养殖密度对2龄刀鲚生长的影响
2021-03-13刘永士王建军朱建明陆根海施永海刘建忠
刘永士,王建军,朱建明,陆根海,施永海,刘建忠
(上海市水产研究所 上海市水产技术推广站,上海 200433)
刀鲚Coilianasus隶属于鲱形目Clupeiformes鳀科Engraulidae鲚属Coilia,为江海洄游性鱼类,主要分布于中国黄渤海和东海一带,凡通海的江河均有分布,以长江下游产量最高[1],刀鲚曾占到长江鱼类天然捕捞量的30%~50%[2],自20世纪90年代起,在江湖阻隔、水域污染加剧和长期高强度捕捞等多重影响下,长江刀鲚资源急剧衰退,洄游区间大幅缩短,分布区域日趋萎缩,各江段捕捞量也持续大幅下滑[3],2001年以后更是急剧下滑,已不能形成鱼汛[4],市面上长江刀鲚价格居高不下,甚至出现“有价无市”的窘境,从2019年开始长江流域禁捕长江刀鲚。因此,开展刀鲚的人工养殖以解决刀鲚市场供应不平衡问题已迫在眉睫。
2011年,上海市水产研究所率先获得了刀鲚室内集约化人工繁殖的成功,并于2013年实现了刀鲚苗种室内全人工繁育和规模化生产[1,5],这为开展刀鲚人工养殖奠定了坚实的苗种基础。目前,刀鲚养殖主要采用活体生物饵料,如枝角类、桡足类、鱼或虾的幼体及糠虾目Mysidacea甲壳动物等[6-11],虽然也有利用人工配合饲料投喂刀鲚的报道[5,8-9,12],但均仅处于试验阶段,尚未应用于实践生产中[5],因此,如何获得足量且适口的天然活体饵料,已成为决定刀鲚养殖成败的关键。活体饵料来源不稳定,根据实际情况选定活体饵料的种类并确定养殖密度是刀鲚人工养殖的基础。为此,本试验中采用浮游动物(枝角类和桡足类等)和日本沼虾作为饵料生物,研究了不同养殖密度下2龄刀鲚的生长,旨在为开展刀鲚产业化养殖提供基础,并为刀鲚养殖模式的发展提供科学参考。
1 材料与方法
1.1 材料
1.1.1 试验用鱼和水 试验用2龄刀鲚为上海市水产研究所奉贤科研基地全人工繁育的刀鲚苗种,经室内强化培育、室外池塘培育、越冬养殖、室外池塘养殖后,于次年4月拉网取出,试验鱼体长为(12.63±1.55)cm,体质量为(4.43±1.25)g。试验用水为当地河水。
1.1.2 池塘条件 养殖池塘位于上海市奉贤区星火农场养殖区域(杭州湾北岸),为相邻的3口池塘(编号:8#北、8#南、9#北),每口池塘面积为4×667 m2,水深1.5 m,呈规整的长方形,东西走向,进排水方便,进水口用0.245 mm筛绢网过滤,排水口网眼为围网(2 mm×3 mm)和闸网(0.37 mm)。利用网片(网眼为2 mm×3 mm)南北向固定于池塘中(底部封土固定,上部高出水面50 cm),将每口池塘均分为4个围隔,每个围隔面积667 m2,每667 m2配备叶轮式增氧机(1.5 kW)1台,池塘上方覆盖尼龙网以防止鸟类偷食。池塘使用前,清塘消毒,池底浸泡消毒24 h后,彻底排干曝晒待用。
1.2 方法
1.2.1 试验设计及鱼种放养 鱼种放养前,池塘要提前培养基础饵料生物,根据放养鱼种时间,提前5~7 d进水,先进水0.5 m水位,后根据水色情况和饵料生物量等逐步添加至1.5 m水位,5~7 d后水色变浓,水体中出现枝角类及桡足类等饵料生物,待饵料生物量达到20~40 ind./L,放养2龄刀鲚鱼种。
选择晴好天气在池塘上风处放苗,提前开启增氧机,使池水不分层,放养鱼种要求规格整齐、鳞片完整且活力较好。
