LncRNA 调控肿瘤细胞辐射敏感性作用的研究进展
2021-03-18赵舒雅李航樊赛军
赵舒雅 李航 樊赛军
中国医学科学院北京协和医学院放射医学研究所,天津市放射医学与分子核医学重点实验室 300192
长链非编码RNA(long non-coding RNA, lncRNA)是一组转录本长度大于200 个核苷酸的RNA 分子,存在于细胞核内或胞浆内,能够通过多种机制影响肿瘤细胞的信号通路,并在染色质水平、转录水平和转录后水平影响靶基因的表达[1]。LncRNA具有特异性细胞类型表达、定位于特定亚细胞区室、与人类疾病相关联等特点[2]。在细胞内,多数lncRNA 定位于细胞核,与核质蛋白结合,以促进核保留,并以顺式(in cis)或反式(in trans)方式结合在其调控基因附近,通过募集转录因子、组蛋白修饰因子或染色质修饰因子调控其靶基因的表达。胞质lncRNA 在调控基因表达的过程中最常作为竞争性内源性RNA(ceRNA),竞争性内源性RNA 通过与微小RNA(m icroRNA,m iRNA)反应原件(MRE)竞争相同的m iRNA 调节靶基因转录本的表达[3]。此外,在空间分布上,肿瘤细胞与正常细胞、辐射或药物敏感细胞与辐射抵抗细胞中lncRNA 的表达量也存在差异[4-5]。
放疗是多种恶性肿瘤的有效治疗方法之一,约半数的恶性肿瘤患者在治疗过程中需要使用放疗。然而,肿瘤细胞的辐射抵抗是导致放疗失败的主要原因[6],因此,提高肿瘤细胞的放疗敏感性是治疗成功的关键。电离辐射可直接作用于细胞核DNA,导致DNA 双螺旋结构断裂,破坏遗传物质;同时,电离辐射能够诱导细胞内的水分子发生电离,产生大量的自由基分子,其与蛋白质等有机大分子发生反应,可直接破坏细胞结构,产生不可逆的损伤,最终导致细胞死亡[7]。
1 LncRNA 的作用模式
LncRNA 能够通过多种方式调控基因的表达。LncRNA 能够通过与DNA、RNA 和蛋白质的相互作用调节染色质的结构和功能以及邻近和远处基因的转录,并影响RNA 的剪接、稳定性和翻译过程,部分lncRNA 还参与细胞器的形成和功能调节。
有研究者通过全基因组RNA-染色质关联分析,结合染色质构象捕获技术,揭示了部分lncRNA调节染色质结构和基因表达的过程[8]。从机制上看,顺式作用和反式作用的核lncRNA 都能与DNA相互作用,改变染色质的结构,部分情况下其通过调控转录因子间接作用[8],其他情况下以特异序列直接结合DNA, 介导基因沉默或激活。
部分lncRNA 通过协同作用、转录依赖和非转录依赖机制调控蛋白编码基因的表达,并能够通过剂量补偿效应介导基因沉默。增强子相关lncRNA(elncRNA)的剪接与其相关增强子的活性和邻近蛋白编码基因的丰度相关[9],并且可以与染色质调节蛋白共同调节染色质结构。
