有害赤潮藻赤潮异弯藻Heterosigma akashiwo 研究进展
2021-03-11姬南京张珍珍黄金旺
姬南京 , 张珍珍 黄金旺 申 欣
(1. 江苏海洋大学 江苏省海洋生物资源与环境重点实验室/江苏省海洋生物技术重点实验室, 江苏 连云港222005; 2. 江苏海洋大学 江苏省海洋生物产业技术协同创新中心, 江苏 连云港 222005; 3. 江苏省海洋资源开发研究院, 江苏 连云港 222005; 4. 中国科学院海洋研究所 海洋生态与环境科学重点实验室, 山东 青岛 266071)
赤潮通常是指一些海洋微藻、原生动物或细菌在水体中过度繁殖或聚集造成水体变色的现象[1]。在海洋系统中, 微型藻类不仅在食物链底端为其他水生生物提供食物, 而且在全球碳循环中也扮演着重要角色, 它们通过光合作用吸收大气中的二氧化碳(CO2), 从而起到固碳作用。然而, 在特定条件下, 某些种类暴发性增殖并聚集或产毒会形成有危害的藻华, 因这类藻华通常会造成水体呈红色(主要由甲藻引起), 所以最早被泛称为赤潮。由于不同物种(浒苔、马尾藻、抑食金球藻等)导致的海水变色不同, 而后又分为绿潮、金潮、褐潮等。近年来, 国际科学界将上述这些造成危害的生态现象统称为有害藻华(Harmful algal bloom, HAB)[2-3]。
赤潮异弯藻(Heterosigma akashiwo)是形成有害藻华的主要物种, 在世界多个海域均有该物种的分布,该物种之所以引起高度关注, 主要因为其藻华发生时会造成鲑鱼、黄尾鱼等养殖鱼类大面积死亡[4]。近年来, 随着CO2水平升高、全球气候变暖、水体富营养化程度日趋严重, 有害藻华的发生规模和频次总体呈逐年增加的趋势, 且相关研究表明赤潮异弯藻在全球气候变化的大背景下可能成为赤潮优势种[5]。鉴于赤潮异弯藻的危害性及其藻华的发生趋势, 开展该物种藻华发生机制研究变得尤为重要。本文针对赤潮异弯藻物种鉴定、基础生物学研究、生态学研究等几个方面的研究进展进行综述, 以期为该物种的藻华发生机制研究、藻华防治等工作提供参考。
1 赤潮异弯藻的发现和地理分布
赤潮异弯藻属于针胞藻纲、异弯藻属, 个体较小,长10~25 μm, 宽8~15 μm, 形状多变, 呈卵圆形或椭圆形, 有两根鞭毛, 细胞内含有岩藻黄素(fucoxanthin)和紫黄质(violaxanthin), 含有10~15 个叶绿体, 在显微镜下为金褐色[6]。该藻为广温性、广盐性生物, 在盐度2‰~50‰范围内均可存活, 在温度14~30 ℃之间有较高的生长率(不同株系之间存在差异), 在美国、日本、新西兰、澳大利亚、新加坡、中国等世界多个海域均有该物种的分布。1985—1987 年(6—8 月)在我国大连湾首次报道赤潮异弯藻藻华, 其后该物种形成的藻华在我国胶州湾、东海等多个海域均曾被发现[7]。2017 年在我国南黄海35°N 断面甚至出现了绿潮、金潮和赤潮三类有害藻华共发现象, 其中赤潮主要由米氏凯伦藻(Karenia mikimotoi)和赤潮异弯藻形成[8]。
几十年来, 有关赤潮异弯藻的命名和分类地位争议不断, 该物种曾被误以为是金色滑盘藻(Olisthodicusluteus)。根据现有文献记载, 1965 年Huiburt 首次在美国马萨诸萨州发现了赤潮异弯藻, 将其命名为Olisthodicus carterae, 两年后研究者Hada在日本也发现了类似的物种, 先后将其命名为Entomosigma akashiwo和H.akashiwo。但根据国际植物命名法, 这一系列名字均属无效, 因为Huiburt 和Hada 都没有提供该物种的主模式标本(holotype), 直到1987 年, 该物种才获得有效的命名H. akashiwo[9]。2016 年, 有研究者在已鉴定的赤潮异弯藻24 个株系中发现了另一个新物种, 并将其命名为Heterosigma minor[6]。