2龄刀鲚养殖设置3个密度梯度,分别为500 ind./667 m2(8#北,组A)、1 000 ind./667 m2(8#南,组B)和1 500 ind./667 m2(9#北,组C),每个养殖密度设3个平行(即每个池塘仅用上述4个围隔中的3个,剩下1个空置),各密度组刀鲚放养后暂养7 d,期间每天捞出死鱼并记录,7 d后根据各试验组死鱼数补入同批2龄刀鲚,之后养殖试验开始计时。
1.2.2 饵料生物投放 除了鱼种放养前培育饵料生物外,在刀鲚放入池塘的7 d内,陆续向每个试验组中添加小型日本沼虾Macrobrachiumnipponense作为其饵料生物,后期根据养殖塘内日本沼虾数量适当补充。
养殖周期内,投放日本沼虾量分别为:A组10 kg、B组15 kg、C组20 kg,并且每天投喂适量南美白对虾配合饲料作为日本沼虾的饲料。
1.2.3 日常管理 每晚开启增氧机8~10 h,晴好天气中午开启增氧机1 h,天气恶劣时,增加开机时间;每天(8:00、16:00)测量两次水温,养殖期间水温为15.2~31.4 ℃,各阶段平均水温见图1;每14 d换水1/3。养殖从4月5日开始到10月11日结束,共189 d。

图1 2龄刀鲚养殖期间各月份平均水温变化Fig.1 Change in mean water temperature in each month during 2-year-old tapertail anchovy Coilia nasus culture
1.2.4 取样及分析 试验期间,每19~30 d取样一次,从每个平行随机取样20尾,测定其体长与体质量。养殖试验结束后,从每个平行取样10尾,先饥饿处理24 h,测定体长与体质量,后置于冰盘中解剖,取出内脏团,分别测定内脏团和肝的质量及肠长,然后放入事先预冷的离心管中备用。
1.2.5 指标的计算 肥满度(CF)、比肠长(RGL)、脏体比(VSI)、肝体比(HSI)、变异系数(CV)、特定生长率(SGR)计算公式分别为
肥满度=m/L3×100%,
比肠长=Li/L,
脏体比=mv/m×100%,
肝体比=mh/m×100%,
变异系数=s/x×100%,
特定生长率=(lnm2-lnm1)/(t2-t1)×100%。
其中:L为体长(cm);Li为肠长(cm);t为养殖时间(d);m为体质量(g);mv为内脏团质量(g);mh为肝质量(g);s为标准差;x为平均值;m2、m1分别为养殖时间t2、t1时的体质量(g)。
1.2.6 体质量与体长关系及生长模式分析 分别采用一次线性函数分析体长、体质量与养殖时间的关系,采用幂函数分析体质量与体长的关系。
体长与养殖时间的关系式为
L=at+b;
体质量与养殖时间的关系式为
m=at+b;
体质量与体长的关系式为
m=aLb。
其中:m为体质量;L为体长;t为养殖时间;a、b为常数。
1.3 数据处理
试验结果用SPSS 20软件进行处理,采用单因素方差分析(One-way ANOVA)和Duncan新复极差法对各组数据进行显著性差异分析和多重比较。采用SPSS 20软件对各回归方程进行显著性检验,显著性水平设为0.05,极显著性水平设为0.01。
2 结果与分析
2.1 2龄刀鲚的生长性能
从表1可见:3个试验组刀鲚均采用同一批2龄鱼种,其初始体质量和体长相同,经过189 d的养殖后,A组末体质量、末体长、增重率、体长增长率、特定生长率均显著优于B组和C组(P<0.05);A组成活率显著高于C组(P<0.05),但与B组无显著性差异(P>0.05);A、B、C组单位面积产量分别为8.59、7.28、11.33 kg/667 m2,尽管C组产量较A组和B组分别提高了31.9%和55.6%,但3组间无显著性差异(P>0.05)。

