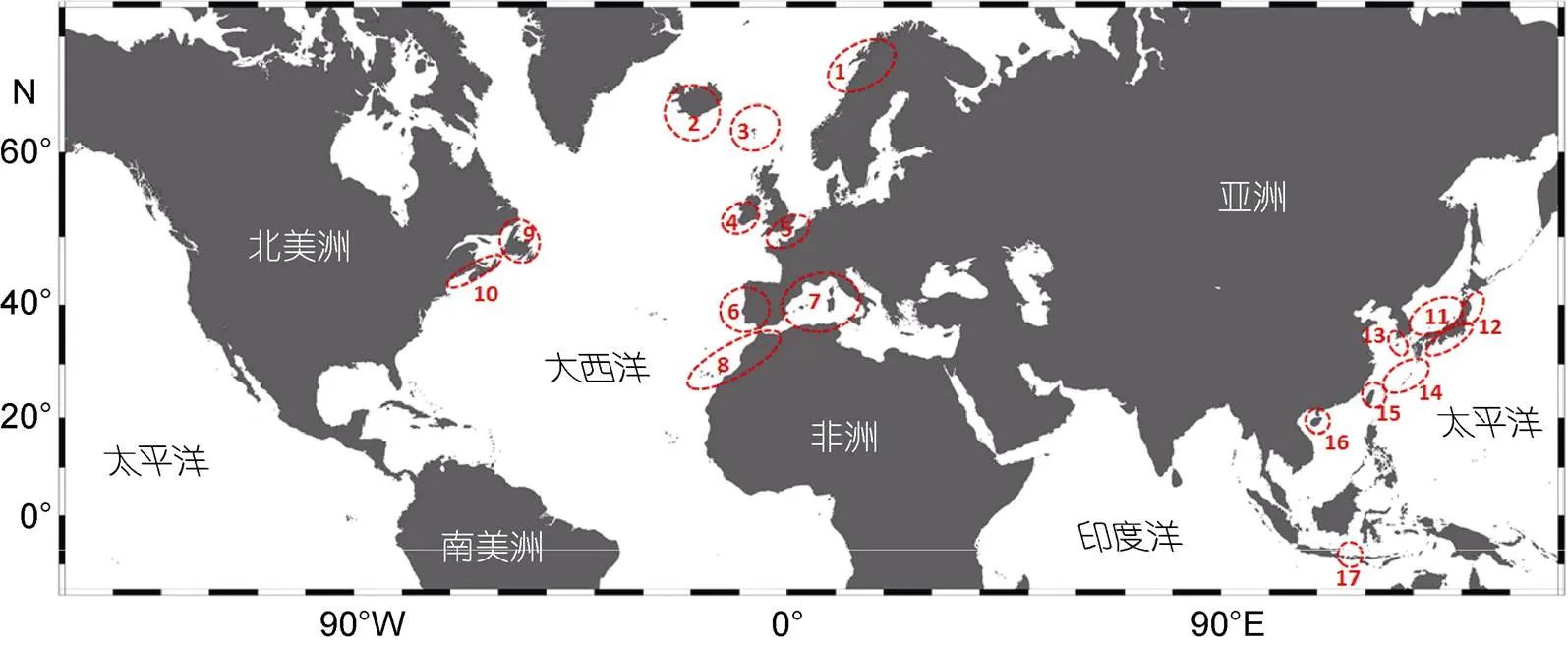
图5 全球范围内潜在的海洋植物冰期避难所位置
注: 1. 挪威北部; 2. 冰岛; 3. 法罗群岛; 4. 爱尔兰西南部; 5. 英吉利海峡; 6. 伊比利亚半岛; 7. 地中海沿岸; 8. 非洲西北部; 9. 纽芬兰省; 10. 新斯科舍省; 11. 日本海南部; 12. 日本-太平洋沿岸; 13. 韩国南部(包括济州岛); 14. 冲绳海槽; 15. 台湾岛北部; 16. 海南岛西南部; 17. 巴厘岛
2.4 克隆繁殖、新种形成与杂交
克隆繁殖是许多海洋植物拥有的一种常见生活史策略, 其特征是种群内某个或几个基因型呈现出极高的本地丰度。在某些受到胁迫或干扰的生境中, 利用克隆进行无性繁殖的植物种群可能代表其特殊的生境适应策略, 揭示这类种群的克隆遗传结构可深入了解海洋植物的生态适应与进化历史。在波罗的海, 墨角藻和种群的少量个体能以克隆方式进行无性繁殖(Tatarenkov, 2005), 而且(仅分布于波罗的海)是在末次冰盛期后由分布广阔的演化而来(Pereyra, 2009)。最近研究发现,中克隆繁殖的比例远高于, 并且在波罗的海北部(盐度低)更为常见。种群主要由少数几个克隆谱系组成, 其释放的配子池偏倚导致有效种群偏小, 而遗传漂变则进一步降低了谱系的遗传变异水平(Johannesson, 2011)。波罗的海的盐度从南往北逐步降低, 当盐度<10时释放的精子的活力和运动速度急剧下降, 当盐度<6—8时它的受精成功率也急剧降低(Serrão, 1996)。因此,在低盐的波罗的海北部难以进行有性繁殖, 其种群中大量克隆繁殖的出现是受低盐环境自然选择的结果。这种选择效应在波罗的海红藻中也有报道, 即其连续遗传变异分布和生态分化与该地区的盐度梯度变化相一致(Gabrielsen, 2002)。
遗传多样性的高低能影响海洋植物的适应性。克隆繁殖则导致有效种群大小降低和遗传漂变增加, 削减了种群的遗传多样性, 进而影响植物群落动态和生态系统稳定性。在受到干扰和非生物因素影响时, 克隆丰富度(Clonal richness)对海草群落作用显著。有研究发现, 当大叶藻()在冬季受到非生物和生物胁迫时, 克隆多样性与其枝芽密度呈正相关, 其作用程度与其他因素(如潮汐高度)相当, 对大叶藻的种群生物量产生重要影响(Hughes, 2009)。因此, 在开展海草资源保护和修复研究时, 有必要将遗传多样性纳入考虑范畴, 以提高其新的生境中的移植率和存活率。
许多海藻都具有微观的单倍体和宏观的二倍体阶段, 其配子融合和减数分裂在时间上完全隔离。减数分裂产生单倍体的配子体, 配子融合产生二倍体的孢子体, 单倍体和二倍体在整个生活史中交替出现。不同于单倍体占主导的高等动植物, 海藻的单倍体时期完全独立且具有完整功能, 能进行大量的体细胞发育(Valero, 1992), 这为研究海藻种间杂交和繁殖隔离机制提供了天然模型。杂交实验显示, 亲缘关系较远的水云[(♂) ×(♀)]种间可以发生胞质融合(plasmogamy), 但杂交体在萌发后大量死亡(Müller,1980)。最新的谱系遗传研究则显示, 同域分布的水云和可自然的进行杂交, 但在所有单倍体和二倍体中该比例只有8.7%, 且杂交体内含有亲本中缺乏的高频罕见等位基因。最重要的是, 大部分杂交发生在二倍体中(97.5%), 单倍体杂交比例只有2.5%, 说明生殖隔离主要发生在水云的合子形成后, 而非合子形成前(Montecinos, 2017)。基因组大小不同的物种杂交后在减数分裂过程中发生了染色体异常分离(染色体重组), 这可能是导致水云不同种之间杂交不亲和的主要原因。
3 海洋植物谱系地理学研究展望
3.1 基于海洋植物的岛屿生物学模型检验
海岛大多远离陆地, 是研究多样性进化模式和生物地理过程(如迁移、灭绝、物种形成)的天然优良系统。岛屿生物学理论由麦克阿瑟和威尔逊于20世纪70年代创立(MacArthur, 1967), 该理论经过四十年的发展, 其要旨大致包括: 大的海岛拥有大的种群和更高的环境异质性, 因此物种灭绝概率较低, 本地物种丰度较高; 隔离的岛屿接受迁入的个体较少, 导致物种丰度降低; 弱基因流导致岛屿上本地种数量变丰富; 岛屿越古老, 它更有可能接受物种迁入, 新物种形成的概率增加, 岛屿上物种数量和本地种丰度也越高(Whittaker, 2007)。
最新研究发现, 在偏僻岛屿上原地(in situ)发生的进化动力学过程使得物种-面积关系(Species-area relationships)明显偏离麦克阿瑟和威尔逊最初提出的经典模式。岛屿递进法则(Island progression rule)主张古老的岛屿作为起源(source)向新生岛屿贡献了新的侵殖物种(colonized species), 促成新生岛屿上多样性的汇聚(sink)(Whittaker, 2017)。群岛的各海岛间繁殖体流动的差异对于调节新的多样性地理分布至关重要。许多缺乏特殊扩散方式和适应能力的植物种类可通过非常规的传播方式(如人类活动)侵殖偏僻岛屿(Whittaker, 2017)。在南大西洋的海脊和岛屿上, 大多数本地珊瑚礁鱼类在更新世发生演化, 岛屿上鱼类的物种形成速率与移居(immigration)速率呈负相关。扩散能力弱的鱼类在岛屿上随机积累加速了物种形成, 而在原地支系形成和适应性辐射的概率降低。此外, 海平面波动和海山位置间歇性地为岛屿侵殖提供了基石, 对海洋物种进化起到重要推动作用(Pinheiro, 2017)。这些新的发现极大的推动了岛屿生物地理学理论的发展和完善。