除了在转录调控和调节染色质结构中的作用外,lncRNA 还控制着基因表达的其他几个方面。部分lncRNA 能够被翻译成具有功能的肽段,部分lncRNA 能够通过RNA 基序或结构域结合蛋白质,形成特定的lncRNA-蛋白质复合物(lncRNP),参与转录后调控,改变mRNA 的剪接过程,进而实现对信号通路的调节。一些lncRNA 可以直接与mRNA 进行碱基配对并招募mRNA 降解相关蛋白质。此外,大量lncRNA 携带m iRNA 的互补位点,可以作为竞争性的内源性RNA 或m iRNA 的“海绵”来调节基因的表达。
部分lncRNA 定位于外泌体和线粒体等特定细胞器。由于外泌体会定期释放到细胞外环境中,因此定位于外泌体的lncRNA 可以最终进入受体细胞,参与表观遗传的调控、细胞类型的重编程等过程,定位于线粒体的lncRNA 可以由核DNA 和线粒体DNA 编码,并且通常与线粒体的代谢和细胞凋亡等过程关联[10]。
2 LncRNA 介导的肿瘤辐射抗性
通过高通量转录组学分析发现,肿瘤患者组和健康对照组中lncRNA 的表达量存在显著差异[11]。LncRNA 能够通过多种分子机制影响细胞的放射敏感性(图1),调节DNA损伤修复、逆转细胞周期阻滞、细胞凋亡、上皮-间充质转化(epithelial-tomesenchymal transition,EMT)和自噬等[12]过程,在肿瘤的治疗和预后过程中具有潜在价值。根据部分lncRNA 在不同类型肿瘤放射敏感性调节等方面的作用对其进行归纳(表1)。
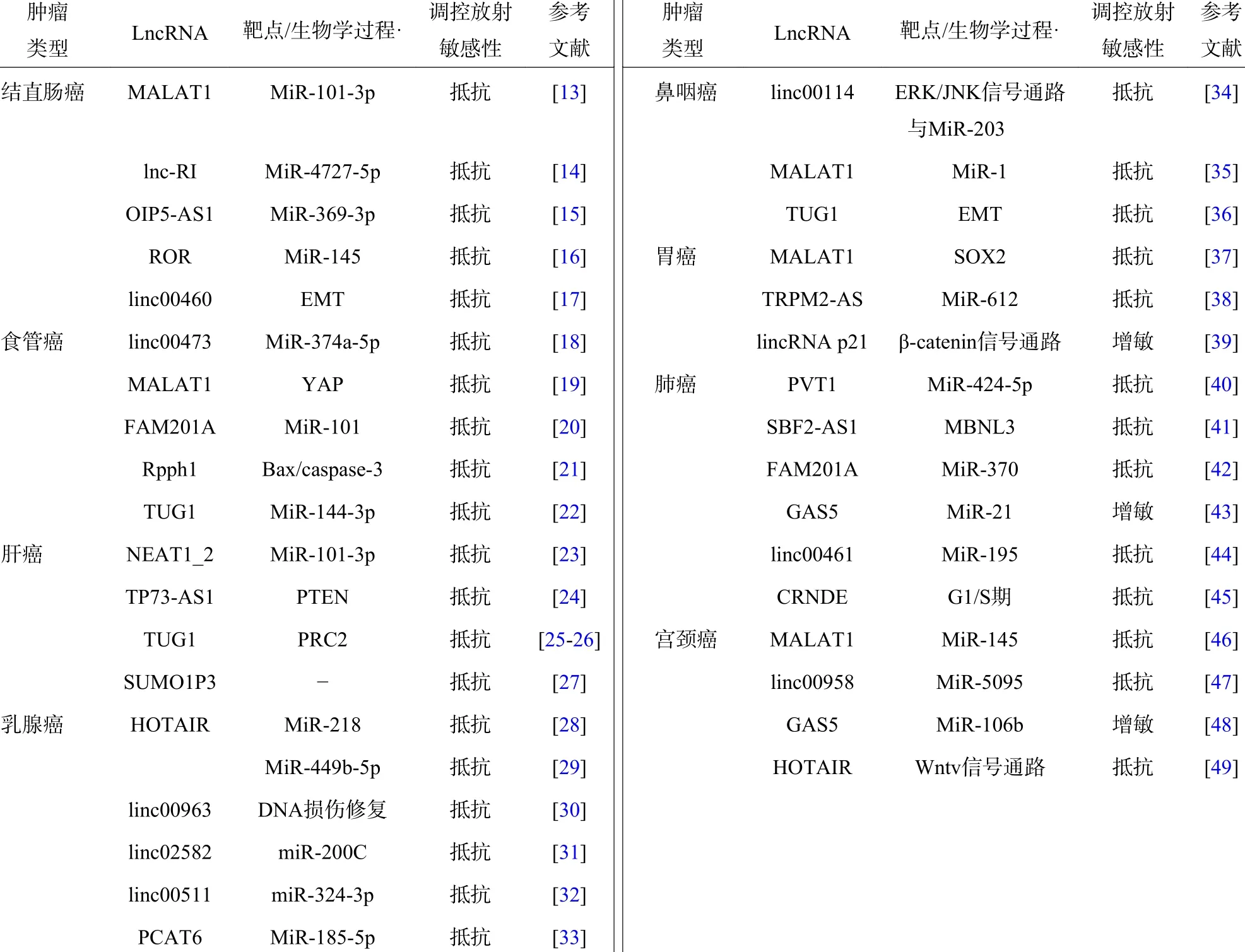
表1 不同类型肿瘤中对放射敏感性具有调节作用的lncRNATab le 1 LncRNA with regulatory effects on radiosensitivity in different tumor types
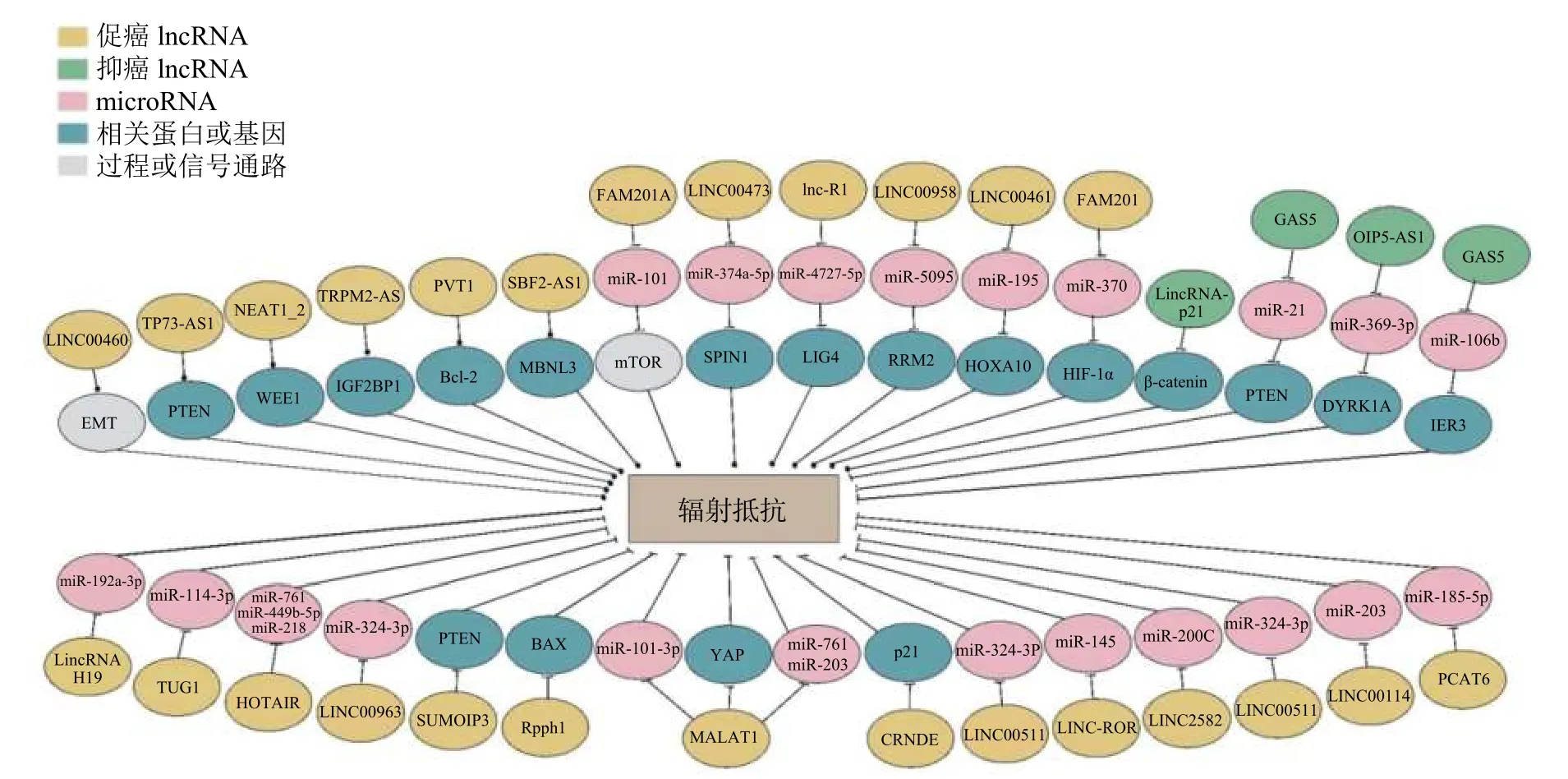
图1 LncRNA 介导肿瘤细胞辐射抵抗的多种机制 lncRNA 为长链非编码RNA;m icroRNA 为微RNA;EMT 为上皮-间充质转化;TP73-AS1 为p53 依赖性凋亡调节物;PTEN 为人第10 号染色体缺失的磷酸酶及张力蛋白同源的基因;NEAT1_2 为核富集转录本1;WEE1 为G2 检查点激酶;TRPM 2 -AS 为瞬时受体电位M 2 反义RNA ;IGF2BP1 为胰岛素样生长因子2 mRNA 结合蛋白1;PVT1 为浆细胞瘤变异易位1;Bcl-2 为B 淋巴细胞瘤2 基因;SBF2-AS1 为SBF2 反义 RNA1;MBNL3 为Muscleblind 样蛋白3;FAM 201A为序列相似性家族201-成员A;m TOR 为雷帕霉素靶蛋白;SPIN1 为甲基赖氨酸读写蛋白spindlin 1;LIG4 为DNA 连接酶Ⅳ;RRM 2 为人核糖核苷酸还原酶M 2;HOXA10 为人类同源异型盒基因A10;HIF-1α 为缺氧诱导因子1α;GAS5 为生长阻滞特异性转录物5;OIP5-AS1 为Opa 相互作用蛋白5 反义转录本1;DYRK1A 为双底物特异性酪氨酸磷酸化调节激酶1A;IER3 为人早期应答基因3;TUG1 