弧唇裂腹鱼生物学特征研究
2021-03-02张建禄黄吉芹方程许涛清王开锋
张建禄 黄吉芹 方程 许涛清 王开锋


摘 要:2017年9月,在西藏察隅县察隅河采集10尾弧唇裂腹鱼(Schizothorax curvilabiatus),其体长范围为136~230 mm,体质量范围为25.7~144.2 g,研究了其部分生物学特征,首次报道弧唇裂腹鱼3项生物学指标:肠体比(RGL)为2.82±0.54、肥满度(CF)为(1.16±0.13)、脏体比(VSI)为(13.55±4.70)%。
关键词:弧唇裂腹鱼(Schizothorax curvilabiatus);生物学特征;西藏
中图分类号:S937. 4
弧唇裂腹鱼(Schizothorax curvilabiatus)隶属鲤科(Cyprinidae),裂腹鱼亚科(Schizothoracinae),裂腹鱼属(Schizothorax),别名弧唇弓鱼Racma curilabiata Wu et tsao(武云飞和曹文宣)、吉啊(门巴族语)[1]。分布于西藏雅鲁藏布江下游干支流及察隅河、丹巴曲,是我国西藏特有鱼类,世界自然保护联盟把弧唇裂腹鱼定义为数据缺乏类。目前可供参考文献仅两处,推测主要原因是分布局限、资源稀少且采样难度大。武云飞和吴翠珍(1992)[1]在《青藏高原鱼类》首次描述了其部分生物学特征。马海鑫(2019)[2]基于SLAF-seq技术开发弧唇裂腹鱼SNP位点并分析其群体遗传学特征。青藏高原属典型的高原地理气候,春季太阳辐射热能弱,水温回升慢,秋季水温下降快,导致鱼类生长期短,越冬期长[3],且西藏高原鱼类繁殖力普遍较低[4],资源一旦破坏很难恢复,使得对其资源的保护显得尤其重要。本研究旨在研究弧唇裂腹鱼生物学特征,为这一物种积累生物学资料。
1 采样及数据分析
2017年9月18日,采样点位于西藏察隅县城西南约10 km处的察隅河(28.616257N; 97.393831E,H: 1 968 m)采集弧唇裂腹鱼样本10尾。采样工具:手撒网。现场测量体长和体质量,体长测量精确到 0.1 mm,体质量称量精确到0.1 g。样品鱼用75%酒精浸泡后带回实验室。
统计所有数据的平均值和标准差,并对体长(L)-体质量(W)关系式进行比较拟合以确定其关系式。生物学性状计算公式如下:
肥满度(condition factor,CF,g/cm3)=体质量/体长3
肠体比(relative gut length,RGL)=肠长/体长
脏体比(viscerosomatic index,VSI,%)=内脏总重/全鱼重×100
2 结果及讨论
2.1 特征描述
2.1.1 外在形态特征 根据弧唇裂腹鱼样本描述其特征:體稍侧扁,吻钝圆。口下位,横裂或弧形。下颌具有锐利且较厚的角质边缘。下唇完整,但中间有皮肤与颏部相连,后缘不能完全游离,表面乳突明显,唇后沟连续。须2对,约等长,吻须末端超过眼前缘,额须末端后伸可达前鳃盖骨前缘。背鳍末根不分枝,鳍条粗硬,后缘具有数枚锯齿,背鳍起点至吻端的距离约等于其至尾鳍基部的距离;腹鳍起点对应于背鳍末根不分支鳍条或者第1根分支鳍条。除鳃颊部小区裸露无鳞外,体表几乎全被细鳞,侧线完全。背侧青灰色,头背部具多数黑点或星状小斑,腹部银白色,各鳍橘黄色。见图1。
2.1.2 可数和可量性状 第一鳃弓外鳃耙22~28,侧线鳞99~116,侧线上鳞19~26,侧线下鳞19~23,见表1。
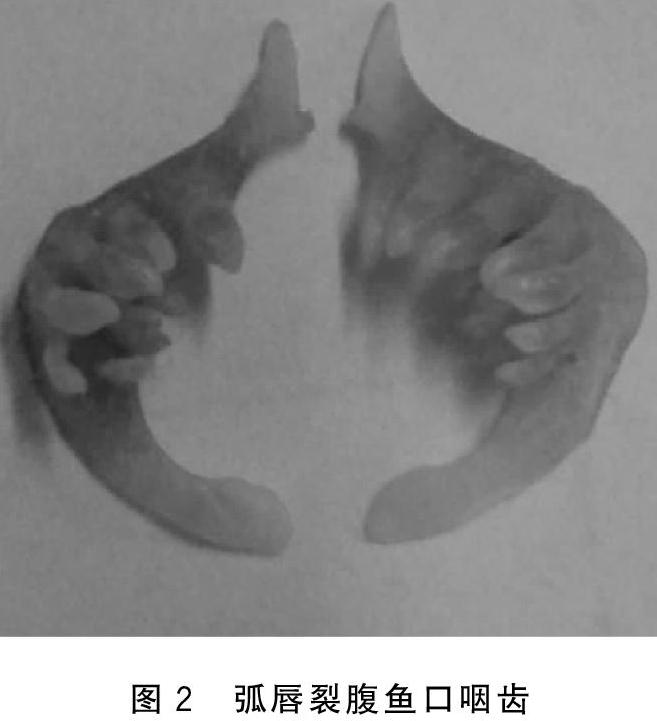
下咽齿细圆,顶端钝尖而略钩曲,咽齿3行,2·3·5/5·3·2,见图2。