尽管如此, 岛屿生物地理模式因物种所处生态系统的不同(如浅海vs.深海)而异, 即使在同一系统内不同物种间也存在差异。Hachich等(2015)发现在大西洋浅海生态系统内, 岛屿面积最适于解释腹足类和海藻的物种丰度, 岛屿间的隔离可用于解释海藻的物种丰度, 但不适于珊瑚礁鱼类和腹足类, 而岛屿年龄可以解释所有研究类群的物种丰度。此外, 扩散能力可显著影响岛屿上的物种多样性, 使得海藻和腹足类(扩散能力较弱)与珊瑚礁鱼类(较强)的物种丰度受岛屿隔离的影响程度存在显著差异(Hachich, 2016)。这是迄今为数不多的涉及海藻的岛屿生物地理模型检验研究, 也凸显海藻的特殊生物学属性及利用其检验岛屿生物地理模式的独特优势。未来, 海藻在揭示特殊的岛屿生物地理模式方面可能会发挥重要作用, 包括一些当前的研究热点(如是否存在物种从岛屿向陆地的反向侵殖?)。
3.2 基于海洋植物-微生物群落互作的功能谱系地理学
海藻与其表面附生的微生物群落互作共利, 形成动态的生态平衡。对于某些海藻, 其形态建成、谱系分布和多样性组成与表面附生菌落种类及数量密切相关。褐藻海带()在深水区(14—25 m)和浅水区(5—11 m)的表型差异与遗传变异显著(Balakirev, 2012a),但不同水层的海带表面附生微生物群落种类则与深度无关(Balakirev, 2012b)。表面附生微生物在不同水层的转移可能导致两个海带世系间发生表型可塑, 并引发生殖隔离。此外, 通过对三种石莼(spp.)附生细菌群落对比, 发现30%的富集功能基因可能与局域适应或宿主特异适应有关, 表明具有不同系统发育起源的微生物多样性有助于宿主/栖息地特异性基因发挥其功能, 进而影响宿主的核心基因功能(Roth-Schulze, 2018)。
目前, 在入侵过程中真核生物宿主和互利共生者间的关系共有三个主要假说: 广义宿主假说(Generalist host hypothesis), 即入侵种的宿主来源相对较为广泛; 增强互利假说(Enhanced mutualism hypothesis), 即在引入种和本地种之间新的互利建立; 伴随互利假说(Accompanying mutualist hypothesis), 即入侵宿主与互利共生者共同引入。同时分析入侵种的遗传库及相关细菌区系的进化多样性, 不仅可在起源地和入侵地检验这些假设, 而且可确定最可能的引入途径和机制, 甄别宿主和细菌之间的相互作用机制, 更好的理解引入种何以能成功变为入侵种。最新的共生体(holobiont, 即海藻与内生细菌群落)功能基因组学研究发现, 全球范围内不同地区杉叶蕨藻基因组与内生菌群落的α多样性(每个取样点和每个取样单元内Shannon多样性指数的标准值)和β多样性(谱系间距和样品间谱系类型的频率)高度一致且密切相关, 支持了共生体的概念。海藻和内生菌的两组基因组数据支持地中海地区杉叶蕨藻单次入侵和澳大利亚南部多次入侵的假设。内生菌群落的代谢功能可能有利于增强共生体的适应能力和竞争能力。因此, 共生体(特别是内生菌)遗传多样性的降低可能导致杉叶蕨藻最近在地中海入侵范围的缩减(Arnaud-Haond, 2017)。这些研究是大型海藻-微生物群落互作的典型实例, 突显宿主-寄主共同进化是提升共生体适应能力和拓殖分布空间的重要因素。因此, 对海洋植物-微生物群落展开谱系遗传结构和生态位转换的协同调查研究, 可从群落和生态系统功能进化层面重新认识海洋生物的多样性结构与适应进化。
3.3 基于基因组扫描的海洋植物隐存多样性挖掘和进化模式解析
海洋植物蕴含丰富的物种多样性和地理演化模式, 但迄今有关的理论发现非常有限, 原因之一是当前使用的分子标记分辨力较低, 难以从基因组尺度对多态变异信息进行完整扫描。历史资料显示红藻属的地理分布范围仅限于菲律宾, 且只有1—3个种。基于多基因位点的物种定界和系统发育研究显示该属在菲律宾群岛至少存在21个隐存的进化单元(Payo, 2013), 这些分布在不同岛屿且高度分化的隐存世系凸显了第四纪冰期气候变化期间微地理环境变量(micro-geographic environmental variables)对海藻物种多样性演化的驱动作用。最近, 基于6个标记序列(3782 bp)的研究进一步发现该属内存在92个隐存的进化单元, 它们大多数扩散能力弱, 分布区狭窄; 该属在白垩纪末期起源于印太的中部地区, 印度-马来群岛(Indo-Malay Archipelago)的悠久地质历史塑造了属的物种多样性(Leliaert, 2018)。鉴于此, 如果在海洋植物谱系多样性挖掘和进化模式研究中引入高分辨率的检测技术(如Restriction-site Associated DNA, RAD), 其提供的数以万计的SNP位点可能会颠覆我们对海洋植物物种丰度和多样性进化历史的认知。
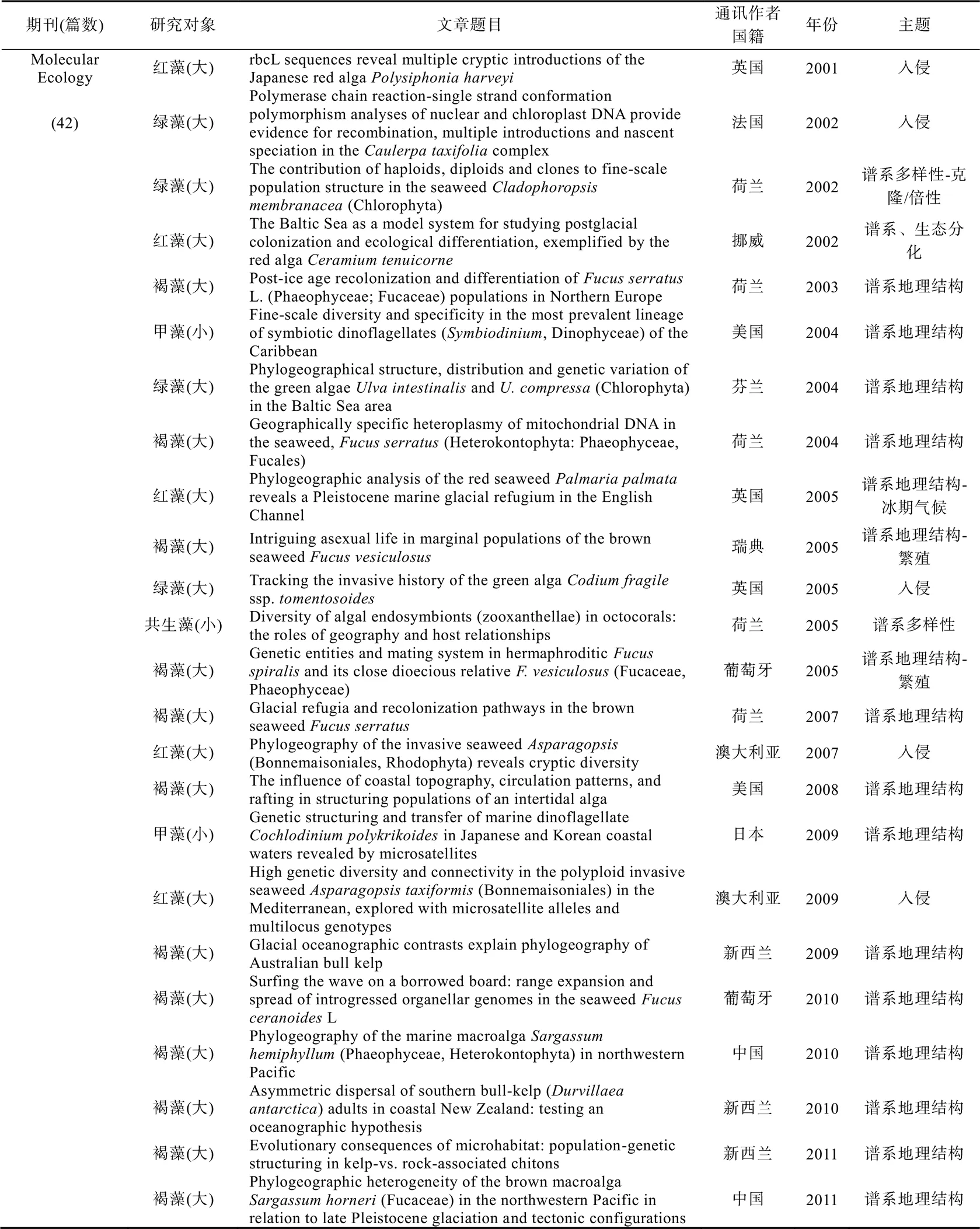
附录 2000—2019年间《Molecular Ecology》和《Journal of Biogeography》发表的海藻和海草谱系地理学论文及有关信息
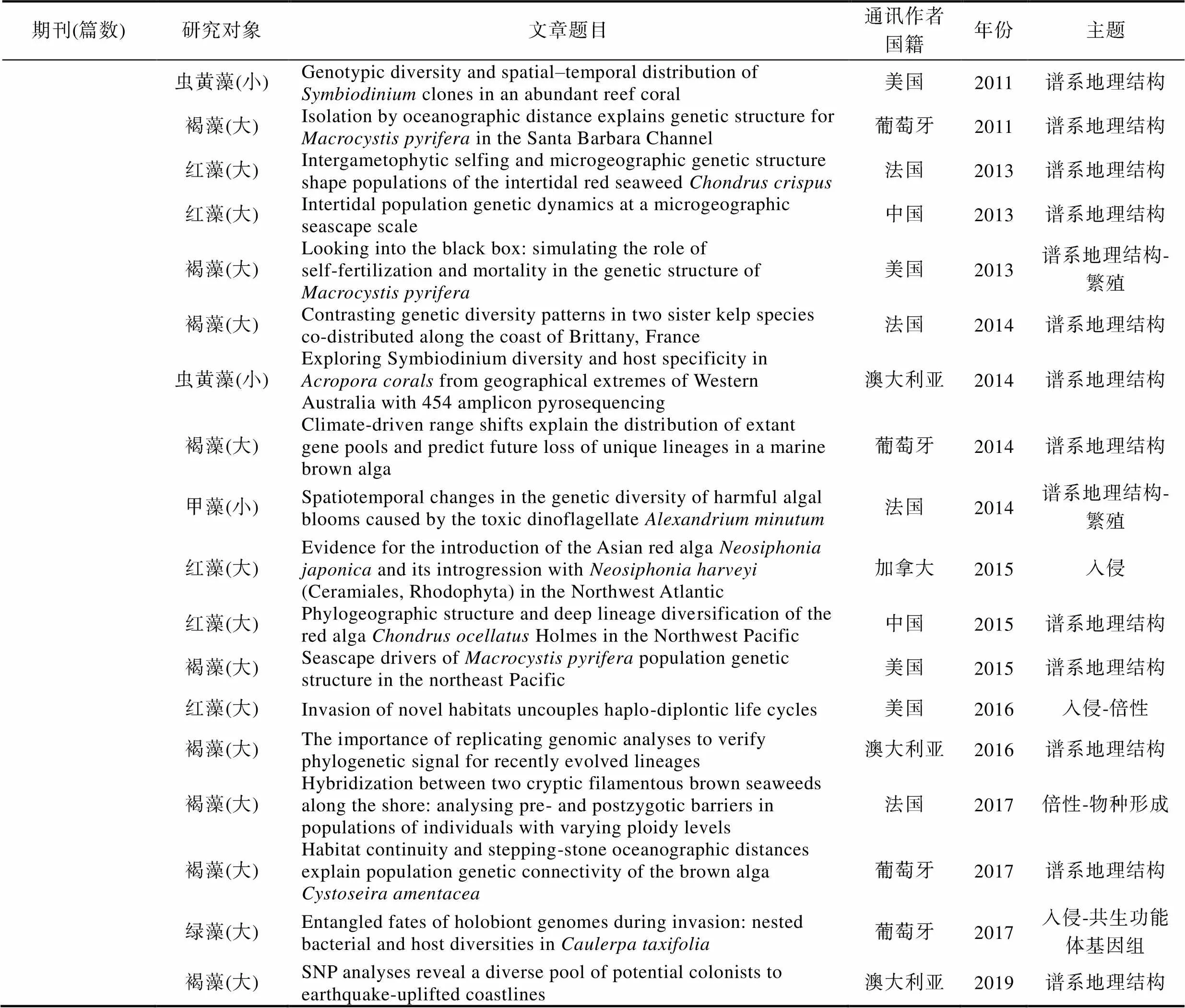
期刊(篇数)研究对象文章题目通讯作者国籍年份主题 虫黄藻(小)Genotypic diversity and spatial–temporal distribution of Symbiodinium clones in an abundant reef coral美国2011谱系地理结构 褐藻(大)Isolation by oceanographic distance explains genetic structure for Macrocystis pyrifera in the Santa Barbara Channel葡萄牙2011谱系地理结构 红藻(大)Intergametophytic selfing and microgeographic genetic structure shape populations of the intertidal red seaweed Chondrus crispus法国2013谱系地理结构 红藻(大)Intertidal