表1 不同养殖密度下2龄刀鲚的生长指标Tab.1 Growth indices of 2-year-old tapertail anchovy Coilia nasus cultured at different densities
2.2 2龄刀鲚形体指标
从表2可见:养殖密度未对2龄刀鲚肥满度、脏体比和比肠长造成显著影响(P>0.05);C组肝体比显著高于B组(P<0.05),但与A组无显著性差异(P>0.05)。
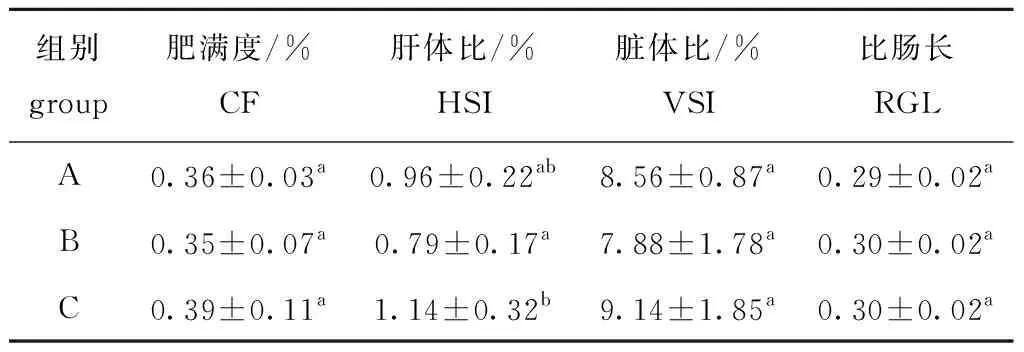
表2 不同养殖密度下2龄刀鲚的形体指标
2.3 2龄刀鲚体质量与体长的关系
从图2可见:不同养殖密度下,2龄刀鲚体质量与体长均呈良好的幂函数关系;A组的b值(3.076 8)接近3,为等速生长;B组和C组b值分别为3.429 1和3.534 3,均大于3,为异速生长,体质量增长略快于体长增长。

图2 不同养殖密度下2龄刀鲚体质量与体长的关系Fig.2 Relationship between body mass and body length of 2-year-old tapertail anchovy Coilia nasus cultured at different densities
2.4 2龄刀鲚一般生长型
从图3可见:不同养殖密度下,2龄刀鲚体长、体质量随养殖时间均呈线性增长关系;A组与C组体长生长直线相交于17.4 d(5月30日),A组与B组体长生长直线相交于55.3 d(5月30日),B组与C组生长直线不相交;A组与C组体质量生长直线相交于19.9 d(4月25日),A组与B组体质量生长直线相交于40.6 d(5月16日),B组与C组生长直线不相交。

图3 不同养殖密度下2龄刀鲚体长、体质量与养殖时间的关系Fig.3 Relationships between body length or body mass and culture time of 2-year-old tapertail anchovy Coilia nasus cultured at different densities
2.5 2龄刀鲚生长离散与体长、体质量分布
从表3可见:B组的体长与体质量变异系数最小,说明B组2龄刀鲚生长离散小,规格较均匀,但与A组和C组无显著性差异(P>0.05)。
对养殖189 d的60尾2龄刀鲚做体长与体质量分布图(图4和图5),3个组刀鲚体长与体质量范围分别为A组:15.3~27.1 cm、12.32~76.24 g;B组:12.8~23.6 cm、12.67~43.28 g;C组:12.5~23.4 cm、8.92~59.54 g。A组体长为18.1~22.9 cm的个体占样本总数的90.0%,体质量为25.31~40.94 g的个体占样本总数的88.3%;B组体长为16.6~20.9 cm的个体占样本总数的88.3%,体质量为15.61~30.24 g的个体占样本总数的93.3%;

表3 不同养殖密度下2龄刀鲚体质量、体长变异系数

图4 不同养殖密度下2龄刀鲚体长分布Fig.4 Distribution of body length of 2-year-old tapertail anchovy Coilia nasus cultured at different densities