10尾弧唇裂腹鱼体长范围136~230 mm,体质量范围25.7~144.2 g,RGL为2.82±0.54,CF为1.16±0.13,VSI为0.13±0.05,见表2。
分析表明,弧唇裂腹鱼体长为体高的4.43~4.89(4.69±0.15)倍;体长为头长的4.56~5.00(4.74±0.15)倍;体长为尾柄长的4.42~5.91(5.02±0.39)倍;尾柄长为尾柄高的1.44~1.79(1.62±0.10)倍。头长为吻长的2.82~4.83(3.32±0.58)倍;头长为眼径的5.17~7.23(5.87±0.70)倍;眼间距为眼径的2.17(2.28±0.42)倍。
2.2 弧唇裂腹鱼体长-体质量关系
鱼类体长-体质量关系在鱼类生物学研究中应用十分广泛,是鱼类种群研究中必不可少的内容,可以反映鱼类个体生长情况并判断种群变动方式[5-7]。鱼类的体长-体质量关系通常采用幂函数(W=aLb)进行回归分析,黄真理等(1999)[8]论证了此幂函数的意义。式中幂指数b反映鱼类生长类型的特征值,b=3为等速增长,说明鱼类体长和体质量的增加速度相等;b>3说明鱼类体质量增长速度快,b<3说明鱼类体长增长速度快。大多数动物的体质量并不是严格随体长匀速增长,即幂指数b接近3但并不等于3[9]。
本研究中弧唇裂腹鱼体长-体质量关系式为:
W=2×10-5 L2.938
式中:幂指数b=2.938,见图3。说明弧唇裂腹鱼体长增长速度大于体质量增长速度,但b值接近3,表明其体型已接近等速生长[10]。由于本研究仅采集到10尾弧唇裂腹鱼,样本量太小,可能偏差较大,因此本方程式仅供参考。已有裂腹鱼类体长-体质量关系的报道中,其幂指数b值分别为:异齿裂腹鱼b=2.828[11]和b= 2.940[12];拉萨裂腹鱼b=2.697 9[13],拉萨裂腹鱼性别未辨群体b= 2.800、雌性群体b= 2.984、雄性群体b=2.999[14];四川裂腹鱼b= 2.9511[15];宽口裂腹鱼b= 2.952,其雌性b= 3.214,雄性b=2.844[16];齐口裂腹鱼(S. prenanti)b=2.9678[17]。2008-2009年采集巨须裂腹鱼(S. macropogon)样本,雌性幂指数b= 3.104,雄性b=3.072;2012年采集巨须裂腹鱼样本,雌性b=2.937,雄性b=2.867[18]。
由此可见,幼鱼和成鱼之间、雌性和雄性之间、甚至同一物种不同时间段的幂指数b值不尽相同。以上多数裂腹鱼类生长关系的幂指数b<3,包括本研究中弧唇裂腹鱼b值。这可能预示着大多数裂腹鱼类生长模式为体长增长速度快于体质量增长速度。
2.3 弧唇裂腹鱼生物学指标
本研究首次报道了野生弧唇裂腹鱼的部分消化系统生物学指标,包括CF、RGL和VSI,这些指标在鱼类营养与饲料科学上有重要意义。消化系统的构造和食物质量密切相关[19]。当食物质量在时间或空间上发生变化时,动物的消化器官随之会产生相应的可塑性变化[20-21]。
2.3.1 肥满度(CF) CF也称丰满度,在生物学研究中应用比较广泛,鱼类学中用来反映鱼类生长的营养状况以及鱼群饵料保障的丰欠。CF受外部条件(如气候、饵料)及鱼类自身年龄等因素影响,也与性腺的发育有关[16]。可用来衡量动物体身体状况的特征值[22],也能反映鱼类个体生理及营养状况[6,23]。戴强等(2006)[23]和Jones等(1999)[24]认为相同体长下,CF值越高,表明其生理和营养状况越好。有研究报道了异齿裂腹鱼种群的平均丰满度,依次为幼鱼1.602 5,雌性1.442 9,雄性1.408 3,总体上幼鱼>雌鱼>雄鱼[12](马宝珊,2011)。拉萨裂腹鱼种群的平均丰满度依次为:幼鱼1.377 4、雌性1.243 5、雄性1.241 8,总体上也是幼鱼>雌鱼>雄鱼(周贤君,2014)[14]。