2 赤潮异弯藻的鉴定和监测
鉴于赤潮异弯藻藻华的危害性, 建立该物种准确、快速的鉴定方法对其藻华监测非常重要。传统的浮游植物鉴定, 主要利用普通光学显微镜对固定的藻细胞进行形态学观察[10], 然而常规浮游植物样品保存方法并不适用于赤潮异弯藻, 主要由于该物种形态多变, 且没有细胞壁, 在鲁哥氏等固定液中极易破碎。近年来, 分子生物学技术的发展为赤潮藻类的监测提供了新的思路[11]。相比于传统的显微镜观察, 分子生物学检测方法具有简单、准确、特异性高等特点, 且一些方法可用于大规模样品检测。目前有关赤潮异弯藻的分子检测方法均基于核酸特征建立, 即分子标记选择、引物或探针设计、物种检测或计数。在分子标记选择方面尽管有多种选择, 但核糖体RNA 标记仍是诸多研究的首选, 主要是由于该段序列由保守区和变异区镶嵌组成, 有利于设计不同分类水平的引物或探针。Tyrrell 等曾尝试将荧光原位杂交技术(Fluorescent in situ hybridization, FISH)用于赤潮异弯藻的检测, 但由于该物种细胞形态多变、不易固定等因素影响了该方法的建立[12]; 尽管Chen 等表明FISH 方法可对样品中70%~80%的赤潮异弯藻进行检测[13], 但笔者认为该方法操作繁琐、耗时, 不适宜赤潮异弯藻的环境样品监测; 三明治杂交(Sandwich hybridization assay, SHA)技术和实时荧光定量PCR 技术(Quantitative real-time PCR, qPCR)无需细胞固定和显微镜观察, 在一定程度上克服了FISH 方法的诸多缺点[14]。Tyrrell 等首次将SHA 方法应用到赤潮异弯藻的监测, 尽管SHA 在建立之初无法对样品中的细胞进行准确计数, 但可实现对该物种的藻华预警, 取得了良好的实际应用效果[12,15]。qPCR 是基于核酸提取的检测技术, Coyne 等利用Taqman 荧光探针法, 建立了赤潮异弯藻标准检测方法, 为了优化检测过程, 作者通过在DNA 裂解液中添加标准DNA, 以去除核酸提取、PCR 反应等过程带来的实验误差, 其结果表明该方法的检测极限为3.5 拷贝每反应, 且适用于环境样品检测[16]。为了比较SHA 和qPCR 检测方法在不同生理状态、不同培养条件和不同样品保存方法等情况下的细胞计数的准确性, 一系列比较实验陆续被展开, 为之后分子检测方法的选择提供了参考依据[17]。近年来, 环介导等温扩增技术(Loop-mediated isothermal amplification,LAMP)和超支化滚环扩增技术(Hyperbranched rolling circle amplification, HRCA)因其检测灵敏度高,也被应用于赤潮藻类检测, Zhang 等分别利用LAMP和HRCA 方法对赤潮异弯藻进行比较检测分析, 研究结果表明这两种方法均优于传统PCR, 且反应结束后可通过直接添加核酸荧光染料实现检测结果可视化[18]。
3 赤潮异弯藻藻华引起鱼类死亡的机制研究