population genetic dynamics at a microgeographic seascape scale中国2013谱系地理结构 褐藻(大)Looking into the black box: simulating the role of self-fertilization and mortality in the genetic structure of Macrocystis pyrifera美国2013谱系地理结构-繁殖 褐藻(大)Contrasting genetic diversity patterns in two sister kelp species co-distributed along the coast of Brittany, France法国2014谱系地理结构 虫黄藻(小)Exploring Symbiodinium diversity and host specificity in Acropora corals from geographical extremes of Western Australia with 454 amplicon pyrosequencing澳大利亚2014谱系地理结构 褐藻(大)Climate-driven range shifts explain the distribution of extant gene pools and predict future loss of unique lineages in a marine brown alga葡萄牙2014谱系地理结构 甲藻(小)Spatiotemporal changes in the genetic diversity of harmful algal blooms caused by the toxic dinoflagellate Alexandrium minutum法国2014谱系地理结构-繁殖 红藻(大)Evidence for the introduction of the Asian red alga Neosiphonia japonica and its introgression with Neosiphonia harveyi (Ceramiales, Rhodophyta) in the Northwest Atlantic加拿大2015入侵 红藻(大)Phylogeographic structure and deep lineage diversification of the red alga Chondrus ocellatus Holmes in the Northwest Pacific中国2015谱系地理结构 褐藻(大)Seascape drivers of Macrocystis pyrifera population genetic structure in the northeast Pacific美国2015谱系地理结构 红藻(大)Invasion of novel habitats uncouples haplo-diplontic life cycles美国2016入侵-倍性 褐藻(大)The importance of replicating genomic analyses to verify phylogenetic signal for recently evolved lineages澳大利亚2016谱系地理结构 褐藻(大)Hybridization between two cryptic filamentous brown seaweeds along the shore: analysing pre- and postzygotic barriers in populations of individuals with varying ploidy levels法国2017倍性-物种形成 褐藻(大)Habitat continuity and stepping-stone oceanographic distances explain population genetic connectivity of the brown alga Cystoseira amentacea葡萄牙2017谱系地理结构 绿藻(大)Entangled fates of holobiont genomes during invasion: nested bacterial and host diversities in Caulerpa taxifolia葡萄牙2017入侵-共生功能体基因组 褐藻(大)SNP analyses reveal a diverse pool of potential colonists to earthquake-uplifted coastlines澳大利亚2019谱系地理结构
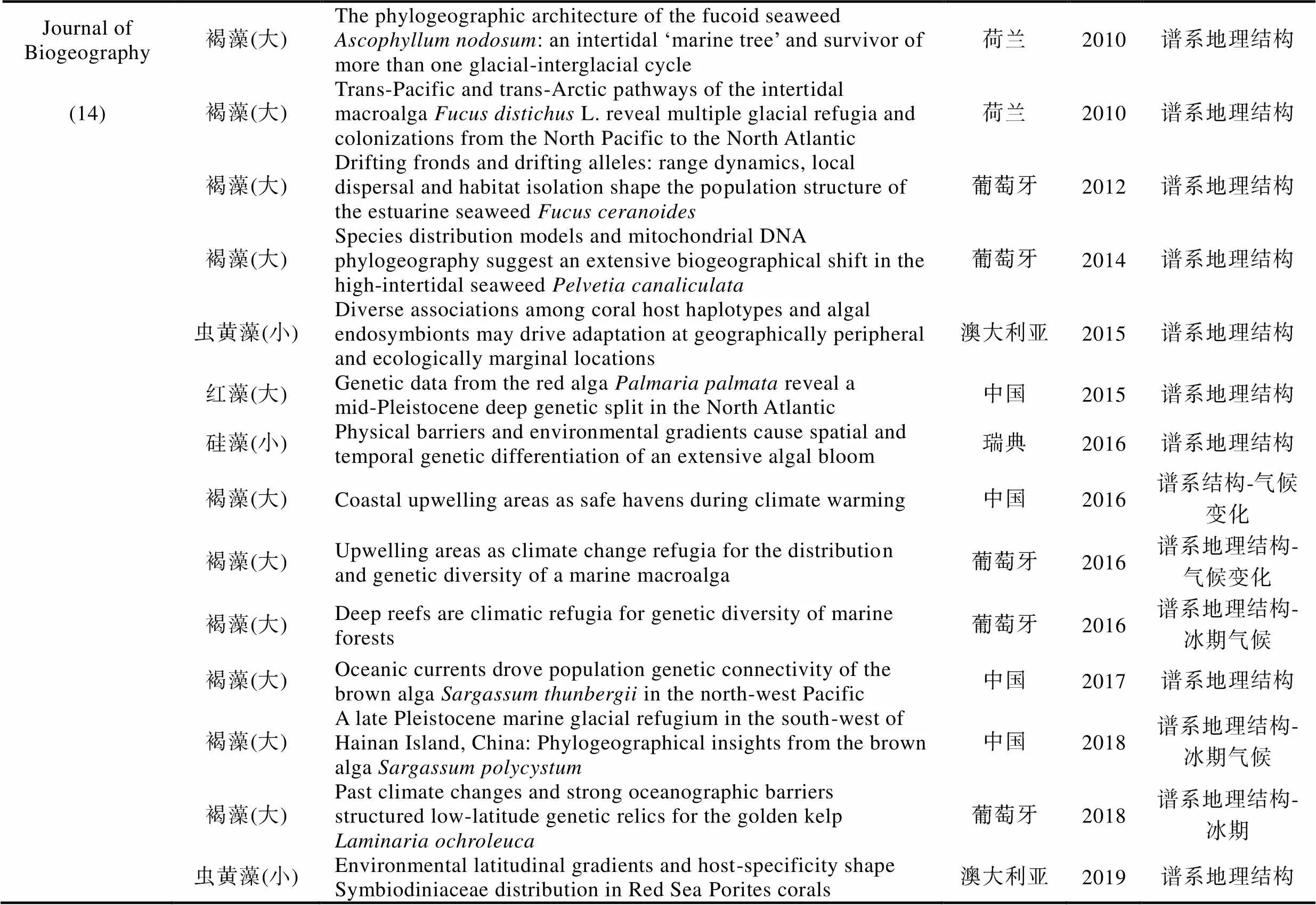
续表
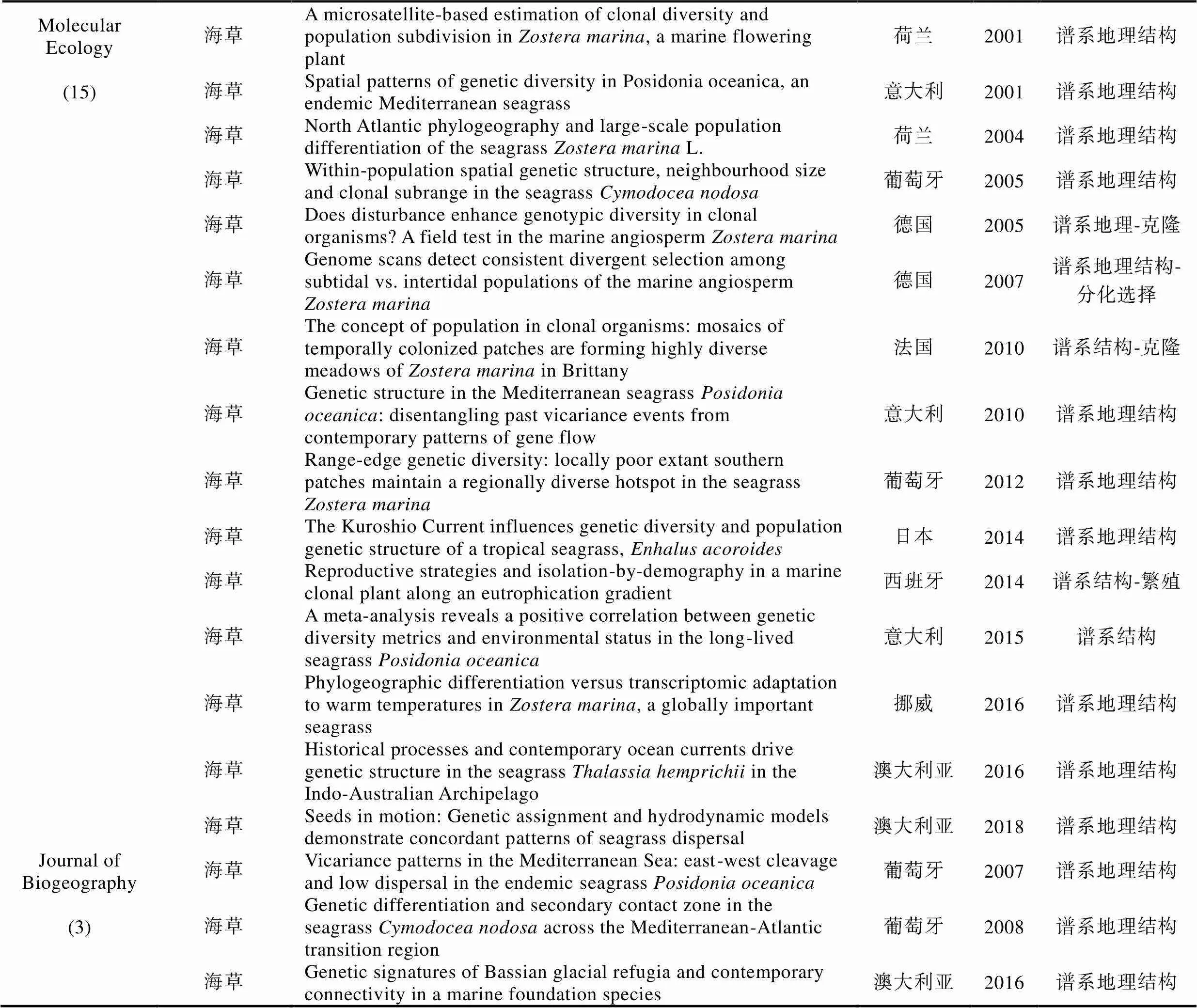
续表
Alberto F, Raimondi P T, Reed D C, 2011. Isolation by oceanographic distance explains genetic structure forin the Santa Barbara Channel. Molecular Ecology, 20(12): 2543—2554
Andreakis N, Kooistra W H C F, Procaccini G, 2009. High genetic diversity and connectivity in the polyploid invasive seaweed(Bonnemaisoniales) in the Mediterranean, explored with microsatellite alleles and multilocus genotypes. Molecular Ecology, 18(2): 212—216