本研究首次报道野生弧唇裂腹鱼CF值1.16±0.13(见表2),为以后相关研究和该鱼种的开发利用提供参考资料。
2.3.2 肠体比(RGL) 肠道是鱼类吸收营养物质的重要场所,RGL是一个重要形态学指标,也是鱼类学中使用最广泛和实用的消化道参数之一[25]。RGL的大小与鱼类食性密切相关,其规律为:碎屑食性>植食性>杂食性>肉食性鱼类[26]。肉食性鱼类肠长仅为体长的1/3~3/4,草食性鱼类肠长甚至达体长的15倍(童裳亮,1988)[20]。影响RGL的主要因素是鱼的种类和食性,其他因素如饲料和生长环境,对其影响不大[27]。作为参照,杂食性的鲤(Cyprinus carpio)RGL为1.5左右[28-29],本研究表明弧唇裂腹鱼平均RGL为2.82(见表2),可见其食性比鲤更偏植物性。
裂腹鱼类食性研究方面的报道不多,已报道的裂腹鱼类的食性主要分为三大类:一是以藻类为主食即偏植物性的裂腹鱼,如中华裂腹鱼(S. sinensis)[30]、昆明裂腹鱼(S.grahami)[31]、拉萨裸裂尻鱼(Schizopygopsis younghusbandi)[32]、异齿裂腹鱼(S. oconori)[12]等;二是以无脊椎动物为主食即偏动物性的裂腹鱼,如四川裂腹鱼(S.kozlovi)[31]、拉萨裂腹鱼(S. waltoni)和巨须裂腹鱼[32]等;三是以鱼类为食即肉食性的的裂腹鱼,如新疆扁吻鱼(Aspiorhynchus laticeps)[33]和尖裸鲤(Oxygymnocypris stewarti)[34]。季强(2008)[32]报道了6种裂腹鱼类的RGL,分别为:拉萨裂腹鱼1.81~2.07,异齿裂腹鱼6.76~9.24,尖裸鲤0.75~0.89,拉萨裸裂尻鱼3.04~4.06,双须叶须鱼(Ptychobarbus dipogon)1.13~1.68,巨须裂腹鱼2.20~2.46。周贤君(2014)[14]报道拉萨裂腹鱼RGL为1.35~2.73,平均为2.02,并指出其食性为偏动物性,兼食着生藻类。由此可见,弧唇裂腹鱼RGL略大于拉萨裂腹鱼和巨须裂腹鱼,略小于拉萨裸裂尻鱼,这预示着弧唇裂腹鱼的食性比偏动物性的拉萨裂腹鱼更偏植物性,比偏植物性的拉萨裸裂尻鱼更偏动物性。
同一种鱼类的RGL随生境不同也会产生较大的差异,如敞水区生活的河鲈(Perca fluviatilis)比沿岸带生活的具有更大的RGL[35-36],因此,也可以通过比较不同河流弧唇裂腹鱼的RGL来评估不同河流中饵料生物的丰富度。有研究发现,弧唇裂腹鱼墨脱群体的遗传多样性高于易贡湖群体,作者推测可能主要是由于两群体所在生境中饵料丰度存在差距,导致易贡湖弧唇裂腹鱼群体的遗传多样性低于墨脱群体[2]。该研究未报道两个弧唇裂腹鱼种群的RGL。我们认为,如果能对这两个群体的RGL开展研究,可验证上述推测。
2.3.3 脏体比(VSI) VSI反映鱼体内脂肪积累量的多少[37],VSI主要受鱼类的种类及摄食情况等因素的影响,在鱼类营养与饲料科学研究中,VSI常用来反映某营养物质对养殖鱼类脂质代谢的影响。向枭等[38]认为,鱼类的VSI会随着鱼体的生长而降低。
目前鲜有关于裂腹鱼属鱼类VSI的报道,尤其野生裂腹鱼类的VSI数据还未见报道。本文作者曾报道了人工养殖齐口裂腹鱼VSI为10.56±1.87[39]。向枭等[38]报道的人工养殖齐口裂腹鱼幼鱼VSI在9.46~11.13之间,郑宗林等[40]报道的人工养殖齐口裂腹鱼幼鱼VSI在8.08~9.10之间。均明显小于本研究中弧唇裂腹鱼VSI(13.55±4.70,见表2)。目前,弧唇裂腹鱼还未开展人工驯养及相关实验研究,本研究数据为后期相关研究甚至人工养殖提供参考。
