红花1-脱氧-D-木酮糖-5-磷酸合成酶基因CtDXS1的克隆及表达分析
2021-03-15谭政委余永亮许兰杰杨红旗鲁丹丹马新明梁慧珍
谭政委,李 磊,余永亮,许兰杰,杨红旗,董 薇,鲁丹丹,马新明,梁慧珍
(1.河南省农业科学院 芝麻研究中心,河南 郑州 450002; 2.河南农业大学 信息与管理科学学院,河南 郑州 450002)
植物体能够产生丰富多样的次生代谢产物[1-2],这些代谢产物在植物生理生长过程中发挥着重要作用,可以增强植物对虫害、病害、UV-B辐射等生物及非生物胁迫的抵抗力,还有一些次生代谢产物,可以作为植物传粉者的引诱剂。植物次生代谢产物被开发成中药、农药、杀虫剂、染料等应用于多个领域。大量研究表明,高等植物的代谢多样性可能由植物为适应不同生态环境需要所驱动[3-4]。
萜类物质是植物中最大的一类次生代谢产物,目前,在植物中鉴定的萜类化合物超过4万种[5-6],包括单萜(如柠檬烯)、倍半萜(如橙花叔醇)、二萜(如紫衫醇)、三萜(如人参皂苷)、四萜(如类胡萝卜素)和多萜(如质体醌)[7]。萜类物质的合成前体异戊二烯由3-异戊烯基焦磷酸(Isopentenyl diphosphate,IPP)及其异构体二甲基丙烯焦磷酸(Dimethylallyl diphosphate,DMAPP)通过2条途径合成,分别是位于质体中的2-C-甲基-D-赤藻糖醇-4-磷酸(2-C-methyl-D-erythritol-4-phosphate,MEP)途径和位于胞质的甲羟戊酸(Mevalonate,MVA)途径[8-9]。1-脱氧-D-木酮糖-5-磷酸(1-deoxy-D-xylulose-5-phosphate,DXP)合成酶(1-deoxy-D-xylulose-5-phosphate synthase,DXS)是MEP途径中的第一个酶,催化丙酮酸和3-磷酸甘油醛生成DXP[10-12]。许多研究表明,DXS是MEP代谢通路的关键酶,也是IPP和DMAPP生物合成途径的限速酶[13]。
试验表明,在植物中过量表达DXS基因导致类胡萝卜素、叶绿素和其他萜类化合物的含量升高[14]。因为MEP途径在许多致病细菌和光合真核生物中是必不可少的,但在动物和人体中不存在该代谢途径,因此,该代谢途径的基因和酶是开发新型抗菌药物和除草剂的靶点[15-18]。研究表明,DXS可以作为靶向作用位点应用于番茄[14]和马铃薯块茎[19]异戊二烯类物质的调控,并且马铃薯StDXS1基因在抗马铃薯晚疫病中发挥一定作用[20],说明DXS基因可能在植物生长发育、抵抗生物及非生物胁迫中发挥一定的作用。
红花(CarthamustinctoriusL.),别名红蓝花、刺红花、草红花,是菊科红花属1年生双子叶草本植物,红花作为红花属中唯一的栽培种,是集油用、药用和工业用于一体的特种经济作物[21]。目前,从红花中分离到的代谢物超过100种,其中主要包括黄酮、生物碱、有机酸和炔烃类化合物,其中,从红花管状花分离到的炔烃类化合物有13种,这类化合物具有很好的抗抑郁活性[22],但到目前为止,关于红花萜类代谢生物合成相关基因的研究尚未见报道。河南省农业科学院芝麻研究中心药用植物研究室以豫红花1号为材料,构建了红花三代测序全长转录组数据库(数据尚未公开发表),以此为基础,通过RT-PCR方法克隆到红花DXS基因的cDNA全长,命名为CtDXS1,对其序列及进化关系进行分析,并对其非生物胁迫及激素处理响应进行相关研究,通过构建原核表达载体进行异源表达,诱导产生目的蛋白,为深入研究红花萜类化合物的生物合成机制提供参考。