图5 不同养殖密度下2龄刀鲚体质量分布Fig.5 Distribution of body mass of 2-year-old tapertail anchovy Coilia nasus cultured at different densities
C组体长为16.90~21.50 cm的个体占样本总数的81.7%,体质量为18.61~36.88 g的个体占样本总数的83.3%。
3 讨论
3.1 养殖密度对刀鲚生长性能的影响
养殖密度是鱼类生长过程中主要的影响因子[13-14]。不同鱼类的养殖密度有一定的阈值,当养殖密度低于阈值,鱼类成活率不受影响;当养殖密度超过阈值,随着养殖密度升高,鱼类成活率呈下降趋势[15]。本研究中,A组(500 ind./667 m2)成活率显著高于C组(1 500 ind./667 m2),而A组和B组(1 000 ind./667 m2)成活率无显著性差异,这说明2龄刀鲚养殖密度的阈值为1 000 ind./667 m2,然而此阈值是否是2龄刀鲚适宜的养殖密度,仍需结合刀鲚的生长过程进行验证。
研究表明,养殖密度过高或过低均会对鱼类生长产生抑制,高养殖密度加剧鱼类对养殖水域空间、食物资源的争夺,且易引起水质恶化,增加相关疾病发生概率,降低鱼类成活率及群体生长率[16-18];过低养殖密度也会降低一些鱼类的生长性能,如俄罗斯鲟鱼Acipensergueldenstaedtii[19-20]、宝石鲈Scortumbarcoo[21]、点带石斑鱼Epinephelusmalabaricus[16]等,这是因为对于群居性鱼类而言,集群生活可帮助生物更有效地防御敌害,寻找食物,从而降低维持生存所消耗的能量,将更多的能量用于生长[22]。本试验中,放养刀鲚的初始规格相同,经过189 d养殖,A组的末体长、末体质量、体长增长率、增重率、特定生长率等生长指标均优于B、C组,且3个组产量无显著性差异,说明2龄刀鲚的养殖密度为500 ind./667 m2时未对其集群行为产生负面影响,且获得较好的生长性能。因此,在本试验条件下(初始生物饵料密度均为20~40 ind./L,养殖过程中陆续添加小型日本沼虾10 kg),500 ind./667 m2是2龄刀鲚适宜的养殖密度。此外,刀鲚人工养殖中严重依赖活体生物饵料,且尚未实现适宜刀鲚各生长阶段摄食活体生物饵料的全年充足供应,这也成为抑制刀鲚养殖系统生产潜力的关键因素。针对上述情况,笔者认为当前的主要解决途径有:
1)开发多种活体生物饵料[6-11],满足刀鲚全年摄食需要;
2)开发刀鲚人工配合饲料,目前已有学者进行了此类研究[5,8-9,12],但尚未取得实质性突破;
3)充分利用刀鲚养殖水体,挖掘养殖系统的生产潜力,发展多种养殖模式,如建立刀鲚、河蟹和日本沼虾混养模式[23]等。
本试验中,低密度组2龄刀鲚成活率高、生长好,可能是因为低密度组饵料相对丰富,若可实现适口饵料的充足供应,尤其是开发出适合刀鲚摄食的人工配合饲料,将会从根本上改变刀鲚目前的养殖方式。研究表明,配合饲料或配合饲料与动物性饵料搭配使用可显著提高台湾泥鳅Paramisgumusdabryanusssp.[24]仔鱼和大黄鱼Pseudosciaenacrocea[25]稚鱼的生长与成活率, 这主要是因为配合饲料营养较为均衡,且可根据不同养殖品种的营养需求进行定制,以弥补长期摄食动物性饵料导致的营养缺乏。施永海等[8]研究表明,配合饲料饲养的刀鲚蛋白质品质略优于活饵料饲养的刀鲚,由此可推测,本试验中2龄刀鲚长期摄食活体生物饵料势必出现营养不均衡的情况,且某一地区养殖刀鲚的饵料生物往往趋于单一,如本试验中2龄刀鲚除摄食浮游动物外,主要摄食日本沼虾,极易导致营养缺乏,这也可能是导致2龄刀鲚养殖成活率普遍偏低的原因,因此,开发刀鲚适口的配合饲料不仅可提供不间断充足的饵料供应,从而提高养殖系统的生物容量,同时也可根据刀鲚的摄食和营养需求定制营养更均衡的饲料,提高刀鲚的成活率,从而提高产量,但这仍需要持续不断的深入研究。