3 弧唇裂腹鱼资源的保护建议
武云飞和吴翠珍(1992)[1]認为弧唇裂腹鱼是产区(察隅、丹巴曲)的主要经济鱼类,但马海鑫等(2019)[2]基于调查的基础上,认为弧唇裂腹鱼自然资源量可能已下降到前所未有的程度,需进一步科学调查并开展保护。由于宗教信仰的缘由,当地藏族人民极少捕食鱼类,发现有捕鱼者甚至会主动阻止、举报,因而西藏鱼类资源受到良好保护。但近年来随着气候、环境变化,经济发展以及人们生活水平的提高,尤其是大量外地人口的迁入,对水产品需求量急剧增加,藏区特产裂腹鱼类资源也受到一定威胁。为了弧唇裂腹鱼资源的保护和合理开发利用,我们参考现有鱼类保护的措施,提出一些建议:一是开展其种群生物生态学研究,掌握其繁殖时间、地点,及种群数量变化规律,为其保护提供技术支持,如划定保护地点,设置禁渔区和禁渔期;二是当地渔政管理部门严格限制总捕捞量,提高起捕年龄,防止幼鱼被过度捕捞;三是严禁地笼网、粘网等小网目渔具,防止对幼鱼的捕捞,严禁电鱼、炸鱼、毒鱼等高破坏性的捕捞方式;四是渔政部门建立有效的禁渔期和禁渔区,目前弧唇裂腹鱼的繁殖期并不清楚,有待进一步的科学研究,但参考西藏地区其它裂腹鱼类的繁殖期一般在每年2~5月;五是开展当地水文环境和弧唇裂腹鱼种群的监测,为研究气候变化对其影响积累资料。
参考文献:
[1] 武云飞,吴翠珍.青藏高原鱼类.[M].成都:四川科学技术出版社,1992:384-385.
[2] 马海鑫.基于SLAF-seq技术的弧唇裂腹鱼SNP位点开发及群体遗传学分析[D].武汉:华中农业大学,2019.
[3] 龙华.青藏高原淡水渔业资源及其开发前景[J].中国水产,2004,29(5):79-80.
[4] 李红敬,谢从新.雅鲁藏布江鱼类资源与利用对策[J].江苏农业科学,2006,34(4):165-167.
[5] WOOTTON R J.The evolution of life histories:Theory and analysis[J].Reviews in Fish Biology & Fisheries,1993,3(4):384-385.
[6] 支兵杰,刘伟,石连玉.人工培育秋大麻哈幼鱼的生长与形态研究[J].水產学杂志,2009,22 (2) :49-52.
[7] ANDERSEN K H,JACOBSEN N S,FARNSWORTH K D.The theoretical foundations for size spectrum models of fish communities[J].Canadian Journal of Fisheries and Aquatic Science,2015,73(4):1-14.
[8] 黄真理,常剑波.鱼类体长与体重关系中的分形特征[J].水生生物学报,1999,23(4):330-336.
[9] 詹秉义.渔业资源评估[M].北京:中国农业出版社,1995:18-24.
[10] 赵天,刘建虎.长江江津江段中华沙鳅耳石及年龄生长的初步研究[J].淡水渔业,2008,38(5):46-50.
[11] 贺舟挺.西藏拉萨河异齿裂腹鱼年龄与生长的研究[D].武汉:华中农业大学,2005.
[12] 马宝珊.异齿裂腹鱼个体生物学和种群动态研究[D].武汉:华中农业大学,2011.
[13] 郝汉舟.拉萨裂腹鱼的年龄和生长研究[D].武汉:华中农业大学,2005.
[14] 周贤君.拉萨裂腹鱼个体生物学和种群动态研究[D].武汉:华中农业大学,2014.