1 材料和方法
1.1 材料
1.1.1 试验材料 红花品种豫红花1号及管状花颜色为白色(W)、黄色(Y)、橙色(O)、红色(R)的红花品系均来自河南省农业科学院芝麻研究中心,大肠杆菌菌株DH5α、大肠杆菌表达菌株Transetta(DE3)和表达载体pET28a为河南省农业科学院芝麻研究中心保存。
1.1.2 试剂 RNA提取试剂盒为华越洋生物公司的Quick RNA Isolation Kit试剂盒,反转录试剂盒、Taq酶、DNA分子质量标准、DNA凝胶回收试剂盒、pMD19-T载体、质粒提取试剂盒、限制性内切酶EcoRⅠ和SalⅠ均为TaKaRa公司产品; 蛋白质分子质量标准、异丙基-β-D-硫代半乳糖苷(IPTG)购自Sigma公司;引物合成和DNA测序分析由河南尚亚生物技术有限公司完成。
1.2 方法
1.2.1 材料种植及取样 豫红花1号及管状花颜色为白色(W)、黄色(Y)、橙色(O)、红色(R)的红花品系种植于河南省农科院现代农业科技试验示范基地,自然条件下生长。以豫红花1号为材料进行基因组织特异性表达分析,红花生长至莲座期取叶片;生殖生长期取根、茎、叶和苞片,并在管状花从花冠总苞片露出的第1、3、4、7天取管状花,分别标为S1、S2、S3、S4期;以生长一致的不同花色红花品系顶果球为材料,取开花第3天的管状花进行不同花色红花基因表达分析。所有组织部位和样品取3个生物学重复,将采集的样品立即放入液氮中保存,并转移至-80 ℃超低温冰箱中保存备RNA提取。
1.2.2 样品处理 将豫红花1号分别播种于1/2浓度的Hoagland标准营养液和培养钵中,基质为泥炭土∶蛭石=1∶1,光照培养箱中生长条件设置为(26±1)℃、16 h/8 h(光照/黑暗);待红花幼苗生长至两叶时,用20% PEG-6000对水培红花幼苗进行模拟干旱胁迫处理,用低温(4 ℃)、茉莉酸甲酯(MeJA,100 μmol/L)、脱落酸(ABA,150 μmol/L)和赤霉素(GA3,100 μmol/L)对土培红花幼苗进行喷施处理,以喷施溶剂(10%乙醇)作为对照,于处理后0、3、6、12、24 h取叶片,并立即置入液氮中保存,转移至-80 ℃超低温冰箱中保存,用于总RNA提取,每个处理设置3个生物学重复。
1.2.3CtDXS1基因cDNA全长的克隆 根据河南省农业科学院芝麻研究中心药用植物研究室构建的红花三代测序全长转录组数据库,用DNAMAN软件设计引物(表1),以豫红花1号莲座期幼苗为材料,按照华越洋生物公司的Quick RNA Isolation Kit试剂盒说明书提取总RNA,以M-MLV反转录酶(TaKaRa)试剂盒反转录合成的单链cDNA为模板进行CtDXS1基因全长克隆。PCR扩增体系为50 μL,含2×PCR buffer for KOD FX 25 μL,上下游引物(10 μmol/L)各1 μL,dNTP 5 μL,cDNA模板1 μL,KOD FX 1 μL,ddH2O 16 μL。PCR扩增程序:94 ℃变性2 min;98 ℃ 10 s,58 ℃ 30 s,68 ℃ 2 min,35个循环;68 ℃延伸10 min。将目的片段回收并连接至pMD19-T载体,转入大肠杆菌DH5α菌株,进行测序鉴定。