为牛磺酸上调基因1;HOTAIR 为人类同源异型盒基因转录反义RNA;SUMO1P3 为小泛素样修饰蛋白1 假基因3;BAX 为Bcl-2 相关X 蛋白质;Rpph1 为核糖核酸酶P RNA 组份H1;YAP 为Yes 相关蛋白;MALAT1 为肺腺癌转录本1;CRNDE为结直肠肿瘤差异表达基因;ROR 为重编程调控因子;PCAT6 为前列腺相关转录本6Figure 1 LncRNA mediates tumor cell radioresistance through multiple mechanisms
2.1 结直肠癌
结直肠癌居全球常见恶性肿瘤第3 位,其高发病率和高病死率严重影响着人类健康。由于临床上结直肠癌对放疗的敏感性较低,因此其治疗效果受到很大影响。LncRNA 的异常表达与结直肠癌的发生、发展、侵袭和转移等过程密切相关。有研究结果表明,部分lncRNA 能够通过海绵化miRNA 实现对放射敏感性的调控,lncRNA 肺腺癌转录本1(metastasis-associated lung adenocarcinoma transcript 1,MALAT1)[13]、长链非编码RNA RI(lncRNA radiation induced,lnc-RI)[14]、Opa 相互作用蛋白5反义转录本1(OIP5-AS1)[15]和lncRNA 重编程调控因子(regulator of reprogramm ing,ROR)[16]可以通过竞争性结合m iR-101-3p、m iR-4727-5p、miR-369-3p 和m iR-145 调节结直肠癌的辐射抗性。DNA 双链断裂的修复效率是影响肿瘤放射敏感性的重要因素之一,lnc-RI 能够通过lnc-RI/m iR-4727-5p/DNA连接酶Ⅳ(LIG4)轴调节DNA 连接酶Ⅳ的表达,提高非同源末端连接的修复效率,促进DNA 双链断裂的修复,降低肿瘤细胞的辐射敏感性[14]。与lnc-RI 参与的耐辐射机制不同的是,lncRNA Opa 相互作用蛋白5 反义转录本1(OIP5-AS1)能够抑制m iR-369-3p 的表达,从而上调m iR-369-3p 下游基因双底物特异性酪氨酸磷酸化调节激酶1A(DYRK1A)的表达,最终抑制细胞克隆存活,促进细胞凋亡,提高结直肠癌细胞的放射敏感性[15],且有研究结果发现,Opa 相互作用蛋白5 反义转录本1(OIP5-AS1)在抗辐射肿瘤组织中高表达。LncRNA ROR 则通过负调控p53/m iR-145 降低结直肠癌细胞系的放射敏感性[16]。此外,linc00460 在人结直肠腺癌(HCT)116 细胞中的表达明显高于其他类型的细胞,辐射能够激活位于linc00460 启动子上的c-Jun 转录因子结合元件,从而诱导linc00460 的表达,促进细胞EMT 并产生辐射抗性[17]。
2.2 食管癌