诸多研究表明赤潮异弯藻藻华可造成养殖鱼类大面积死亡, 但其引起鱼类死亡的机制却一直存在争议[19-20]。目前为止, 关于赤潮异弯藻引起鱼类死亡的机制主要有三种: (1) 赤潮异弯藻产生超氧化物(Reactive oxygen species, ROS), 如超氧自由基、羟基自由基、过氧化氢等, 引起鱼类鳃损伤, 导致窒息死亡[21]; (2) 产生类似于裸藻毒素的有机神经毒素,引起鱼类心脏或者鳃部致病[22]; (3) 藻体分泌的粘液覆盖在鱼的鳃上, 导致鳃部病变, 引起鱼类窒息而死[23]。然而, 上述三种解释都尚存缺陷: (1) 尽管目前很多研究认为赤潮异弯藻的鱼毒性由该物种释放的ROS 引起, 但培养条件下该物种产生的过氧化氢浓度远未达引起脊椎动物细胞系和无脊椎动物死亡的浓度[20,24]。(2) 目前只在赤潮异弯藻中鉴定到裸藻毒素类似的有机物, 而不是通过毒素的作用机理来解释该藻的鱼毒性[22]; (3) 尽管鱼的鳃部黏液会引起窒息, 但无法确定黏液来自藻体还是鱼类, 且Chang 等认为致病鱼类鳃部的黏液量不至于引起鱼类鳃部严重病变[23]。此外, 有研究表明鱼类鳃部的黏液可能刺激赤潮异弯藻产生更多的ROS[25]。Twiner 等则认为赤潮异弯藻导致鱼类死亡原因可能是ROS 和有机毒素共同作用的结果, 首先藻细胞通过释放ROS 破坏鱼类细胞的免疫力, 然后利用产生的有机毒素改变细胞的离子平衡, 最后引起细胞凋亡[26-27]。综上, 赤潮异弯藻导致鱼类死亡的机制研究虽然已取得一定进展, 但尚存争议, 有待今后加强研究。
4 环境因子对赤潮异弯藻生长的影响
有害藻华的形成通常是赤潮生物本身(内因)和外界环境因子(外因)共同作用的结果。正常情况下,不同浮游植物通过相互协作或竞争, 构成相对稳定的生物群落结构, 当环境因子适宜特定生物生长时,则有利于该物种藻华形成。
营养盐(氮、磷等)是浮游植物赖以生存的基本元素, 其在海水中的浓度、存在形式等均会对赤潮生物的生长产生影响。在实验室培养条件下, Wang 等比较赤潮异弯藻、中肋骨条藻等5 种赤潮藻对9 种有机磷的利用情况, 结果表明赤潮异弯藻相对于中肋骨条藻可以利用更多形式的有机磷, 且在磷限制条件下(48 h), 赤潮异弯藻的碱性磷酸酶活性升高2.5倍[28]; Herndon 等研究发现赤潮异弯藻可有效利用三种常见氮源(硝酸盐、铵盐和尿素), 且在利用铵态氮时该物种生长率最高[29]。因此, 在营养盐匮乏的海域, 赤潮异弯藻对有机磷和多种氮源的利用能力可能有利于该物种获得种间竞争优势。
温度、光照以及盐度三个环境因子对浮游植物生长也起着重要作用。颜天等研究表明, 温度对赤潮异弯藻生长有显著影响, 盐度对该物种生长影响较小,且该物种的生长速率随着光照强度增强而增加[30]。Hennige 等研究结果显示在光强1 200 μmol·m–2·s–1条件下, 赤潮异弯藻仍未出现光抑制现象, 表明该物种拥有光保护机制[31]。除此之外, 有研究表明温度和光照在赤潮异弯藻孢囊萌发和赤潮起始阶段也起着重要作用[32-33]。
5 赤潮异弯藻基础生物学研究
Jeong 等基于赤潮藻的自身迁移行为、营养获取方式以及生物间的相互作用形式等, 将藻华生成机制(Generation Mechanisms, GM1-GM4)归纳为四个不同层次, GM1: 光合自养型; GM2: 具有昼夜垂直迁移行为; GM3: 混合营养型; GM4: 直接或间接抑制其它生物生长[34]。目前, 根据赤潮异弯藻的研究状况, 初步判断该物种可能属于上述分类的最高层次GM4(图1)。