表1 引物序列
1.2.4 CtDXS1蛋白生物信息学分析 通过在线软件ProtParam对CtDXS1蛋白的氨基酸组成、蛋白质分子质量、理论等电点及稳定性等参数进行分析;CtDXS1蛋白的二级结构预测通过ExPASy中的SOPMA工具完成;通过SWISS-MODEL在线软件对CtDXS1蛋白的三级结构进行建模预测;通过TMHMM Server v2.0对CtDXS1蛋白的跨膜结构域进行分析;利用SignalP-5.0 server对CtDXS1蛋白的信号肽进行预测分析;利用TargetP-2.0 Prediciton对CtDXS1蛋白的亚细胞定位及转运体进行预测;利用NCBI中蛋白质结构域数据库(Conserved domain database,CDD)对CtDXS1蛋白质保守结构域进行分析;以CtDXS1蛋白序列为模板,通过NCBI数据库中BLASTP模块进行比对分析,找出拟南芥(Arabidopsisthaliana)、烟草(Nicotianatabacum)、水稻(Oryzasativa)、苜蓿(Medicagotruncatula)、黄花蒿(Artemisiaannua)等物种中CtDXS1蛋白的同源序列,利用DNAMAN对CtDXS1蛋白与其他物种的同源蛋白进行同源性分析;通过MEGA 7.0软件中的Neighbor-joining构建CtDXS1蛋白系统进化树,并通过Bootstrap方法对进化树进行检测,Bootstrap值设置为1 000。
1.2.5CtDXS1基因在不同组织和不同胁迫时间的表达分析 不同处理样品总RNA的提取参照越洋生物公司的Quick RNA Isolation Kit试剂盒说明书完成,按照天根生物科技有限公司的Fast Quant RT Super Mix说明书进行逆转录反应,根据CtDXS1基因cDNA全长序列设计荧光定量PCR引物,以Ct60s作为内参基因,引物序列如表1所示,荧光定量PCR反应参照天根生物科技公司的Super Real Pre Mix(SYBR Green)试剂盒进行,扩增体系20 μL:2×SuperReal Pre Mix 10 μL、上游引物(10 μmol/L)1.0 μL、下游引物(10 μmol/L)1.0 μL、cDNA模板5 μL、ddH2O 3 μL。荧光定量PCR反应程序:95 ℃变性20 s、55 ℃退火20 s、72 ℃延伸20 s扩增45个循环。荧光定量PCR在Eppendorf Mastercycler ep Realplex 2.2 Detection System上进行。相对定量分析采用2-ΔΔCt方法进行分析。