3.2 养殖密度对形体指标的影响
研究表明,硬骨鱼类的肝体比一般为1%~2%[26],这与本试验中刀鲚(0.79%~1.14%)相似。比肠长可反映鱼类的食性,肉食性鱼类肠道较短,为体长的1/3~3/4,杂食性或草食性鱼类的肠道较长,可达体长的2~5倍,甚至可达15倍[27-28],如肉食性鱼类褐鲳鲉Sebastiscusmarmoratus[29]、军曹鱼Rachycentroncanadum[30]、波纹唇鱼Cheilinusundulatus[31]、兰州鲇Siluruslanzhouensis[32]的比肠长分别为0.54、0.43、0.43和0.912;草食性鲢Hypophthalmichthysmolitrix、鳙Aristichthysnobilis比肠长分别为6.82~13.45和4.84~6.83[26];杂食性弗氏兔脂鲤Leporinusfriderici比肠长为1.25[33]。本试验中2龄刀鲚比肠长为0.29~0.30,说明刀鲚为肉食性鱼类,其数值略高于刀鲚幼鱼的比肠长(0.241±0.010)[34],这可能与刀鲚不同发育阶段及摄食活体饵料组成结构变化有关。
鱼类的消化道指数,如比肠长、肝体比、脏体比等,与鱼类的消化能力密切相关[32],并受到以下多种因素的影响:
1) 个体发育,如菊黄东方鲀Takifuguflavidus比肠长随着个体生长呈显著下降趋势,而肝体比则反之[35];
2) 饵料性质,如河鲈Dicentrarchuslabrax长期摄食低质难消化的食物时,其肠道系统会更长[36];
3) 营养结构,如台湾泥鳅Taiwanloach的脏体比和肝体比随饲料蛋白质水平的提高呈先升高后趋于稳定的趋势[37];
4) 养殖模式,如采用池塘内循环流水养殖的黄颡鱼Pelteobagrusfulvidraco脏体比和肝体比显著低于常规池塘养殖。
上述除了因素1)以外,均可通过改变摄入营养的形态或改变营养在体内积累和消耗方式,改变鱼类的消化道指数。本试验中,不同养殖密度未对2龄刀鲚脏体比产生显著影响,但C组肝体比显著高于B组,这可能是因为高密度组试验养殖前期一部分刀鲚死亡,养殖后期刀鲚养殖密度降低,但仍以高密度组来添加日本沼虾,造成饵料生物丰富,摄食的营养在肝脏中累积,但该推测仍需进一步试验验证。
3.3 养殖密度对体质量与体长关系的影响
鱼类的体质量与体长关系可用公式m=aLb表达[38],式中a为条件因子,反映种群所处环境的优劣[39],b为异速生长因子,反映生长发育的不均匀性,如b<3为负异速生长,体长增长快于体质量的增加,如b=3为等速生长,体长和体质量增加等速,如b>3则为正异速生长,体质量的增加快于体长的增长[40]。衷思剑等[41]研究表明,黄鮟鱇Lophiuslitulon在生长条件较好情况下,a值往往更大。本试验中,2龄刀鲚体质量与体长关系可用公式m=aLb表达,式中a值随养殖密度增加逐渐减小,A组的a值(0.002 8)最大,说明在此密度下2龄刀鲚所处的生长环境更适宜。研究表明,大部分鱼类的b值分布在2.5~3.5[42]和2.7~3.4[40],幼鱼阶段b值多小于3,成鱼时b值接近或大于3[38]。