图1 基于营养获取方式、昼夜迁移行为、生物间互作和群体结构示意赤潮异弯藻藻华发生机制(根据Jeong等图1 绘改[34])Fig. 1 Illustration of the mechanisms of bloom formation categorized based on nutrition, behaviors, and biological interactions of Heterosigma akashiwo and its community (Modified from Jeong et al. Fig. 1[34])
5.1 赤潮异弯藻昼夜垂直迁移行为
昼夜垂直迁移(Diurnal vertical migrations, DVMs)行为是一种广泛存在于浮游植物中的生物学现象,即黎明时藻细胞上浮至表层水体, 黄昏时下潜至底层。细胞通过向上迁移, 在水体表面聚集, 不仅加剧了藻华的形成, 而且垂直迁移还有利于浮游植物充分利用上层的光能和底层的营养盐。Shikata 等研究表明赤潮异弯藻等4 种鞭毛藻的DVM 行为主要受光周期调控, 且蓝光在保持和调节DVM 行为过程中起着重要作用[35]。Watanabe 等利用1.5 m 高的玻璃箱,在实验室条件下模拟表层低盐度、低磷酸盐, 底层高盐度、高磷酸盐垂直分层环境, 以期模拟水体混合较弱的夏季海区环境, 结果表明赤潮异弯藻夜间通过下移行为从底层水体获取磷, 并以多聚磷的形式进行储存, 而光照期间通过光合磷酸化利用夜间储存的磷[36]。因此, 相比较于硅藻等非鞭毛藻, 具有DVM行为的赤潮异弯藻在光能利用、营养盐摄取以及躲避浮游动物捕食方面具有一定的种间竞争优势, 这一特征在其藻华形成过程中可能起着重要作用。
5.2 赤潮异弯藻混合营养型生态特征研究
根据营养获取方式, 可将原生生物分为自养型、混合营养型和异养型。在研究之初, 赤潮异弯藻等针胞藻被归类为自养型浮游植物, 但Nygaard 等研究发现在磷限制条件下赤潮异弯藻可通过细菌吞噬(Bacterivory)作用获取磷[37], 其后Jeong 等利用荧光标记和扫描电镜技术再次确认了赤潮异弯藻属于混合营养型这个结论, 且该物种的捕食率也随着培养体系中被捕食者浓度增加而提高[38]。海洋浮游生物混合营养型是指这类生物既能进行光合作用, 又能进行营养摄取。因此, 相对于自养型浮游植物, 赤潮异弯藻混合营养型这一生物学特征不仅在一定程度上扩大了其营养来源范围, 更有利于该物种在营养限制条件下快速增殖, 取得种间竞争优势, 加剧藻华形成。
5.3 赤潮异弯藻化感作用抑制共发生微藻生长的研究
“化感作用”(allelopathy)一词最初由德国科学家Molisch 于1937 年提出。1996 年, 国际化感作用研究协会将化感作用定义为植物、藻类、细菌和真菌等产生的次级代谢产物(化感物质)对生物生长发育和农业体系产生的影响。在水生生态系统中, 化感作用作为浮游植物种间相互竞争的方式之一, 在赤潮物种取得竞争优势的过程中发挥着重要作用[39]。早在1966 年, 研究者就曾发现赤潮异弯藻和另一常见赤潮藻中肋骨条藻(Skeletonema costatum)之间存在种间竞争关系, 当赤潮异弯藻藻华发生时, 环境中中肋骨条藻的细胞丰度显著降低, 随着赤潮异弯藻藻华消亡, 中肋骨条藻的细胞密度才逐渐升高[40]。相对于中肋骨条藻, 赤潮异弯藻的细胞生长率较低,因此赤潮异弯藻藻华的发生不应归因于该物种的自身生长率优势, 这也表明赤潮异弯藻在赤潮发生时可能通过某种机制抑制环境中中肋骨条藻的生长。