1.2.6CtDXS1原核表达载体的构建及异源表达 以连接pMD19-T载体并测序验证过的CtDXS1质粒为模板,用CtDXS1-PE-F、CtDXS1-PE-R引物扩增CtDXS1基因全长,回收目的片段,用EcoRⅠ和SalⅠ双酶切目的片段和pET28a原核表达载体,酶切产物纯化后,用T4连接酶将酶切目的片段和酶切载体进行连接,构建pET28a-CtDXS1原核表达载体。将pET28a-CtDXS1转化至大肠杆菌DH5α感受态细胞,挑选单克隆,通过酶切、测序对构建的原核载体进行鉴定,将鉴定正确的pET28a-CtDXS1质粒转化至E.coliTransetta(DE3)感受态细胞。挑取阳性克隆单菌落,接种于10 mL含50 mg/L羧苄青霉素LB液体培养基中,37 ℃ 180 r/min培养6 h,然后按照1∶100比例,加入到200 mL新鲜含有50 mg/L羧苄青霉素的LB液体培养基中,37 ℃ 180 r/min培养至菌液OD600达0.6左右,加入IPTG至终浓度为0.6 mmol/L,将摇床温度调至16 ℃诱导培养12 h,以16 ℃诱导培养12 h的E.coli(pET28a)作为空白对照。诱导结束后,取1 mL菌液,10 000×g离心1 min,收集菌体,加50 μL超纯水重悬,作为总蛋白待检。其余菌液通过10 000×g离心1 min收集菌体,用20 mL Binding buffer(50 mmol/L磷酸盐缓冲液,300 mmol/L NaCl,20 mmol/L咪唑,10 mmol/L MgCl2,10%甘油,pH值7.4)洗涤收集2次,最后用15 mL Binding buffer重悬,将菌体置于冰上超声破碎。破碎菌液于4 ℃、10 000×g离心20 min,将上清液经HisTrap HP纯化柱纯化、Washing buffer(50 mmol/L磷酸盐缓冲液,300 mmol/L NaCl,20、50、100、300 mmol/L咪唑,10 mmol/L MgCl2,pH值7.4)梯度漂洗和Elution buffer(50 mmol/L磷酸盐缓冲液,300 mmol/L NaCl,500 mmol/L咪唑,pH值7.4)洗脱,洗脱液利用PD10柱进行脱盐,利用Assay buffer(50 mmol/L磷酸盐缓冲液,100 mmol/L KCl,20 mmol/L MgCl2,10%甘油,2 mmol/L DTT)进行洗脱,洗脱液通过SDS-PAGE检测。
2 结果与分析
2.1 红花CtDXS1基因全长cDNA的克隆
根据红花三代测序全长转录组数据及注释分析设计红花DXS基因cDNA全长扩增引物,以红花莲座期叶片为材料,提取RNA并反转录合成单链cDNA,利用PCR方法扩增得到2 100 bp左右的片段(图1)。将PCR产物回收连入pMD19-T载体上,转化至大肠杆菌DH5α菌株,挑取单克隆进行测序,经NCBI比对分析和Open Reading Frame Finder分析,确定该序列为DXS基因cDNA全长,命名为CtDXS1,序列全长为2 136 bp,编码711个氨基酸。
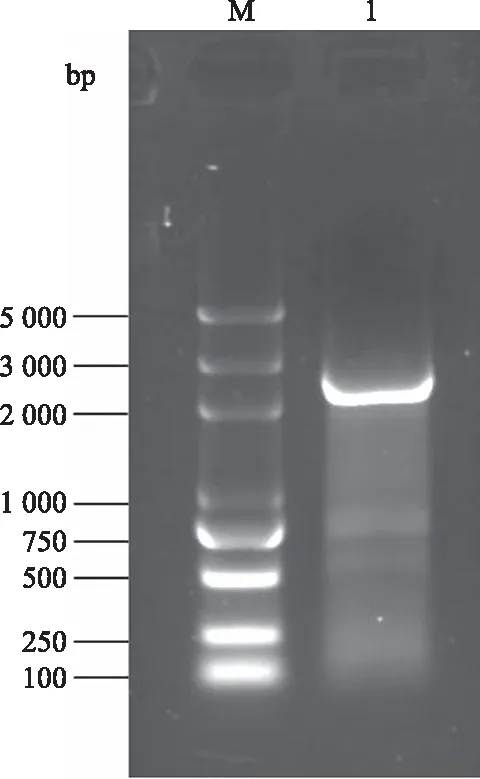
M:DL2000 plus DNA marker;1:CtDXS1基因PCR产物
