镉胁迫下巨桉幼树的生长、养分含量变化及镉富集特性
2021-03-29刘喜建胡红玲胡庭兴张成浩铁得祥代大川张雪媛
刘喜建,陈 刚,胡红玲,胡庭兴,张 建,张成浩,铁得祥,周 鑫,代大川,张雪媛
(1四川农业大学 林学院,四川 成都 611130;2 四川省林业勘察设计研究院,四川 成都 610036)
当前,随着各类废弃物的不断排放,重金属污染逐步加重[1],目前我国已有近1/5耕地遭受多种重金属不同程度的污染[2]。镉(Cd)作为造成土壤污染最严重的重金属元素之一,具有隐蔽性、长期性和不可逆转性等特点,不仅能导致土壤生产力下降,还会进入整个生态系统,并通过食物链进入人体而危害人类健康[3]。有研究表明,一定量的Cd胁迫能降低土壤养分有效性、改变植物细胞质膜透性[4],从而降低植物对N、P、K的吸收、转运与分配能力[5],导致植物体内营养元素缺乏,生理代谢紊乱,同时外部呈现相应的缺素症状[6],进而抑制植物生长,甚至改变生物量的分配格局。
植物修复是指利用重金属富集能力强的植物吸附土壤中的重金属,达到降低土壤重金属浓度、修复生态环境的目的[7]。与物理修复和化学修复相比,植物修复能在不破坏土壤生态环境的条件下进行修复且不易造成二次污染,此技术的关键是正确选择修复植物[8]。近年来有关Cd污染植物修复的研究报道颇多,但其研究对象多为草本植物,如宝山堇菜(Violabaoshanensis)[9]、东南景天(Sedumalfredii)[10]等,而这些植物因生物量小、根系分布浅等缺点,对重金属吸收存在一定的局限性[11]。
桉树(Eucalyptusrobusta)为高大乔木,具有生长快、产量高、适应性强、根系分布广等特点,目前在全球热带、亚热带地区已广泛引种栽培,其种植面积超过2 000万hm2,占世界人工林面积的23%,而我国是桉树人工林栽植面积最大的国家之一[12]。有研究表明,桉树能够在重金属污染地区生长,并对Cd具有一定的富集作用[13]。巨桉(Eucalyptusgrandis)具有速生丰产的特点和优良的造纸性能,近20年来,我国南方地区在退耕还林、防护林营造等生态工程建设和农业产业结构调整中,营造了大量的巨桉人工林,因此研究重金属Cd对巨桉生长和生理特性的影响以及巨桉对Cd的耐受性,分析Cd元素在巨桉各组织器官内的分配和富集特性,探讨巨桉修复Cd污染土壤的可能性,具有一定的现实意义。但目前针对Cd胁迫下巨桉幼树的研究鲜有报道。本研究采用盆栽试验,探究巨桉幼树在不同程度Cd胁迫下的生长状况、养分分配及Cd富集特性,以期为耐Cd树种选择及Cd污染土壤的植物修复提供参考。
1 材料与方法
1.1 试验地概况