Andreakis N, Procaccini G, Maggs C, 2007. Phylogeography of the invasive seaweed(Bonnemaisoniales, Rhodophyta) reveals cryptic diversity. Molecular Ecology, 16(11): 2285—2299
Arnaud-Haond S, Aires T, Candeias R, 2017. Entangled fates of holobiont genomes during invasion: nested bacterial and host diversities in. Molecular Ecology, 26(8): 2379—2391
Assis J, Serrao E Á, Coelho N C, 2018. Past climate changes and strong oceanographic barriers structured low-latitude genetic relics for the golden kelp. Journal of Biogeography, 45(10): 2326—2336
Avise J C, 2000. Phylogeography: The History and Formation of Species. Cambridge, Massachusetts: Harvard University Press
Avise J C, Arnold J, Ball R M, 1987. Intraspecific phylogeography: the mitochondrial DNA bridge between population genetics and systematics. Annual Review of Ecology and Systematics, 18: 489—522
Balakirev E S, Krupnova T N, Ayala F J, 2012a. DNA variation in the phenotypically-diverse brown alga. BMC Plant Biology, 12(1): 108
Balakirev E S, Krupnova T N, Ayala F J, 2012b. Symbiotic associations in the phenotypically-diverse brown alga. PLoS One, 7(6): e39587
Beheregaray L B, 2008. Twenty years of phylogeography: the state of the field and the challenges for the Southern Hemisphere. Molecular Ecology, 17(17): 3754—3774
Blossey B, Notzold R, 1995. Evolution of increased competitive ability in invasive nonindigenous plants: a hypothesis. Journal of Ecology, 83(5): 887—889
Brennan G, Kregting L, Beatty G E, 2014. Understanding macroalgal dispersal in a complex hydrodynamic environment: a combined population genetic and physical modelling approach. Journal of the Royal Society Interface, 11(95): 20140197
Cabello-Pasini A, Muñiz-Salazar R, Ward D H, 2003. Annual variations of biomass and photosynthesis inat its southern end of distribution in the North Pacific. Aquatic Botany, 76(1): 31—47
Cabello-Pasini A, Muñiz-Salazar R, Ward D H, 2004. Biochemical characterization of the eelgrassat its southern distribution limit in the North Pacific. Ciencias Marinas, 30(1A): 21—34
Charbit S, Ritz C, Philippon G, 2007. Numerical reconstructions of the Northern Hemisphere ice sheets through the last glacial-interglacial cycle. Climate of the Past, 3(1): 15—37
Gabrielsen T M, Brochmann C, Rueness J, 2002. The Baltic Sea as a model system for studying postglacial colonization and ecological differentiation, exemplified by the red alga. Molecular Ecology, 11(10): 2083—2095