2.2 CtDXS1蛋白理化性质、亚细胞定位、跨膜区分析
通过Protparam对CtDXS1基因编码蛋白质的理化性状进行预测,结果表明,CtDXS1蛋白分子质量为76 503.10 u,理论等电点为6.35,分子式为C3 384H5 328N952O1 012S30;蛋白质氨基酸组成分析表明,CtDXS1蛋白中甘氨酸含量最高,为10.4%,色氨酸含量最低,为0.3%;不稳定系数为40.07,属于稳定蛋白;脂肪系数86.33,总平均亲水性GRAVY为-0.108,为亲水性蛋白。利用SignalP-5.0 server预测CtDXS1无信号肽,属于非分泌蛋白;利用TMHMM Server v2.0对CtDXS1的跨膜区域进行分析,预测结果表明,CtDXS1没有跨膜区域,为非跨膜蛋白;利用TargetP-2.0 Prediciton对CtDXS1蛋白的亚细胞定位及转运体进行预测,结果表明,CtDXS1定位于叶绿体,并且在N端区域有50个氨基酸组成的转运肽,这与DXS参与的MEP途径发生在质体中的现象相吻合。
2.3 CtDXS1蛋白的二级结构分析及三维结构预测
通过ExPASy中的SOPMA和SWISS-MODEL分别对CtDXS1蛋白的二级结构和三级结构进行了分析,结果表明,CtDXS1蛋白的二级结构主要由α-螺旋(α-helices)组成,共有286个,占比为40.23%;其次是无规则卷曲(Random coil),共有270个,占比为38.54%;延伸链(Extended strand)97个,占比为13.64%;β-折叠(β-turn)54个,占比为7.59%。推测α-螺旋和无规则卷曲是最主要的二级结构元件,延伸链和β-折叠散布于整个蛋白质中(图2)。CtDXS1的三维结构模型结果表明,CtDXS1是由2个单体并行排列形成二聚体(图3)。
蓝色:α-螺旋;绿色:β-折叠;红色:延伸链;紫色:无规则卷曲
图3 CtDXS1蛋白的三级结构预测
2.4 CtDXS1氨基酸序列分析和系统进化树构建
利用NCBI的蛋白质保守区数据库(Conserved domain database,CDD)对CtDXS1蛋白进行保守区预测,CtDXS1蛋白具有典型的转酮醇酶的保守域,属于转酮醇酶超家族(图4),含焦磷酸硫胺素结合位点GDG(X)8E(X)4A(X)11NDN和转酮醇酶结构域DRAGX28PXD(图5)。
多重序列比对表明,CtDXS1氨基酸序列与拟南芥、烟草、水稻、苜蓿、黄花蒿DXS1氨基酸序列相似性分别为80.56%、84.24%、82.50%、84.46%、93.31%,说明CtDXS1氨基酸序列与其他植物中DXS1氨基酸序列间具有较高的同源性(图5)。
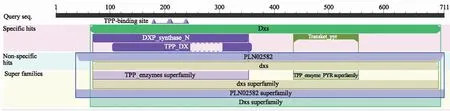
TPP:焦磷酸硫胺素