2007 年, Yamasaki 等基于共培养实验证明赤潮异弯藻与中肋骨条藻之间存在化感作用, 两年后该研究团队在实验室和环境样品均分离到赤潮异弯藻分泌的一种潜在的化感物质, 多糖-蛋白复合物(Polysaccharideprotein Complexes, APPCs), 且这种复合物可以专一性抑制中肋骨条藻的生长[41-42], 然而该物种化感作用抑制中肋骨条藻生长的机制目前却并不清楚。笔者利用宏转录组方法对赤潮异弯藻藻华发生时的环境样品进行了分析, 发现藻华发生时赤潮异弯藻可通过多种途径从环境中获取资源以促进自身快速增殖, 而中肋骨条藻的叶绿体核糖体蛋白、ATP 合成酶和光合作用相关基因表达水平却显著下调表达[43]。上述研究结果表明, 赤潮异弯藻可通过化感作用抑制中肋骨条藻的生长, 而且化感作用的机制可能也是复杂多样的。
6 赤潮异弯藻的分子生物学研究
分子生物学技术不仅在HAB 生物监测中广泛应用, 还促进了赤潮藻类的生物学、生态学研究, 如传统的生态学主要分析该物种在不同环境下的生理反应、与其他微生物的相生相克以及其行为学等方面,而分子生态学则从微观角度研究生态学现象的具体分子机制。
6.1 赤潮异弯藻DNA 分子标记的应用
为了研究全世界范围内赤潮异弯藻的分布情况,Ki 等基于DNA 测序技术, 对来自日本、韩国及北美等不同株系的多个分子标记(约26.7 kbp)进行测序,并结合NCBI 数据库中已知序列进行比对分析, 结果表明该研究所涉及的赤潮异弯藻株系基因型一致[44];Engesmo 等综合利用DNA 测序、扫描电镜和核糖体转录间隔区(ITS)二级结构分析, 在已鉴定的赤潮异弯藻中发现一个新种[6]。此外, 13 个多态性丰富的微卫星分子标记在赤潮异弯藻中也被开发, 并应用于该物种的遗传结构分析[45-46]。
叶绿体与线粒体是植物细胞内的重要细胞器,作为半自主性细胞器, 它们可以编码与自身功能相关的部分基因, 并参与光合作用、能量生成等重要生命活动。关于叶绿体和线粒体的起源, 目前“内共生”理论是普遍被大家认可的, 即叶绿体起源于内共生的蓝藻, 而线粒体起源于内共生的细菌, 因此分析叶绿体及线粒体基因组序列信息在揭示物种起源和分析不同物种之间的进化关系等方面具有重要的应用价值。Cattolico 等利用Fosmid 克隆技术成功的对西太平洋和西大西洋两株赤潮异弯藻的叶绿体基因组进行了测序分析, 结果表明该物种的叶绿体大小为160 kbp, 编码197 个基因, 且均存在22 kbp 的反向重复序列, 在基因编码区和间隔区均发现核苷酸多态性[47]; Masuda 等利用兼并引物成功的对针胞藻中的赤潮异弯藻和海洋卡盾藻(Chattonella marinavar.marina)线粒体基因组进行了测序分析, 结果表明赤潮异弯藻的线粒体全长38.7 kbp, 编码62 个已知功能和7 个未知功能基因, 且两个物种均含有重复基因组序列(Partial genome duplication)[48], 基于海洋卡盾藻的重复序列分析表明, 该重复区域序列变异较大, 可用于研究该物种的遗传多样性分析。
6.2 赤潮异弯藻分子生态学研究