Hachich N F, Bonsall M B, Arraut E M, 2015. Island biogeography: patterns of marine shallow-water organisms in the Atlantic Ocean. Journal of Biogeography, 42(10): 1871—1882
Hachich N F, Bonsall M B, Arraut E M, 2016. Marine island biogeography. Response to comment on ‘Island biogeography: patterns of marine shallow-water organisms’. Journal of Biogeography, 43(12): 2517—2519
Hampe A, Rodríguez-Sánchez F, Dobrowski S, 2013. Climate refugia: from the last glacial maximum to the twenty-first century. New Phytologist, 197(1): 16—18
Hewitt G M, 2004. The structure of biodiversity——insights from molecular phylogeography. Frontiers in Zoology, 1(1): 4
Hoareau T B, Boissin E, Paulay G, 2013. The Southwestern Indian Ocean as a potential marine evolutionary hotspot: perspectives from comparative phylogeography of reef brittle-stars. Journal of Biogeography, 40(11): 2167—2179
Hofreiter M, Stewart J, 2009. Ecological change, range fluctuations and population dynamics during the Pleistocene. Current Biology, 19(14): R584—R594
Hu Z M, Duan D L, Lopez-Bautista J, 2016a. Seaweed phylogeography from 1994 to 2014: an overview. In: Hu Z M, Fraser C eds. Seaweed Phylogeography: Adaptation and Evolution of Seaweeds Under Environmental Change. Dordrecht: Springer, 3—22
Hu Z M, Fraser C, 2016b. Seaweed Phylogeography: Adaptation and Evolution of Seaweeds under Environmental Change. The Netherlands: Springer
Hu Z M, Kantachumpoo A, Liu R Y, 2018. A late Pleistocene marine glacial refugium in the South-West of Hainan Island, China: phylogeographical insights from the brown alga. Journal of Biogeography, 45(2): 355—366
Hughes A R, Stachowicz J J, 2009. Ecological impacts of genotypic diversity in the clonal seagrass. Ecology, 90(5): 1412—1419
Johannesson K, Johansson D, Larsson K H, 2011. Frequent clonality in fucoids (and: Fucales, Phaeophyceae) in the Baltic Sea. Journal of Phycology, 47(5): 990—998
Jormalainen V, Danelli M, Gagnon K, 2017. Genetic variation of a foundation rockweed species affects associated communities. Ecology, 98(11): 2940—2951
Jueterbock A, Tyberghein L, Verbruggen H, 2013. Climate change impact on seaweed meadow distribution in the North Atlantic rocky intertidal. Ecology and Evolution, 3(5): 1356—1373
Keppel G, Van Niel K P, Wardell-Johnson G W, 2012. Refugia: Identifying and understanding safe havens for biodiversity under climate change. Global Ecology and Biogeography, 21(4): 393—404
Krueger-Hadfield S A, Kollars N M, Byers J E, 2016. Invasion of novel habitats uncouples haplo-diplontic life cycles. Molecular Ecology, 25(16): 3801—3816