食管癌是常见的消化道肿瘤,居全球最常见的恶性肿瘤第7 位,病死率居第6 位。食管癌的病理类型主要有鳞癌和腺癌,我国以鳞癌为主。Linc00473 在几种人类恶性肿瘤中异常表达,Chen等[18]发现linc00473 和m iR-374a-5p 之间存在相互抑制的作用,甲基赖氨酸读写蛋白spindlin1(spindlin1,SPIN1)是m iR-374a-5p 的下游靶点,linc00473 可以通过负调控m iR-374a-5p 的表达上调SPIN1 的表达,从而增强食管鳞状癌(ESCC)细胞的辐射抗性。此外,lncRNA MALAT1 在多种恶性肿瘤中具有促进肿瘤发生发展的作用,Yao 等[19]发现敲除MALAT1 能够降低食管鳞状癌细胞的干性与迁移能力,增强细胞的放射敏感性,同时MALAT1 还可直接与Yes 相关蛋白(Yes related protein 1,YAP)结合,促进YAP 蛋白的表达并提高YAP 的转录活性,从而逆转MALAT1 敲除对食管鳞状癌细胞的干性和放射敏感性的影响。LncRNA序列相似性家族201-成员A(fam ily with sequence sim ilarity 201-member A,FAM 201A)在辐射抵抗的食管鳞状癌(ESCC)肿瘤组织中表达上调,FAM 201A敲除可以通过负调控m iR-101 的表达水平,调控雷帕霉素靶蛋白(m TOR)信号通路,从而增强人食管癌细胞ECA109 和(或)ECA109R 的放射敏感性[20]。LncRNA 核糖核酸酶P RNA 组份H1(ribonuclease P RNA component H1,Rpph1)在食管癌中高表达,降低Rpph1 的表达水平可增加凋亡相关蛋白Bax 和caspase-3 的表达,同时B 淋巴细胞瘤2 基因(Bcl-2)表达水平降低。此外,干扰Rpph1 能够减轻辐射诱导的G2/M 期阻滞,进而显著抑制食管癌细胞中与细胞增殖、迁移和EMP 调控相关蛋白的表达[21]。LncRNA 牛磺酸上调基因1(taurine upregulation gene 1,TUG1)在食管鳞状癌细胞和组织中的表达上调,抑制LncRNA TUG1 能够抑制食管癌细胞的增殖和集落的形成,同时诱导细胞凋亡,体内和体外实验均已证明敲除lncRNA TUG1可以上调m iR-144-3p 的表达水平,通过间质-表皮转化因子(cellular-mesenchymal epithelial transition factor,c-Met))/表皮生长因子受体(epithelial grow th factor receptor,EGFR)/丝氨酸-苏氨酸激酶(Akt)轴提高食管鳞癌的辐射抗性[22]。
2.3 肝癌