试验地位于四川农业大学(成都校区)教学科研试验基地,地理位置为东经103°51′29″,北纬30°42′18″,属亚热带湿润气候区,四季分明,气候温和,雨量充沛。年度平均气温为16.8 ℃,月平均最高温为25.9 ℃(7月),月平均最低温为6.9 ℃(12月),极端高温38 ℃(7月),极端低温-2 ℃(1月);年均相对湿度84%;年降水量为759.1~1 155.0 mm;年均日照时数为840.2 h。
1.2 试验材料
1.2.1 土壤与容器 试验用土取自四川雅安,为坡积土,其理化性质为:pH 6.5,全氮15.06 g/kg,全磷1.69 g/kg,全钾12.10 g/kg,Cd 0.72 mg/kg。栽植容器采用规格一致的塑料盆(盆口直径30 cm,底径25 cm,高26 cm),每盆装土8 kg(干土质量)。
1.2.2 供试植株 供试植株为健康的巨桉百日苗,每盆栽植1株,巨桉为速生树种,为避免Cd胁迫期间土壤中缺乏营养元素影响处理植株的生长,从而干扰胁迫效果,参照当地巨桉百日苗造林经验和文献[14-15],首先在2017年12月进行土壤装盆时,每盆添加15 g氮磷钾复合肥作为基肥;栽植之后分别于1月中旬、2月中旬和4月中旬3次追施氮磷钾复合肥。第1次施肥量为2 g/株(植株尚小,株高15 cm),第2次施肥量为4 g/株(株高50 cm),第3次施肥量为5 g/株(株高85 cm);追肥方式为:以盆口边缘距植株地径13 cm为半径,沿盆口用小铁锹开3 cm深环形沟,将肥料沿沟均匀施入,然后覆土并适量浇水。对巨桉苗进行为期5个月的适应性培养,期间定期采用HH2土壤水分测定仪(ML2x,GBR)监测土壤水分状况,使土壤体积含水量保持在18%左右(即田间持水量的70%~80%)。正式试验开始前15 d,所有处理植株均停止施肥,此时巨桉幼树苗龄接近1年,平均株高(86.3±4.2)cm,平均地径(7.98±0.96)mm。
1.3 试验设计
根据调查数据,四川土壤平均Cd含量为0.86 mg/kg,Cd污染最严重的区域可达22.2 mg/kg[16]。为预测未来重金属排放持续增加可能加重对植物的危害,并考虑到巨桉速生且生长季节长,植株个体大的特点,如果试验设置的Cd胁迫含量较低则其对巨桉幼树的胁迫效果可能不明显,故本试验设置了2个高含量(50,100 mg/kg)Cd处理做参考。本试验设置的Cd胁迫含量分别为:0 mg/kg(CK),12.5 mg/kg(T1),25 mg/kg(T2),50 mg/kg(T3)和100 mg/kg(T4),T1~T4处理氯化镉(CdCl2)施入量分别为0,25.4,50.8,101.6和203.2 mg/kg,每处理5次重复。