Leliaert F, Payo D A, Gurgel C F D, 2018. Patterns and drivers of species diversity in the Indo-Pacific red seaweed. Journal of Biogeography, 45(10): 2299—2313
Li J J, Hu Z M, Gao X, 2017. Oceanic currents drove population genetic connectivity of the brown algain the North-West Pacific. Journal of Biogeography, 44(1): 230—242
MacArthur R H, Wilson E O, 1967. The Theory of Island Biogeography. Princeton, NJ: Princeton University Press
Maggs C A, Castilho R, Foltz D, 2008. Evaluating signatures of glacial refugia for North Atlantic benthic marine taxa. Ecology, 89(sp11): S108—S122
Manel S, Schwartz M K, Luikart G, 2003. Landscape genetics: combining landscape ecology and population genetics. Trends in Ecology & Evolution, 18(4): 189—197
McIvor L, Maggs C A, Provan J, 2001.L sequences reveal multiple cryptic introductions of the Japanese red alga. Molecular Ecology, 10(4): 911—919
Meusnier I, Olsen J L, Stam W T, 2001. Phylogenetic analyses of(Chlorophyta) and of its associated bacterial microflora provide clues to the origin of the Mediterranean introduction. Molecular Ecology, 10(4): 931—946
Meusnier I, Valero M, Destombe C, 2002. Polymerase chain reaction-single strand conformation polymorphism analyses of nuclear and chloroplast DNA provide evidence for recombination, multiple introductions and nascent speciation in thecomplex. Molecular Ecology, 11(11): 2317—2325
Mitarai S, Siegel D A, Watson J R, 2009. Quantifying connectivity in the coastal ocean with application to the Southern California Bight. Journal of Geophysical Research: Oceans, 114(C10): C10026
Montecinos A E, Guillemin M L, Couceiro L, 2017. Hybridization between two cryptic filamentous brown seaweeds along the shore: analysing pre- and postzygotic barriers in populations of individuals with varying ploidy levels. Molecular Ecology, 26(13): 3497—3512
Muhlin J F, Engel C R, Stessel R, 2008. The influence of coastal topography, circulation patterns, and rafting in structuring populations of an intertidal alga. Molecular Ecology, 17(5): 1198—1210
Muñiz-Salazar R, Talbot S L, Sage G K, 2005. Population genetic structure of annual and perennial populations ofL. along the Pacific coast of Baja California and the Gulf of California. Molecular Ecology, 14(3): 711—722
Müller D G, Gassmann G, 1980. Sexual hormone specificity inand(Phaeophyceae). Naturwissenschaften, 67(9): 462—463
Ni G, Li Q, Kong LF, 2014. Comparative phylogeography in marginal seas of the northwestern Pacific. Molecular Ecology, 23(3): 534—548