为进一步分析DXS1基因与其他植物DXS1基因的进化关系,通过NCBI蛋白质数据库和文献检索等方式,收集到其他植物的26条DXS蛋白序列,进化树分析表明,这些植物DXS序列可以聚类为3个分支,分别为CladeⅠ、CladeⅡ和Clade Ⅲ,CtDXS1属于CladeⅠ分支,与来自黄花蒿、橡胶树(Heveabrasiliensis)、蓖麻(Ricinuscommunis)和苜蓿中的DXS基因亲缘关系较近(图6), 拟南芥和苜蓿中DXS1主要行使看家基因的功能,参与叶绿体的发育和叶绿素的合成过程,但并不参与次生代谢物的合成,这暗示CtDXS1可能主要参与红花叶绿体的发育。

虚线框代表焦磷酸硫胺素结合位点,实线框代表转酮醇酶结构域
2.5 CtDXS1基因表达特性分析
分别以红花根、茎、叶、花和苞片等5个组织部位为材料研究CtDXS1基因的组织表达特性,其中,将叶分为莲座期叶和生殖生长期的叶,管状花根据发育时期分为S1、S2、S3和S4四个时期。结果表明,CtDXS1基因在苞片中表达量最高,其次是叶和茎,在根和花中也有表达但表达量相对较低,莲座期叶较生殖生长期叶表达量高;不同发育时期管状花中CtDXS1基因的定量分析表明,随着发育时期的延长,红花管状花中CtDXS1基因表达量呈明显下降趋势(图7A)。
为了进一步研究CtDXS1基因在不同花色红花中的表达量是否有差异,选取开花期S2期白色(W)、黄色(Y)、橙色(O)和红色(R)红花管状花进行CtDXS1基因定量分析,结果表明,CtDXS1基因在黄色红花中表达量最高,在白色、橙色和红色红花中表达量相对较低(图7B)。
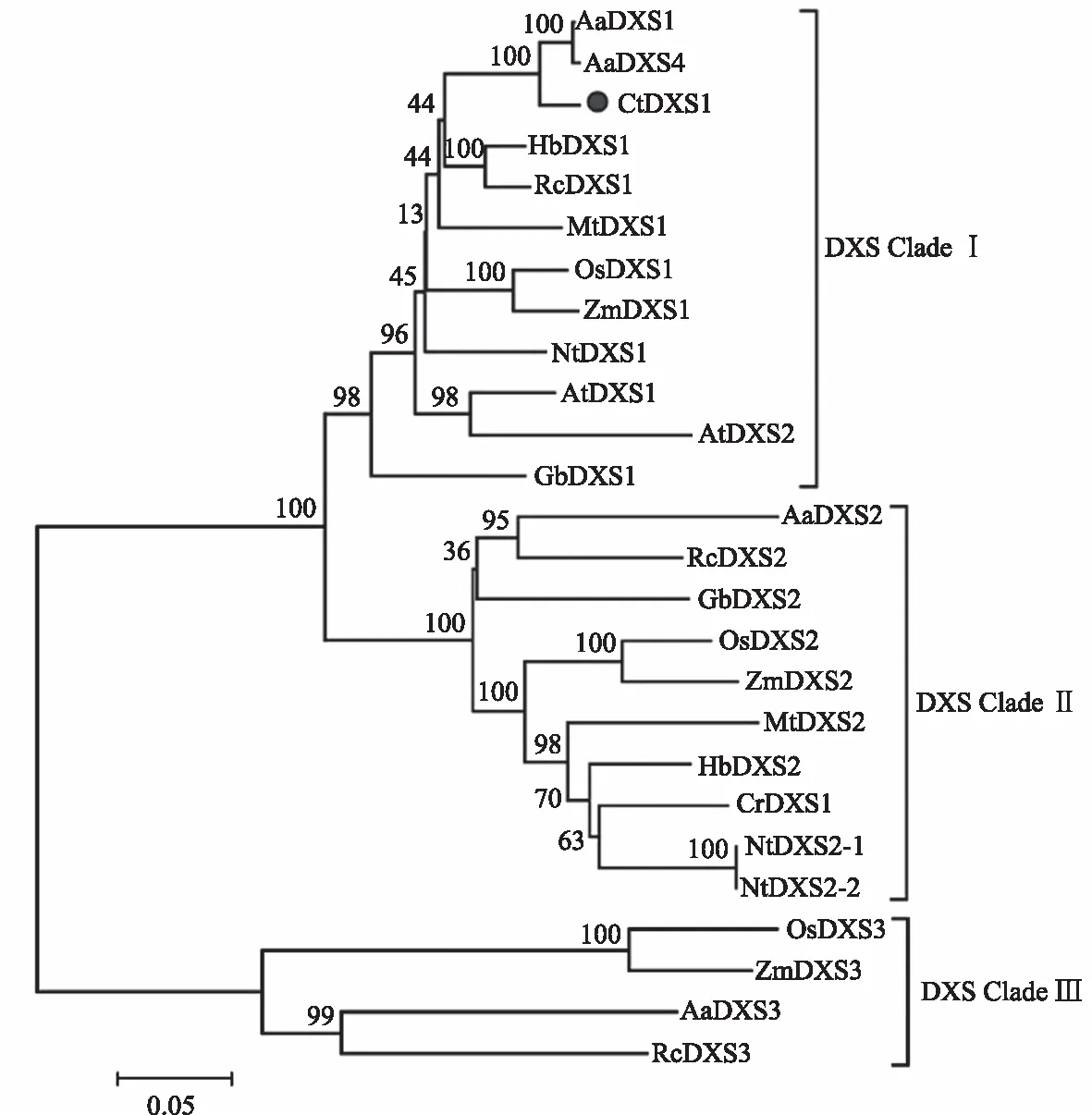
GbDXS1:Ginkgo biloba DXS1,AAS89341.1;GbDXS2:AAR95699.1;AtDXS1:AAN86173.1;AtDXS2:ACI49774.1;NtDXS1:CBA12009.1;NtDXS2-1:AFM78321.1;NtDXS2-2:NP_001312088.1;OsDXS1:NP_001055524.1;OsDXS2:NP_001059086.1;OsDXS3:BAA83576.1;CrDXS1:Catharanthus roseus DXS1,CAA09804.2;MtDXS1:CAD22530.1;MtDXS2:AAD56390.2;AaDXS1:AAD56390.2;AaDXS2:PWA79560.1;AaDXS3:PWA60387.1;AaDXS4:PWA43715.1; ZmDXS1:Zea mays DXS1,ACT32136.1;ZmDXS2:ADN22972.1;ZmDXS3:ABP88135.1;HbDXS1:AAS94123.1;HbDXS2:ABF18929.1;RcDXS1:XP_002516843.1;RcDXS2:XP_002533688.1;RcDXS3:XP_002514364.1
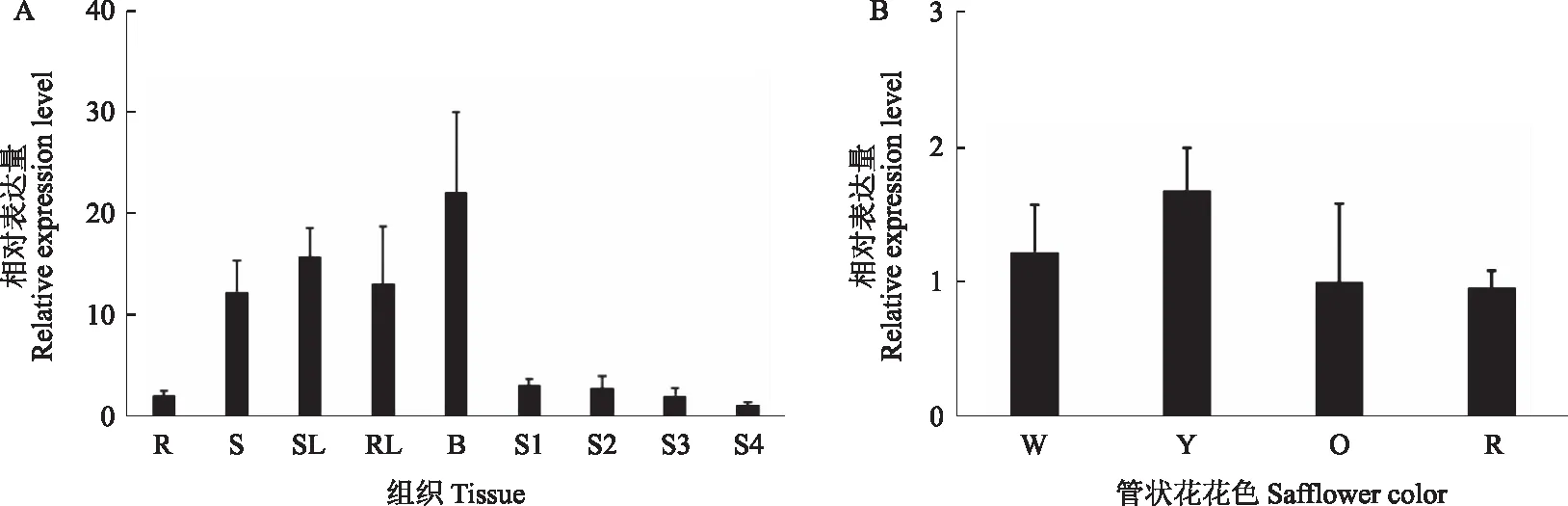
A:CtDXS1基因在不同组织中的表达模式分析(R:根,S:茎,SL:莲座期叶,RL:生殖生长期叶,B:苞片,S1:开花后1 d的管状花,S2:开花后3 d的管状花,S3:开花后5 d的管状花,S4:开花后7 d的管状花);B:CtDXS1基因在管状花颜色为白色(W)、黄色(Y)、橙色(O)和红色(R)红花品种中的表达分析
2.6 CtDXS1基因在非生物胁迫下的表达分析