试验于2018年5月初开始,为使Cd施入方式与Cd排放以及植物根系吸收过程相似,将设计的氯化镉施入量等分为5份,分别于2018年5,6,7,8,9月分5次施入,每盆用500 mL去离子水将CdCl2配制成溶液,均匀浇灌在盆内土壤表面,盆底托盘中如有渗出溶液,将其浇回土壤中,处理CK浇等量的去离子水。2018年10月分别测定各处理株高、地径,然后收获全部植株,并按吸收根、运输根、主干、枝条和叶5个器官分别进行相关生物量等指标的测定,其中分析测定的叶样均为成熟的功能叶;植株根系按功能划分法[17]进行分类,其中1~3级为吸收根,4~5级为运输根。
1.4 测定指标及方法
1.4.1 生长指标 (1)株高、地径净生长量。每个处理随机取3株,分别采用直尺和电子游标卡尺测定Cd处理前后巨桉幼树的株高和地径,计算不同处理株高、地径净生长量,计算公式为:株高(地径)净生长量=Cd处理后株高(地径)-Cd处理前株高(地径)。
(2)生物量。试验结束后,将巨桉幼树按吸收根、运输根、主干、枝条、叶全株收获,用去离子水将各器官洗净并晾干后,于105 ℃杀青30 min,于65 ℃恒温烘干至质量恒定,称其生物量,计算总生物量:总生物量=吸收根干质量+运输根干质量+主干干质量+枝条干质量+叶干质量。
(3)根系活力。采用TTC法[18]测定根系活力。
1.4.2 各器官营养元素含量 将各器官(吸收根、运输根、主干、枝条、叶)置于烘箱中105 ℃杀青30 min后,再经65 ℃烘干至质量恒定,粉碎并过孔径1 mm筛,测定N、P、K含量,其中N含量使用全自动间断化学分析仪(AMS,Italy)测定,P含量使用紫外吸收分光光度计(Shimazu,Japan)测定,K含量使用原子吸收分光光度计(Shimazu,Japan)测定。
1.4.3 Cd含量 将各器官(吸收根、运输根、主干、枝条、叶)于105 ℃杀青30 min后,再经65 ℃烘干至质量恒定,粉碎研磨并过孔径1 mm筛,采用干法灰化法消解制样;将土壤样品在室内自然风干并过孔径1 mm筛后,用HNO3-HClO4(二者体积比为9∶1)消解制样。分别采用石墨炉法和火焰法测定植株各器官和土壤中的Cd含量,测定仪器为原子吸收分光光度计。计算各器官中Cd的分配比例。
1.5 数据处理与统计分析
Cd富集系数(bioconcentration factor,BF)和转运系数(translocation factor,TF)计算的方法如下[19]:富集系数=植物地上部分(主干、枝条和叶)Cd含量/土壤中Cd含量,转运系数=植物地上部分(主干、枝条和叶)Cd含量/植物地下部分(吸收根、运输根)Cd含量。