肝癌是人类第二大致死性恶性肿瘤,lncRNA的异常表达被认为与肝癌辐射抵抗的发生和治疗预后密切相关。其中,lncRNA 核富集转录本1(NEAT1_2)[23]能够通过竞争性结合m iR-101-3p调节肝癌的放射抗性。下调lncRNA 核富集转录本1(NEAT1_2)可以通过调节m iR-101-3p/丝氨酸/苏氨酸G2 检查点激酶 1(WEE1)轴使肝癌细胞的放射敏感性增强。已有研究结果证实,lncRNA p53 依赖性凋亡调节物(tumor protein P73 antisense RNA 1,TP73-AS1)在肝癌中高表达,并通过人第10 号染色体缺失的磷酸酶及张力蛋白同源的基因(phosphate and tension homology deleted on chromsome ten,PTEN)/丝氨酸-苏氨酸激酶(Akt)信号通路参与肝癌的放射抵抗[24]。核转录因子特异性蛋白1(specificity protein,SP1)能够诱导lncRNA TUG1 过表达,TUG1 可与多梳蛋白抑制复合体 2(PRC2)结合并招募至Krüppel 样因子2(KLF2) 的启动子区域,从而抑制Krüppel 样因子2 的转录,TUG1 可作为肝癌的生物靶标,增强其放疗敏感性[25-26]。此外,lncRNA 小泛素样修饰蛋白1 假基因3(SUMO1P3)的敲除可以显著抑制肝癌的生长、侵袭、促进细胞凋亡并增强细胞的放射敏感性,为小泛素样修饰蛋白1 假基因3(SUMO1P3)可能成为一种潜在的生物标志物和肝癌的治疗靶点提供了可靠证据[27]。
2.4 乳腺癌
近年来,乳腺癌的发病率呈逐年上升的趋势,其发病率排在女性恶性肿瘤的首位。术前和术后放疗在乳腺癌的治疗中起着举足轻重的作用,但经常因肿瘤细胞产生辐射抵抗,最终导致患者预后不良。LncRNA 人类同源异型盒基因转录反义RNA(homeotic genes transcript antisense RNA,HOTAIR)在乳腺癌细胞和组织中表达上调,最近的研究结果表明,HOTAIR 的表达随照射时间的延长而增加,且HOTAIR 基因的敲除抑制了细胞的存活,增强了细胞对电离辐射的敏感性,这一放射增敏作用与HOTAIR 的竞争性内源RNA m iR-218 上调有关[28]。此外,Zhang 等[29]的研究结果证实,lncRNA HOTAIR 可与m iR-449b-5p 竞争性结合,诱导休克蛋白A1A(HSPA1A)的表达,导致乳腺癌细胞的辐射抗性显著增强。有研究结果证实,沉默linc00963可以抑制乳腺癌细胞的增殖和成瘤,敲除linc00963会促进DNA 损伤和氧化应激,并使乳腺癌细胞的放射敏感性增强[30]。Linc02582 能够将m iR-200C与其靶蛋白细胞周期检测点激酶1(Chk1)连接到一起,通过miR-200C 下调检测点激酶1 的表达水平,从而提高乳腺癌细胞的辐射敏感性[31]。此外,有研究结果表明,linc00511 在乳腺癌组织中的表达水平显著升高,并与保乳手术后行放疗患者的复发和生存不良密切相关。体外小干扰RNA 敲除实验结果证实,linc00511 能够抑制细胞增殖,诱导凋亡加快,最终增强细胞的放射敏感性;同时,敲除linc00511 在体内实验中表现为肿瘤的生长受到抑制,辐射的敏感性增强[32]。最近的研究结果表明,lncRNA 前列腺相关转录本6(prostate cancer associated transcript 6,PCAT6)在三阴性乳腺癌(TNBC)的组织和细胞中高表达,PCAT6 可以通过与m iR-185-5p 的竞争性结合调节肿瘤蛋白D52(tumor protein D52,TPD52)的表达,从而调节三阴性乳腺癌细胞的放射敏感性,抑制PCAT6 能够通过调节m iR-185-5p/肿瘤蛋白D52 轴增强三阴性乳腺癌细胞的放射敏感性[33]。