[15] 李忠利,胡思玉,陈永祥,等.乌江上游四川裂腹鱼的年龄结构与生长特性[J].水生态学杂志,2015,36(2):75-80.
[16] 曹希全,李艳慧,魏杰,等.宽口裂腹鱼体长-体质量关系和肥满度[J].西北农业学报,2019,28(8):1380-1386.
[17] 段鹏翔.金沙江下游齐口裂腹鱼种群动态研究[D].长沙:湖南农业大学,2015.
[18] 刘洁雅.西藏巨须裂腹鱼个体生物学和种群动态研究[D].阿拉尔:塔里木大学,2016.
[19] SIBLY R.Strategies in digestion and defecation[M]//TOWNSEND C R,CALOW PEDS. Physiological Ecology: an Evolutionary Appraoch to Resource Use.Oxford: Blackwell Scientific Publications,1981:109-139.
[20] 童裳亮.鱼类生理学[M].北京:科学出版社,1988:253-255.
[21] HAMMOND K A. Seasonal changes in gut size of the wild prairie vole (Microtus ochrogaster) [J].Canadian Journal of Zoology,1993,71(4):820-827.
[22] 孙波,鲍毅新,张龙龙,等.千岛湖岛屿化对社鼠的肥满度之影响[J].动物学研究,2009,30(5):545-552.
[23] 戴强,戴建洪,李成,等.关于肥满度指数的讨论[J].应用与环境生物学报,2006,12(5):715-718.
[24] JONES R E,PETRELL R J,PAULY D.Using modified length-weight relationships to assess the condition of fish[J]. Aquacultural Engineering,1999,20(4):261-276.
[25] ELLIOTT J P,BELLWOOD D R.Alimentary tract morphology and diet in three coral reef fish families[J].Journal of Fish Biology,2003,63(6):1598-1609.
[26] Kramer D L,Bryant M J.Intestine length in the fishes of a tropical stream: 1. Ontogenetic allometry[J].Environmental Biology of Fishes,1995,42(2):115-127.
[27] 王爱民,刘波,黄金田,等.不同食性鱼的血红蛋白及其测定方法的比较研究[J].盐城工学院学报(自然科学版),2006,19(1):64-67.
[28] 封功能,杨文平,王爱民,等.饥饿胁迫对鲤形体、体成分及血液生理指标的影响[J].上海海洋大学学报,2011,20(6):814-819.
[29] 王文娟,汪水平,向梟,等.维生素E对鲤生长性能和肌肉品质的影响[J].中国粮油学报,2013,28(8):51-58.
[30] 冷永智,周祖清,黄德祥.中华裂腹鱼的生物学资料[J].动物学杂志,1984(6):45-47.
[31] 钱瑾,徐刚.乌江上游两种裂腹鱼食性的初步分析[J].毕节师专学报,1998(1):79.
[32] 季强.六种裂腹鱼类摄食消化器官形态学与食性的研究[D].武汉:华中农业大学,2008.
[33] 任波,任慕莲,郭焱,等.扁吻鱼的生物学研究[J].水产学杂志,2006,19(2):9-22.
[34] HUO B,XIE C X,MA B S,et al.Reproductive biology of Oxygymnocypris stewarti in the Yarlung Zangbo River in Tibet,China[J].Environmental Biology of Fishes,2013,96(4):481-493.
[35] SVANBCK R,EKLV P.Morphology dependent foraging efficiency in perch:a trade-off for ecological specialization?[J].Oikos Journal,2003,102(2):273-284.
[36] OLSSON J,QUEVEDO M,COLSON C,et al.Gut length plasticity in perch: into the bowels of resource polymorphisms[J].Biological Journal of the Linnean Society,2007,90(3):517-523.
[37] 付旭,崔前进,陈冰,等.饲料脂肪水平对淡黑镊丽鱼生长及色素蓄积的影响[J].大连海洋大学学报,2020,35(1):56-62.
[38] 向枭,曾本和,杨理想,等.高脂饲料中添加胆汁酸对齐口裂腹鱼幼鱼生长性能、肌肉营养成分含量、消化酶活性及血清生化指标的影响[J].动物营养学报,2019,31(11):5089-5099.
[39] 张建禄,王开锋,边坤,等.汉中地区齐口裂腹鱼生物学研究[J].基因组学与应用生物学,2018,37(12):5263-5268.
[40] 郑宗林,曾本和,向枭,等.饲料中胆汁酸添加水平对齐口裂腹鱼幼鱼生长性能、形体指标及体成分的影响[J].动物营养学报,2016,28(8):2423-2430.