所有数据均使用Excel 2016(Microsoft Office,USA)软件进行整理,采用SPSS 22.0(IBM Inc,USA)进行单因素方差分析(One-way ANOVA),显著水平α设定为0.05,用Duncan’s法进行多重比较,最后用Origin 2018(Origin,USA)制图。
2 结果与分析
2.1 Cd胁迫对巨桉幼树生长的影响
由图1和表1可知,随着Cd含量的增加,巨桉幼树株高和地径的净生长量均呈下降趋势,其中除了T1处理,T2、T3和T4处理株高和地径的净生长量与CK间差异均达显著水平;总生物量呈先下降后上升的趋势,且各处理间无显著差异;地下部生物量呈上升的趋势,其中T2、T3和T4处理与CK间差异达显著水平;地上部分生物量呈先下降后上升趋势,其中T2、T3和T4处理均显著低于CK;根系活力变化趋势与地上部分生物量相似,也呈下降的趋势,其中T3和T4处理与CK差异达显著水平。
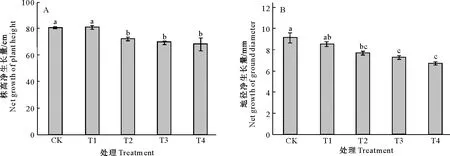
图柱上标不同小写字母表示不同处理间差异显著(P<0.05)

表1 Cd胁迫对巨桉幼树生物量及根系活力的影响
2.2 Cd胁迫对巨桉幼树各器官中营养元素含量的影响
表2显示,随着Cd含量的增加,巨桉幼树各器官中N含量总体均呈降低趋势。除了T4处理吸收根、主干和叶中的N含量显著低于CK外,其他处理各器官中N含量与CK均无显著差异。
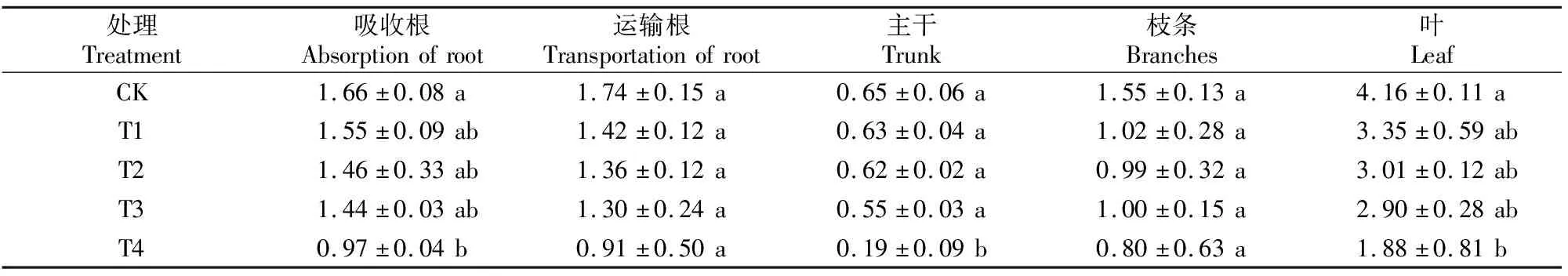
表2 Cd胁迫对巨桉幼树体内N含量的影响
表3显示,随着Cd含量的增加,各器官中P含量总体均呈降低趋势。除了T2、T3、T4处理吸收根和T4处理主干 P含量显著低于CK外,其他处理各器官中P含量与CK均无显著差异。
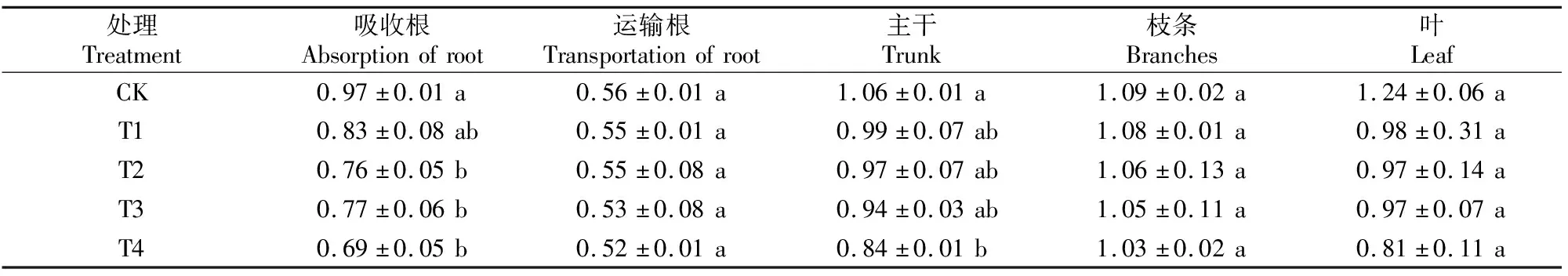
表3 Cd胁迫对巨桉幼树体内P含量的影响
表4显示,随着Cd含量的增加,各器官中K含量总体均呈降低趋势。T1~T4处理吸收根和主干中的K含量较CK均显著降低;T4处理枝条和叶中K含量显著低于CK,其他处理与CK间差异不显著;T1~T4处理运输根中的K含量与CK之间均无显著差异。结果表明,Cd胁迫下巨桉幼树对N、P、K元素的吸收均受到一定程度的抑制。

表4 Cd胁迫对巨桉幼树体内K含量的影响
2.3 Cd胁迫对巨桉幼树各器官Cd富集与转运的影响
Cd胁迫对巨桉幼树各器官Cd含量的影响如图2所示。图2显示,随着Cd含量的增加,巨桉幼树各器官中Cd含量均呈增加趋势。在相同Cd处理水平下,各器官中Cd含量由大到小的顺序依次为吸收根>运输根>主干>枝条>叶,其中地下部分器官(吸收根、运输根)中Cd含量远高于地上部分器官(主干、枝条、叶)。
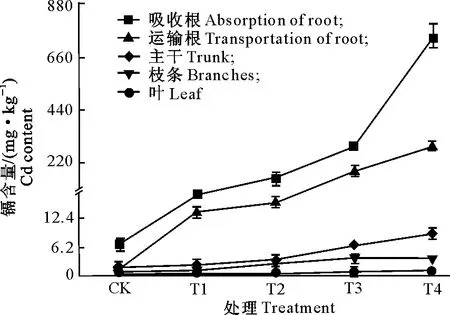