2.5 鼻咽癌
鼻咽癌是一种起源于鼻咽上皮的恶性肿瘤,易发生远处转移和放射抵抗,lncRNA 的异常表达与鼻咽癌的发生发展、侵袭转移和治疗敏感性等方面密切相关。LncRNA linc00114 在鼻咽癌患者的血清、鼻咽癌组织和鼻咽癌细胞系中表达上调,且通过调节细胞外调节蛋白激酶(ERK)/ c-Jun 氨基末端激酶(JNK)信号通路和m iR-203 诱导鼻咽癌的发生发展和辐射抵抗,这提示linc00114 是一种很有前景的与鼻咽癌发生发展和辐射抵抗相关的生物标志物[34]。此外,lncRNA MALAT1 在鼻咽癌细胞系和组织中显著上调,体内外实验结果均证实,敲除MALAT1 基因可提高鼻咽癌细胞的辐射敏感性,其分子机制为MALAT1 能够通过调节肿瘤干细胞的活性介导辐射抵抗,因此,MALAT1 有望成为鼻咽癌患者治疗的靶点[35]。LncRNA TUG1 在鼻咽癌组织和鼻咽癌细胞系中的表达水平高于正常鼻咽上皮组织和正常鼻咽细胞系,TUG1 水平较高的鼻咽癌患者的总存活率较低,TUG1 下调可以抑制EMT,降低鼻咽癌细胞的增殖、迁移和侵袭能力,而过表达TUG1 则显著促进鼻咽癌细胞的增殖、迁移和侵袭[36]。
2.6 胃癌
胃癌是全球最常见的恶性肿瘤之一,是恶性肿瘤第三大病死原因。近些年的研究结果证实,lncRNA 在胃癌的诊断和预后中或可扮演重要角色。既往研究结果证实,lncRNA MALAT1 能够促进结直肠癌的EMT 和血管生成,过表达MALAT1能增强胃癌悬浮细胞的干性,MALAT1 敲降后,胃癌悬浮细胞的放射敏感性和化疗敏感性均增强[37]。其机制为通过MALAT1 直接与Y 染色体性别决定区相关的高迁移率族盒蛋白2(Sox2)结合,增强Y 染色体性别决定区相关的高迁移率族盒蛋白2(Sox2)的稳定性并促进其表达,Y 染色体性别决定区相关的高迁移率族盒蛋白2(Sox2)敲降可以部分逆转MALAT1 过表达对胃癌细胞干性的促进作用。也有研究通过筛选胃癌的肿瘤基因组(TCGA)数据库,确定了在胃癌组织中异常表达的关键lncRNA瞬时受体电位M 2 反义RNA(TRPM 2-AS)[38],其可作为miRNA 的“海绵”或竞争性内源RNA 抑制肿瘤与miR-612 结合,从而调节胰岛素样生长因子2 mRNA 结合蛋白1(IGF2BP1)和叉头框蛋白M 1(FOXM 1)的表达,胰岛素样生长因子2 mRNA 结合蛋白1 上调后提高c-Myc 的表达,最终促进癌细胞的发生发展。此外,lincRNA p21 也能够通过靶向β-catenin 信号通路提高胃癌细胞的放疗敏感性[39]。