Palumbi S R, 1994. Genetic divergence, reproductive isolation, and marine speciation. Annual Review of Ecology and Systematics, 25: 547—572
Payo D A, Leliaert F, Verbruggen H, 2013. Extensive cryptic species diversity and fine-scale endemism in the marine red algain the Philippines. Proceedings of the Royal Society B: Biological Sciences, 280(1753): 20122660
Pereyra R T, Bergström L, Kautsky L, 2009. Rapid speciation in a newly opened postglacial marine environment, the Baltic Sea. BMC Evolutionary Biology, 9(1): 70
Pinheiro H T, Bernardi G, Simon T, 2017. Island biogeography of marine organisms. Nature, 549(7670): 82—85
Provan J, Maggs C A, 2012. Unique genetic variation at a species’ rear edge is under threat from global climate change. Proceedings of the Royal Society B: Biological Sciences, 279(1726): 39—47
Provan J, Murphy S, Maggs C A, 2005. Tracking the invasive history of the green algassp.. Molecular Ecology, 14(1): 189—194
Richardson D M, Whittaker R J, 2010. Conservation biogeography — foundations, concepts and challenges. Diversity and Distribution, 16(3): 313—320.
Roth-Schulze A J, Pintado J, Zozaya-Valdés E, 2018. Functional biogeography and host specificity of bacterial communities associated with the marine green algaspp. Molecular Ecology, 27(8): 1952—1965
Savoie A M, Saunders G W, 2015. Evidence for the introduction of the Asian red algaand its introgression with(Ceramiales, Rhodophyta) in the Northwest Atlantic. Molecular Ecology, 24(23): 5927—5937
Serrão E A, Kautsky L, Brawley S H, 1996. Distributional success of the marine seaweedL. in the brackish Baltic Sea correlates with osmotic capabilities of Baltic gametes. Oecologia, 107(1): 1—12
Tatarenkov A, Bergström L, Jönsson R B, 2005. Intriguing asexual life in marginal populations of the brown seaweed. Molecular Ecology, 14(2): 647—651
Valero M, Richerd S, Perrot V, 1992. Evolution of alternation of haploid and diploid phases in life cycles. Trends in Ecology & Evolution, 7(1): 25—29
Wernberg T, Coleman M A, Bennett S, 2018. Genetic diversity and kelp forest vulnerability to climatic stress. Scientific Reports, 8(1): 1851
White C, Selkoe K A, Watson J, 2010. Ocean currents help explain population genetic structure. Proceedings of the Royal Society B: Biological Sciences, 77(1688): 1685—1694
Whittaker R J, Fernández-Palacios J M, 2007. Island Biogeography: Ecology, Evolution, and Conservation. Oxford: Oxford University Press