为了验证CtDXS1是否在植物非生物胁迫中起作用,对红花幼苗进行干旱和4 ℃低温处理,以相同培养条件不做处理的红花作为对照,在处理后的不同时间点取样,提取RNA后进行荧光定量PCR分析。结果表明,与对照相比,模拟干旱和低温都能不同程度诱导CtDXS1基因的表达。20% PEG-6000诱导产生的干旱胁迫条件下,CtDXS1基因表达量在处理6 h后达到高峰,随后下降(图8A)。4 ℃低温处理后,CtDXS1基因表达量在处理3 h后最高,随后降低并趋于稳定(图8B)。
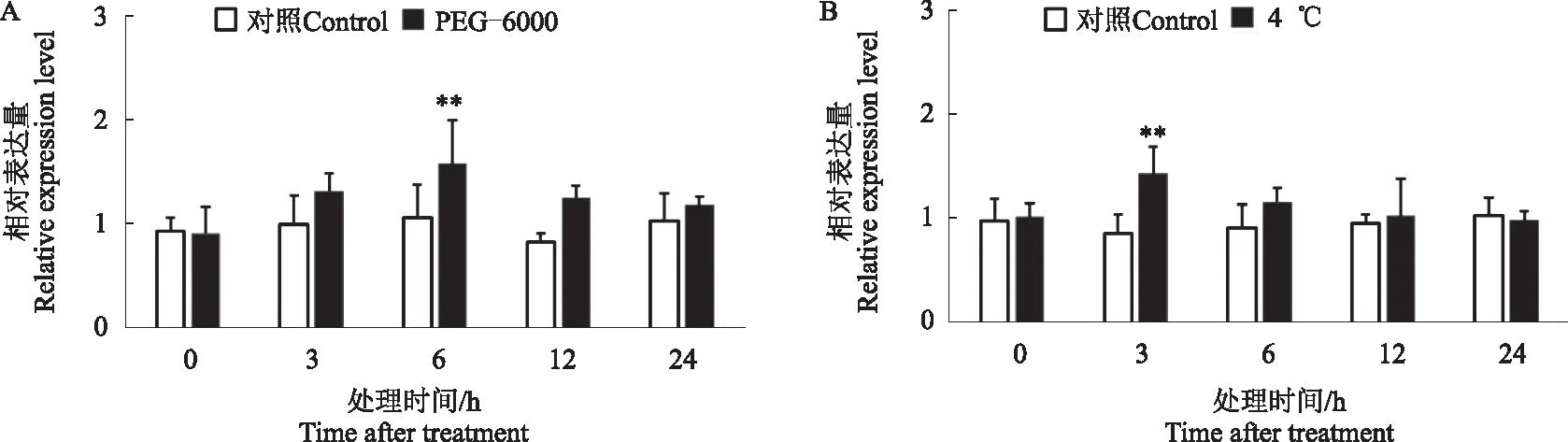
**表示与对照相比差异显著(P<0.01),下同
2.7 CtDXS1基因在不同激素诱导下的表达分析
为了研究CtDXS1基因表达是否受激素调控,对红花幼苗进行MeJA、GA3和ABA处理,通过荧光定量PCR对其表达变化进行定量分析,结果表明,与对照相比,MeJA对CtDXS1基因的表达具有一定诱导作用,其表达量在激素处理6 h达到高峰,随后下降;而GA3和ABA对CtDXS1基因表达具有抑制作用,GA3处理3 h,CtDXS1基因表达量降至最低,随后上升,但仍低于对照,而ABA处理12 hCtDXS1基因表达量降至最低(图9)。
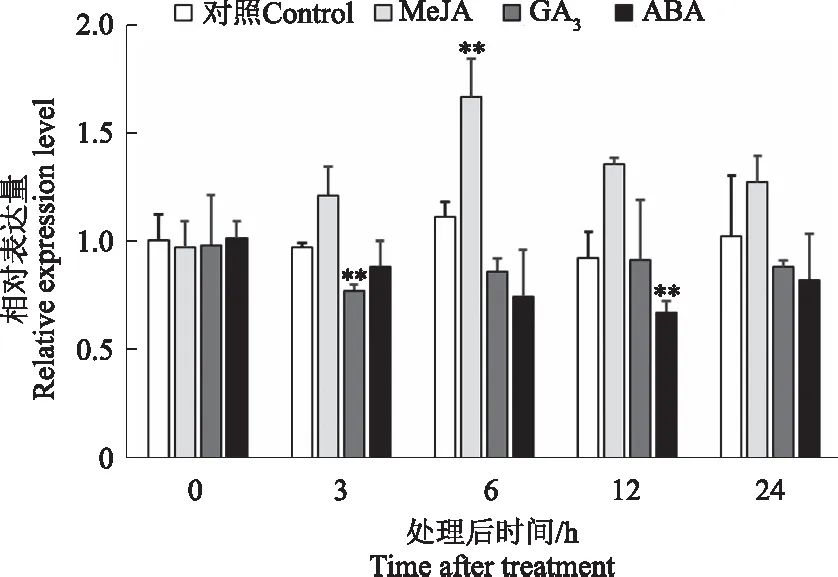
图9 CtDXS1基因在不同激素处理下表达变化
2.8 CtDXS1基因的原核表达分析
将构建好的原核表达载体pET28a-CtDXS1质粒转入大肠杆菌E.coliTransetta(DE3)后,通过加入IPTG至终浓度为0.6 mmol/L,在16 ℃诱导培养12 h后,通过SDS-PAGE检测发现,在76 ku处有大量目的蛋白条带,而空白对照载体没有出现目的条带,根据pET28a质粒的His标签对目的蛋白进行纯化,通过SDS-PAGE对纯化蛋白进行检测发现,其大小与预测的CtDXS1大小相符,确认其为目的蛋白(图10)。
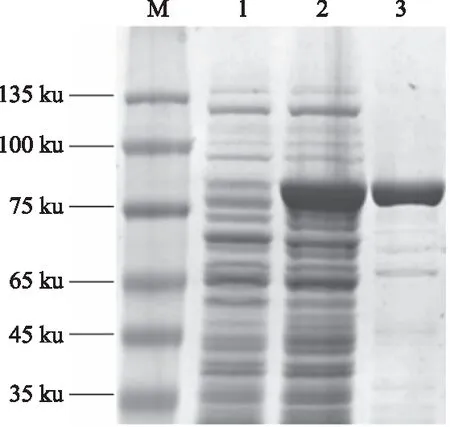
M:蛋白质Marker,1:pET28a空载体,2:pET28a-CtDXS1重组蛋白,3:纯化后的CtDXS1重组蛋白
3 结论与讨论
MEP是位于植物质体中异戊二烯生物合成途径,DXS是该途径中的第一个限速酶[23],DXS在植物生长发育和环境适应方面都起着重要作用,另外,DXS也是次生代谢产物叶绿素、维生素E、类胡萝卜素、赤霉素和萜类等化合物的生物合成途径的第一限速酶[24]。DXS基因最早从大肠杆菌(E.coli)[25]中分离出来,截至目前,已经从100多种植物中克隆到了DXS基因[26],但尚未见到红花中DXS基因的相关报道。为了研究红花DXS基因序列特征,根据转录组数据,从红花叶片中克隆得到1条开放阅读框为2 136 bp的红花DXS基因序列(CtDXS1),对其进行了生物信息学分析,分析表明,CtDXS1基因具有DXS家族必需的质子传递域和硫胺焦磷酸结合域,并且在不同物种中高度保守。