2.7 肺癌
LncRNA 参与了包括非小细胞肺癌(non-small cell lung cancer,NSCLC)在内的多种肿瘤的发展。Wang 和Hu[40]研究发现,浆细胞瘤变异易位 1(plasmacytoma variant translocation 1,PVT1)是NSCLC 中上调最显著的lncRNA,NSCLC 组织中的PVT1、共激活因子相关的精氨酸甲基转移酶 1(coactivator associated arginine methyltransferase 1,CARM 1)、基质金属蛋白酶(matrix metalloprotein,MMP )-2、MMP-9 和B 淋巴细胞瘤2 基因表达量升高,miR-424-5p 和Bax 表达量降低。PVT1 敲降或过表达miR-424-5p 后,PVT1、CARM 1、MMP-2、MMP-9 和B 淋巴细胞瘤2 基因下调,细胞增殖、迁移和浸润受阻;m iR-424-5p、Bax 上调,细胞凋亡增加,NSCLC 细胞的辐射敏感性增强。Yu 等[41]的研究结果表明,lncRNA SBF2 反义RNA1(SBF2-AS1)敲降后,Muscleblind 样蛋白3(MBNL3)下调,NSCLC 细胞的辐射敏感性增强,细胞凋亡显著增加。LncRNA FAM 201A 能够通过竞争性靶向miR-370 提高EGFR 和缺氧诱导因子1α(hypoxia inducible factor-1α,HIF-1α)的水平;FAM 201A 敲降后,EGFR 和HIF-1α 的水平降低,细胞的放射敏感性增强[42]。电离辐射能有效促进lncRNA 生长阻滞特异性转录物5(grow th arrest-specific transcript 5,GAS5)与m iR-21/PTEN/Akt 轴之间的相互作用,lncRNA GAS5 对NSCLC 具有放射增敏作用[43]。有研究结果表明,linc00461 和人类同源异型盒基因A10(HOXA10)在原发性和转移性肺腺癌中高表达;同时linc00461 可与m iR-195 结合,通过人类同源异型盒基因A10 促进肺腺癌细胞的迁移和侵袭,并降低癌细胞的放射敏感性,为肺腺癌的靶向治疗提供了新的依据[44]。LncRNA 结直肠肿瘤差异表达基因(colorectal neoplasia differentially expressed,CRNDE)在肺腺癌(lung adenocarcinoma,LAD)组织和辐射抵抗的LAD 细胞系中显著上调,CRNDE的高表达与NSCLC 的低分化、TNM 分期、淋巴结转移、放疗疗效和总生存期显著缩短均密切相关。功能获得和功能丧失实验结果表明,CRNDE 敲降可以通过影响体外培养的LAD 细胞的G1/S 期转换和诱导细胞凋亡影响LAD 细胞的放射敏感性[45]。
2.8 宫颈癌