分析不同代谢通路或典型基因对不同环境变化的响应是目前分子生态学研究的热点, 而赤潮异弯藻的相关分子调控机制研究却鲜有报道。Coyne 等发现赤潮异弯藻可通过快速调控硝酸还原酶基因(Nitrate reductase, NR)表达应对环境中氮营养盐的变化, 在18~28℃范围内该基因的表达水平未见明显波动, 且在硝酸盐缺失和铵盐存在情况下, NR 基因仍可以持续表达, 该结果表明赤潮异弯藻可通过快速调控NR 表达应对环境氮水平变化[49]; 为准确分析赤潮异弯藻功能基因在不同环境下的调控规律, 笔者在实验室条件对该物种在不同培养条件下的qPCR最适内参基因进行了系统筛选[50]。随后, 利用筛选的内参基因, 对赤潮异弯藻蓝光受体基因、减数分裂相关基因等功能基因进行了深入研究[51-52]。此外, 有研究发现赤潮异弯藻铵态氮转运蛋白的昼夜调控节律与细胞的昼夜垂直迁移行为可能协同促进了该物种对铵态氮的利用[53]。
随着高通量测序成本的降低、通量的提高, 多组学分析逐渐被应用于浮游植物生态学研究。2014 年海洋真核微生物转录组项目(MMETSP)公布了近680 个转录组数据, 其中针对赤潮异弯藻的4 个不同株系就设计了14 个处理组(表1), 涵盖不同营养盐、不同盐度等培养条件[54]。Haley 等转录组数据分析表明赤潮异弯藻在氮和磷限制条件下分别有31%和16%的转录本显著差异表达, 其中氮转运、代谢相关基因在氮限制条件下显著上调表达, 而磷转运蛋白和有机磷利用相关基因的表达水平在磷限制条件下显著上调[55]。此外, Hennon 等研究表明CO2水平的升高在生理和分子水平均会对赤潮异弯藻产生影响, 且生理反应和基因表达调控并非呈线性[56]。针对环境样品, 笔者利用18S rDNA 条形码和宏转录组(Metatranscritpome)方法对赤潮异弯藻藻华形成过程的浮游植物多样性和代表种的代谢通路进行分析,研究结果表明藻华形成过程中环境浮游植物群落结构明显变化, 且赤潮异弯藻可通过多种途径从环境中获得营养物质[43]。比较宏蛋白组(Metaproteomics)原位样品分析结果显示, 与甲藻东海原甲藻(Prorocentrum dongdaiense, 现更名P. shikokuense)相比, 赤潮异弯藻在浑浊度高、无机营养盐充足的水体中, 可通过上调一系列基因表达实现对磷、氮、无机碳等利用, 进而加速藻华形成[57]。
综上所述, 尽管赤潮异弯藻分子生态学研究已取得一定进展, 但相对于其他赤潮藻类研究仍处于落后水平, 很多生理学现象及藻华形成机制未能从分子水平上得到阐述。
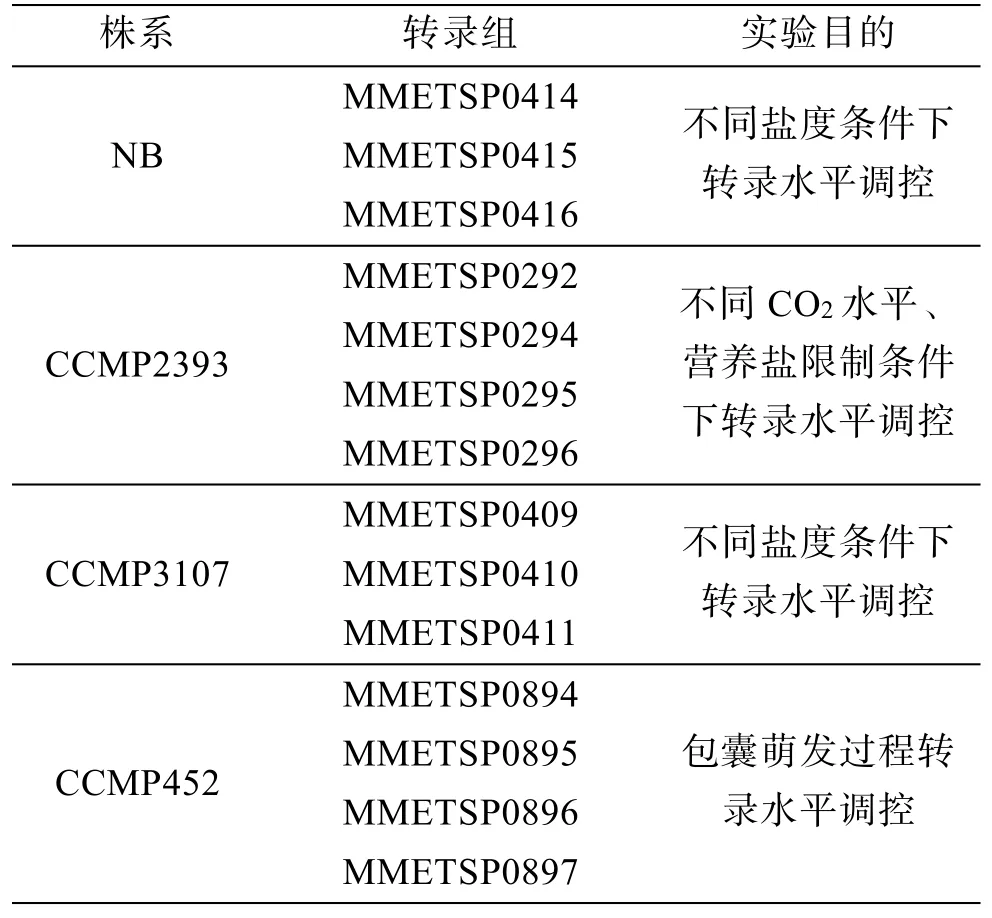
表1 MMETSP 项目中赤潮异弯藻转录组数据信息[54]Table. 1 Information of H. akashiwo transcriptomes in MMETSP[54]
7 全球气候变化下赤潮异弯藻藻华的发生趋势
许多研究表明全球气候变暖等现象导致浮游植物的群落结构逐渐趋于不稳定, 加剧了赤潮暴发[58]。温度对浮游植物个体的直接作用主要表现在影响细胞新陈代谢、孢子萌发、营养摄取和光合作用等生理过程[32-33,58]。不仅如此, 温度变化还可以改变生物间的相互作用以及营养物质输运等过程, 如温度升高不利于底层营养盐的上行输入, 进而增强了具有迁移行为的浮游植物竞争优势[58]。2018 年10 月政府间气候变化专门委员会(IPCC)在韩国仁川发布最新报告称, 与工业化前相比, 目前全球平均气温已升高了约1.0℃。研究者预测, 在不进行调控的情况下, 到2030—2052 年全球平均气温将升高约1.5℃[59], 因此海水升温对浮游植物群落结构的影响在未来几十年有可能继续加剧。此外, CO2作为光合作用的原料, 其水平高低在一定程度上也影响着浮游植物的群落结构。