目前发现的MEP途径相关蛋白酶都定位于质体中,对其他DXS基因的研究发现,大部分DXS蛋白在N端都有30~60个氨基酸残基组成的质体转运肽[27-29],如在番茄中转化番茄DXS融合GFP表明,该蛋白质定位于叶绿体[30]。本研究通过生物信息学分析表明,CtDXS1蛋白在N端有50个氨基酸组成的转运肽,这与DXS定位于质体相一致,暗示CtDXS1基因也能参与叶绿体的形成,对植物的正常生长发育以及次生代谢产物的合成具有很重要的作用。
植物DXS家族基因根据其结构及功能可以分为3种类型。Ⅰ型为管家基因,其所编码的酶催化前体物质形成叶绿素、类胡萝卜素等与光合有关的萜类物质,如拟南芥中AtDXS1(CLA1)基因功能缺失突变体表现出白化现象[31]。Ⅱ型基因编码植物特异性次生代谢产物,如苜蓿MtDXS2可能跟植物的防御功能有关[32],Ⅲ型基因所编码酶的功能尚不清晰。本研究中,利用其他植物DXS家族成员构建系统进化树发现,CtDXS1与其他物种的DXS1聚为一类,并且与黄花蒿AaDXS1亲缘关系最近。通过不同组织部位表达分析发现,CtDXS1基因主要在茎、叶、苞片等叶绿素较多的部位表达,这暗示着CtDXS1基因可能参与红花叶绿素的合成。另外,通过不同花色红花中CtDXS1基因的定量分析发现,CtDXS1基因在黄色红花中表达量较高,说明CtDXS1基因可能也参与红花类胡萝卜素的生物合成。
研究表明,DXS受多种激素和环境信号因子的调控[33],如赤霉素、水杨酸、茉莉酸甲酯和油菜素内脂处理铁皮石斛后,原球茎中DXS的基因转录水平升高[34];在白木香(Aquilariasinensis)中的研究发现,AsDXS1受物理、化学和H2O2的调控,AsDXS2和AsDXS3只受物理和化学调控而不受H2O2调控,但MeJA处理能够诱导3个基因的表达[35],说明不同DXS基因对激素及各种环境、胁迫的响应不同。本研究表明,CtDXS1在受到干旱和低温胁迫时表达量上升,茉莉MeJA能诱导CtDXS1基因的表达,而GA3和ABA对CtDXS1基因表达有一定的抑制作用。在玫瑰的研究中发现,DXS基因表达与花朵发育程度密切相关,随着花朵的衰败,表达量降低[36]。本研究分析开花后不同时间的红花花朵CtDXS1基因表达量发现,随着红花冠状花的衰老,CtDXS1基因表达量也呈现下降趋势。
利用原核表达诱导产生目的蛋白,通过体外酶活试验手段对其催化功能进行研究已成为鉴定植物次生代谢相关酶功能的主要手段[37],本研究通过构建CtDXS1原核表达载体将其在大肠杆菌中进行诱导表达,并将目的蛋白进行纯化,经检测与目的蛋白大小一致,为后续CtDXS1的功能鉴定奠定了基础。
综上,从红花中克隆得到CtDXS1基因的全长序列,长度2 136 bp,编码711个氨基酸,该蛋白质具有典型的转酮醇酶家族功能域。系统进化分析显示,CtDXS1与来自黄花蒿的DXS亲缘关系最近,属于DXS基因家族的Ⅰ类基因。CtDXS1基因在苞片中表达量最高,其次是叶和茎,在其他组织部位表达量较低。CtDXS1基因在管状花为黄色的红花中表达量较高。干旱、低温胁迫能够诱导CtDXS1基因表达上调;MeJA诱导CtDXS1基因的表达,而GA3和ABA对CtDXS1基因表达有一定的抑制作用。另外,通过原核表达诱导获得并纯化了目的蛋白。