本试验中,2龄刀鲚b值(3.076 8~3.534 3)与上述研究结果相似,但其随养殖密度的增加而逐步增大,说明养殖密度的提高加大了2龄刀鲚生长的不均匀性,这可能是因为养殖前期高密度组刀鲚对饵料及空间的竞争激烈,造成生长放缓;养殖中后期,一部分刀鲚死亡,养殖密度降低,此时添加一定量的活体饵料,2龄刀鲚竞争强度降低,生长加快。另外,2龄刀鲚已性成熟,养殖中后期摄食的能量更多用于性腺发育,这也导致了体质量的快速增长。根据体质量与体长关系式可知,放养密度为500 ind./667 m2时,2龄刀鲚的生长条件更优,体长与体质量生长更均衡。
3.4 养殖密度对体长、体质量生长模式的影响
本试验条件下,2龄刀鲚的体长与体质量生长均可采用一次线性函数拟合,表现为匀速生长,而徐钢春等[10]研究表明,大规格刀鲚体长采用一次线性函数拟合,这与本试验结果相一致,而体质量生长则采用指数函数拟合,出现加速生长情况,这主要是该研究中刀鲚摄食的饵料包括浮游动物(轮虫、枝角类和桡足类)及小型日本沼虾、虾虎鱼等,饵料来源较本研究更丰富且充足,为刀鲚生长提供了较充足且均衡的营养,说明饵料来源与丰度可影响刀鲚的生长模式。
养殖密度可影响2龄刀鲚的生长速度,本研究中刀鲚初转入新环境,集群生活可帮助其更有效防御敌害,寻找食物,从而降低维持生存所消耗的能量,将更多的能量用于生长[22],因此,低密度组(500 ind./667 m2)刀鲚的体长与体质量生长在养殖初期均低于高密度组,随着2龄刀鲚对环境的适应,以及高密度组刀鲚对空间和饵料的竞争加剧,不同密度组刀鲚的生长发生变化,A组刀鲚体长与体质量分别在养殖17.4 d和19.9 d后超过C组,在养殖55.3 d和40.6 d后超过B组。因此,今后如短期暂养刀鲚(如暂养放流鱼种),可选择高密度放养模式。
3.5 养殖密度对生长离散的影响
养殖密度较高加剧了种内对生存空间和饵料资源的竞争,导致资源在养殖系统内部的不均匀分配,处于生长优势的鱼生长加快,生长劣势鱼的生长率进一步下降,造成鱼群呈现生长离散现象[43]。有研究表明[44],种群密度增加,导致对食物和空间的竞争,加速鱼群大小分布的差异和不对称性,有可能在种群内部形成社会等级制度,使生长不均匀。本试验中,尽管不同养殖密度下2龄刀鲚体长、体质量变异系数无显著性差异,但体长变异系数较小,体质量变异系数较大(表3),这与对白点鲑Salvelinusleucomaenis[45]和俄罗斯鲟Acipensergueldenstaedti[44]幼鱼的研究结果相似, B组体长与体质量变异系数明显小于其余两组(表3),原因是A组养殖密度较小,生存空间和饵料资源压力相对较小,生长优势鱼最大体长与体质量分别达到27.10 cm和76.24 g,远大于其余两组,从而导致A组的体长、体质量变异系数大于B组,但A组体长与体质量分布相对集中(图4、图5),且整体规格要大于其余两组,由此可见,采用活体饵料投喂2龄刀鲚,在低密度条件下,虽然生长优势鱼易占据有利资源,生长较快,但并不影响同组其他鱼的生长也优于高密度组,且体长与体质量分布区域相对集中。
4 结论
1)在本试验条件下(初始生物饵料密度为20~40个/L,养殖过程中陆续添加小型日本沼虾10 kg),2龄刀鲚的适宜养殖密度为500 ind./667 m2。
2)2龄刀鲚体长与体质量均呈幂函数相关m=aLb,b值随着养殖密度增加而逐步增大;体长与体质量生长均可采用一次线性函数来拟合,表现为匀速生长。
3)2龄刀鲚养殖密度为500 ind./667 m2时,体长与体质量分布区域更集中。