图2 Cd胁迫对巨桉幼树各器官Cd含量的影响
不同Cd处理下植株各器官中的Cd含量占全株的比例见表5。表5显示,各处理吸收根Cd分配比例最高,其余器官依次为运输根、主干、枝条、叶,其中吸收根Cd分配比例为65.59%~77.67%;运输根Cd分配比例为10.09%~26.93%;主干Cd分配比例不超过10%,且随着Cd含量的增加显著降低;枝条和叶中Cd含量所占比例较低,且随Cd含量的增加总体呈下降趋势。CK各器官分配比例显示,主干Cd含量在全株中所占比例最高,超过50%。
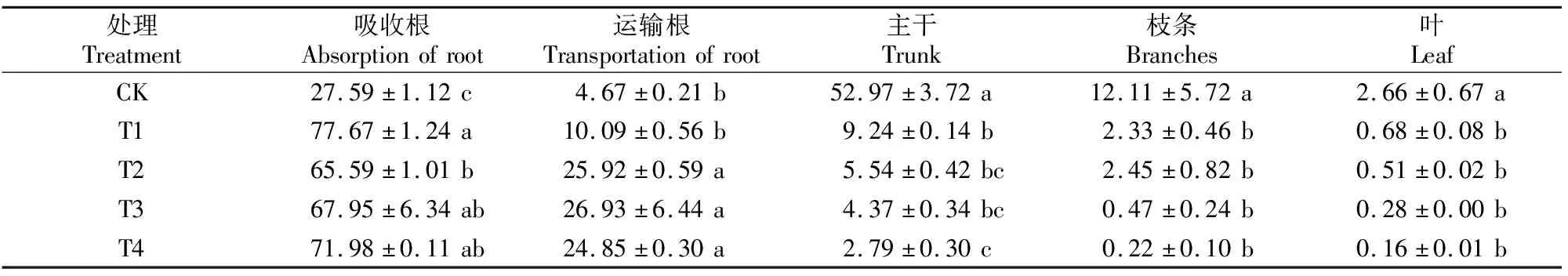
表5 Cd胁迫对巨桉幼树各器官Cd分配比例的影响
不同Cd处理下巨桉幼树各器官的Cd富集系数和转运系数见表6。表6显示,随着Cd含量的增加,吸收根的Cd富集系数呈增加趋势;运输根的Cd富集系数呈先增后降的趋势;主干和叶的Cd富集系数均在T1处理时下降,T2处理时上升,之后再次下降;枝条的Cd富集系数呈先上升后降低趋势,各处理间无明显变化。随着Cd含量的增加,各处理的Cd转运系数呈降低趋势。结果表明,巨桉幼树的地下部分对Cd有较强的吸收能力,但其转运能力偏低。
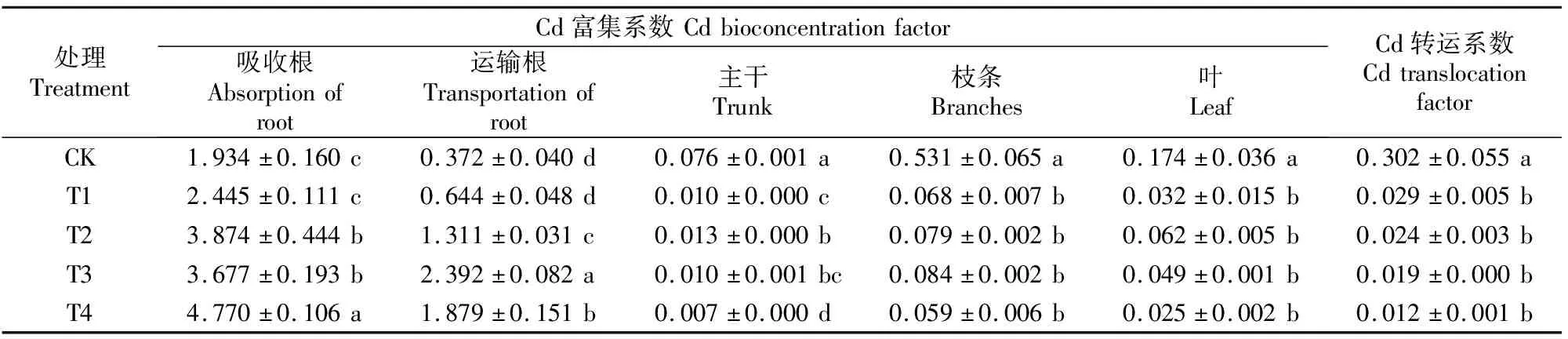
表6 不同Cd处理下巨桉幼树各器官的Cd富集系数和转运系数
3 讨 论
3.1 Cd胁迫对巨桉幼树生长的影响
植物对长期持续的Cd胁迫最突出的响应表现为叶片失绿、生长缓慢、生物量下降等[3]。本试验中,随着Cd胁迫的加剧,巨桉幼树株高和地径的净生长量总体均呈降低趋势,表明在较高含量Cd胁迫下巨桉幼树受到一定程度的毒害作用,这与Lunácková等[20]和Scebba等[21]的研究结果相似。此外,植物在遭受Cd胁迫时,其根系是最先接触Cd的器官,因此根系活力是判断植物根系抗御逆境能力的重要生理指标[22]。本研究结果表明,随着Cd含量的不断升高,巨桉幼树根系活力受到了明显抑制,说明Cd胁迫对巨桉幼树根系产生了毒害作用。本研究中,随着Cd胁迫的加重,巨桉幼树生物量的分配格局也发生明显改变,其中地上部生物量减少,这一方面可能是由于Cd胁迫导致植株的光合作用、膜系统和酶系统等与新陈代谢相关的多种生理活动受抑所致,另一方面,Cd可能还干扰了植株对矿质元素及水分的吸收、同化及分配过程[22],从而抑制其地上部分的生长[23-24];而地下部分生物量增加,这可能是其根系抗氧化酶的协同作用对植株的保护作用比根系活力下降的影响更显著,从而保证了根系的正常生长[25-26]。地下部分生物量增加以及地上部分生物量减小,二者两两相抵,最终导致总体上生物量变化不显著,表明巨桉幼树对Cd的胁迫有一定的耐受能力。