宫颈癌是最常见的妇科恶性肿瘤之一,是一种世界范围内病死率较高的妇科疾病,目前包括放化疗在内的针对宫颈癌的治疗有效率均偏低。MALAT1 是一种在包括宫颈癌在内的多种肿瘤中发挥致癌作用的lncRNA,MALAT1 在放射抵抗的肿瘤组织中的表达水平明显高于放射敏感的肿瘤组织。有研究结果证实,在高危型人乳头状瘤病毒(HR-HPV)阳性患者的宫颈癌细胞中,MALAT1和m iR-145 的表达呈负相关,敲除MALAT1 基因的人宫颈癌肠转移CaSki 和人宫颈癌Hela 细胞受到辐射后的集落形成率明显降低,G2/M 期阻滞比例明显升高,细胞凋亡率明显升高,辐射敏感性提高[46]。放疗联合linc00958 沉默、下调人核糖核苷酸还原酶M 2(RRM 2)或过表达m iR-5095 在体内外均能抑制宫颈癌细胞的增殖和肿瘤生长,同时促进细胞凋亡。Linc00958 可通过海绵化m iR-5095 提高人核糖核苷酸还原酶M 2 水平,从而调节宫颈癌细胞对放疗的敏感性[47]。LncRNA GAS5 过表达可通过m iR-106b 上调人早期应答基因3(IER3),增强宫颈癌细胞对放疗的敏感性[48]。放疗可降低肿瘤组织、Hela 细胞和C33A 细胞中HOTAIR 和HIF-1α的表达,而HOTAIR 过表达可阻断辐射对宫颈癌Hela 和宫颈癌C33A 的细胞活力和细胞凋亡的影响[49]。因此,阐明宫颈癌中lncRNA 介导肿瘤细胞放射敏感性的分子机制,或可为今后宫颈癌的治疗提供潜在的诊断和治疗靶点[50]。
3 小结
放疗是一种重要的肿瘤治疗方法,但其有效性受肿瘤细胞辐射抗性的影响。目前,肿瘤耐辐射的潜在机制尚未完全阐明,降低肿瘤细胞的辐射抗性、增强其辐射敏感性是恶性肿瘤研究领域的重要内容之一。
LncRNA 已经被证明可参与多种生物过程,如转录调控、蛋白修饰、翻译及RNA-蛋白和(或)蛋白-蛋白复合物的形成,通过靶向蛋白或m iRNA 调控细胞凋亡、EMT、DNA 损伤修复等多个过程,最终影响肿瘤对辐射的敏感性。就分子机制来说,lncRNA 可通过m iRNA、蛋白和相关信号通路介导肿瘤细胞的辐射抵抗,其中,m iRNA 参与多种lncRNA 介导的辐射抵抗过程。研究结果显示,miRNA 在肿瘤细胞的进展以及耐药性方面也发挥关键作用[51],且部分lncRNA 可调节多种肿瘤的放射敏感性,如,MALAT1 在结直肠癌、食管癌和鼻咽癌中都有促进辐射抵抗的作用,这为非编码RNA诱导的肿瘤的辐射抵抗和耐药性的治疗提供了一些新思路。此外,近年来有关lncRNA 在肿瘤中表达的研究进展提示了其作为生物标志物在患者诊断和预后中的潜在作用,但还没有研究揭示放疗疗效与lncRNA 作为放疗患者的生物标志物之间的关系,因此有必要对临床放疗敏感性与lncRNA 的相关性进行研究,进而直接证明lncRNA 与肿瘤细胞放疗敏感性的关系,也将为今后lncRNA 放疗相关靶向分子制剂的临床研究提供可能。同时,这一领域还需要进行更多的基础和临床研究对lncRNA 和信号分子如何协同作用进行探讨,从而调节肿瘤细胞的放射敏感性,研究和阐明lncRNA在放疗中的应用价值,为肿瘤治疗带来新的突破。