Fu 等研究结果显示相对于微小原甲藻(Prorocentrum minimum), 温度和CO2水平升高均能刺激赤潮异弯藻生长, 表明在全球气候变化的大背景下, 赤潮异弯藻具有一定的种间竞争优势[5]。此外,研究表明随着CO2水平升高赤潮异弯藻的向下游泳行为将增强, 且该物种运动相关基因的表达水平也显著上调, 进而影响赤潮异弯藻的藻华形成[56,60]。尽管上述研究从宏观水平说明了温度和CO2变化对赤潮异弯藻的藻华形成会有一定影响, 但其影响机制研究仍有待深入, 且在未来研究中应加强原位观测、综合模拟等分析。
8 赤潮异弯藻藻华防治方法研究现状
有害藻华频发对海洋生态系统和人类健康持续造成威胁, 因此对其治理和减灾是目前亟待解决的生态学问题。目前, 有害藻华防治主要从物理、化学、生物等方面展开, 其中改性黏土在治理赤潮和褐潮方面已取得重大突破。Sengco 等研究表明, 0.5 g/L的IMC-P2 磷黏土可以有效去除赤潮异弯藻, 且去除效果随着细胞浓度升高而增强[61]; 2006 年, 吴萍等研究发现0.03 g/L 的新型阳离子表面活性剂双烷基聚氧乙烯基三季铵盐改良后的黏土在24 h 内可以实现100%去除赤潮异弯藻, 且这种改性剂对养殖生物毒性较小[62]。化学法是指利用化学物质杀死赤潮藻,研究表明噻唑烷二酮衍生物(thiazolidinedione derivative) TD49 和灵菌红素(prodigiosin)在一定浓度下可有效杀死赤潮异弯藻[63-64]。相对于物理和化学方法,生物控藻是一种最为环保的方法, Kim 等研究表明溶藻菌在赤潮异弯藻凋亡期起着重要作用[65], 同样Zhang 等研究中使用的灵菌红素就是从赤潮异弯藻溶藻菌Hahellasp. KA22 分泌物中筛选得到的一种杀藻化合物[64]。综上, 目前已报道的赤潮异弯藻防治方法大多停留在实验室研究阶段, 其中改性黏土在治理赤潮异弯藻藻华治理方面可能有一定的应用前景。
9 总结与展望
近年来, 在全球气候变化和人类活动的双重作用下, 有害藻华造成的生态灾害日趋严重。赤潮异弯藻作为典型的赤潮生物, 其生物学和生态学研究一直备受关注。尽管本文从不同角度对赤潮异弯藻的研究进展进行了综述, 但该物种仍有多个问题待进一步解决: (1) 早期一些研究由于受实验方法和条件限制,赤潮异弯藻鱼毒性机制研究仅从宏观水平对生物学现象进行了描述。利用生态组学从分子水平深入研究毒性来源和作用机制有待进一步展开; (2) 针对赤潮异弯藻化感作用抑制环境中中肋骨条藻等浮游植物生长机制的研究不够深入, 如综合转录组、宏转录组、代谢组等多组学手段, 并结合传统生理学方法开展研究, 将有利于我们从不同水平全面阐释化感作用的机制; (3) 目前有关赤潮异弯藻的研究主要在实验室模拟条件下展开, 且多数研究仅聚焦于单因子影响分析,不能全面反映藻华发生时的实际情况, 因此在后期研究中应加强原位实验; (4) 尽管目前已针对赤潮异弯藻建立多个分子检测方法, 但大多局限于实验室分析,无法实现原位观测, 因此应推进该物种的藻华监测预警平台建立; (5) 有关气候变化对浮游植物群落结构以及赤潮异弯藻藻华发生趋势影响研究, 大多是基于单一物种模拟培养, 不能全面反应不同藻类之间的竞争关系, 因此应加强实验设计和长时间的原位调查,并辅以多组学研究阐明内在机制。综上, 从微观水平对宏观现象进行阐释将有助于我们认识赤潮异弯藻,有益于解析其藻华形成机制, 也将为该物种的藻华防控提供基础数据。