3.2 Cd胁迫对巨桉幼树体内养分特征的影响
N、P元素的缺乏会导致植株矮小、叶片变黄、长势减弱、叶边缘卷曲、叶片脱落加剧;K元素的缺乏会导致植株对CO2的吸收受阻,影响光合作用[27]。本研究结果发现,随着Cd含量的增加,巨桉幼树各器官N、P、K含量总体均呈降低趋势,这与谭长强等[28]的研究结果相似,可能是植物根系中的细胞质膜不存在Cd的特殊转运蛋白通道[29],Cd进入植物体内,占据其他营养元素离子通道,抑制了巨桉幼树根系对营养元素的吸收;也可能是Cd胁迫引起的膜脂过氧化增加了细胞膜透性,导致部分小分子营养物质外流[27],降低了植株对N、P、K元素的吸收效率。
3.3 巨桉幼树对Cd的富集、转移特性
植物在土壤中对Cd2+的吸收及其在不同器官中的分布因不同物种而异。本研究结果表明,在同一Cd处理水平下,吸收根Cd含量最高,其余依次为运输根、主干、枝条、叶,这与贾中民等[30-31]的研究结果一致,说明植物将大量Cd集中在根系,其原因主要有3方面:(1)根系细胞壁中纤维素、木质素中的羧基、羟基、醛基等活泼基团与Cd2+结合形成沉淀,积累于果胶位点上,阻止较多的Cd2+进入细胞质内[32-33],造成大量Cd2+集中在根系细胞壁中。(2)根系细胞质中的氨基酸、有机酸、多肽或无机物等与细胞质中游离的Cd2+结合,通过液泡膜上的运载体或通道蛋白进入液泡中,将Cd2+限制在液泡中[34],从而抑制Cd2+向地上部分运输。(3)植物可以通过植物螯合肽、金属硫蛋白、有机酸、氨基酸等螯合剂与Cd2+产生螯合作用[35],将Cd2+滞留在根系中,最终导致大量的Cd集中在根部,限制其向地上部分转移,从而减少Cd积累对植株的伤害,增强其对Cd的耐受性[36-37]。
定义Cd超富集植物有两个基本标准,第一是植物叶片或地上部中含Cd达到100 mg/kg(以干质量计),第二是转运系数大于1[38]。本研究中,巨桉幼树地上部Cd含量远低于100 mg/kg,转运系数均小于1,表明巨桉幼树对Cd的富集转运能力有限。
植物各器官生理功能和代谢活动不同,其细根、叶、花、果实等器官常会因死亡、凋落并经分解反回土壤,构成Cd二次污染,而茎(主干+枝条)固定的Cd才具有一定的稳定性,有利于收获后进一步的处理。巨桉幼树茎中Cd含量随Cd胁迫程度的加剧呈上升趋势,并且随着树龄的增大,其茎的生物量会进一步增加,Cd的累积量也将逐渐增加。
综上可知,巨桉幼树虽然不是Cd的超富集植物,对Cd的转运能力有限,但其生长迅速,即使在高含量Cd的胁迫下其生长并未受到明显抑制,说明巨桉幼树对Cd有较强的适应性,因此可以初步确定巨桉为重金属Cd污染地区土壤修复树种。