Whittaker R J, Fernández-Palacios J M, Matthews T J, 2017. Island biogeography: taking the long view of nature’s laboratories. Science, 357(6354): eaam8326
Zhang J, Yao J T, Hu Z M, 2019. Phylogeographic diversification and postglacial range dynamics shed light on the conservation of the kelp. Evolutionary Applications, 12(4): 791—803
PHYLOGEOGRAPHIC PATTERNS AND GENETIC CONNECTIVITY OF MARINE PLANTS: A REVIEW
HU Zi-Min1, 2, DU Yu-Qun1, 3, LIANG Yan-Shuo1, 3, ZHONG Kai-Le1, 3, ZHANG Jie1, 2
(1. Key Laboratory of Experimental Marine Biology, Center for Ocean Mega-Science, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China; 2. Laboratory for Marine Biology and Biotechnology, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao 266237, China; 3. University of Chinese Academy of Sciences, Beijing 100049, China)
Marine plant phylogeography is a relatively minority research area with no more than 30 years, but it has made considerable contribution to the understanding of how environmental changes altered marine species diversity patterns, ecological adaptation, and evolutionary capability across time and space. We firstly searched and screened papers on the marine plant phylogeography published in two most well-known international journalsand, and then summarized the main tendency and research status of marine plant phylogeography during 2000—2019 in terms of country/region of the author(s) and taxonomic position of research objectives in the world. We subsequently reviewed the mechanistic roles of ocean currents in shaping phylogeographic structure and diversity of marine plants, population genetic variation and reconstruction of colonization routes of invasive species, the genetic imprints of glacial refugia in extant genetic connectivity and geographic shifts of marine plants, and the contribution of clonal reproduction to phylogeographic diversification. Finally, this article forecasts the future research directions of marine plant phylogeography in terms of incorporating island biogeographic hypotheses, phylogeographic co-evolution of symbiotic plants and microbiome, and the application of high-throughput sequencing data to thoroughly understand diversity, distribution, and evolution of marine plants.
seaweed; seagrass; climate change; glacial refugia; invasion genetics; genetic diversity conservation
* 国家自然科学基金面上项目, 31971395号; 中国科学院战略性先导科技专项(B类)子课题, XDB42030203号。胡自民, 博士, 研究员, E-mail: huzimin9712@163.com
张 杰, 博士, 助理研究员, E-mail: zhangjie0616@qdio.ac.cn
2020-06-18,
2020-08-09
Q178.532
10.11693/hyhz20200600168
