不同生育期对‘118’饲草谷子青贮品质的影响
2021-03-02赵牧其尔王志军贾玉山张永虎格根图
赵牧其尔,王志军,贾玉山,张永虎,温 蕊,包 健,格根图
(1.内蒙古农业大学草原与资源环境学院 / 农业部饲草栽培、加工与高效利用重点实验室 / 草地资源教育部重点实验室,内蒙古 呼和浩特 010019;2.内蒙古自治区农牧业科学院, 内蒙古 呼和浩特 010031)
谷子(Setaria italica)为禾本科(Gramineae)狗尾草属(Seataria)一年生草本植物,是重要的粮食作物和饲料作物[1]。近年来,随着谷子种植面积的扩大,其产物除了作为人类的食物外,还可以应用于畜禽养殖。谷子(或小米)除了作为猪鸡的能量饲料和牛羊的精料补充料外,谷子生物体产草量比例更大的植株部分还可作为草食动物的饲草使用。谷子的茎秆是牛、马、羊等家畜的优质粗饲料,其具有质地柔软、营养丰富、适口性好、和消化率高等特点,同时也是我国历史上重要的军草[2-3]。
优质饲草产品是连接草产业与畜牧业的纽带,草产业的健康健全发展可弥补国内畜牧业发展快速而草业发展缓慢带来的草、畜链条脱节问题,从而更好地保障和促进草牧业的健康发展。但我国草产品分布不均、饲草资源开发不够、秸秆利用率低、企业产能不足等问题导致国内草畜供需失衡,内循环动力严重不足。近年来,饲草谷子在我国农牧结合区和牧区发展迅速,使得饲草谷子的研究成为了热点[4-5]。有研究表明,在抽穗期至开花期刈割的新鲜谷子中,粗蛋白质含量较高,可直接青饲,也可调制干草、青贮饲料或加工成颗粒饲料等草产品[6]。李林等[7]探索不同切碎长度和发酵方式对全株饲用青贮谷子质量的影响,结果表明不同切碎长度和发酵方式对于全株饲用谷子青贮质量均有显著影响,降低切碎长度和使用复合菌剂发酵均有助于促进纤维分解,减少丁酸产量,改善青贮感官评分。薄玉琨等[8]探索全株饲用谷子青贮对育肥期阳原驴生长性能的影响及饲喂后肉驴血清抗氧化能力变化,对照组肉驴以花生(Arachis hypogaea)秧干草 + 玉米(Zea mays)秸秆饲喂,试验组以全株饲用谷子青贮按干物质(dry matter, DM)比例1 ∶ 1 替代花生秧干草,试验结果表明育肥期阳原驴日粮中添加全株饲用谷子青贮能够增加肉驴的采食量,提高育肥期日增重,增强肉驴机体抗氧化能力。刘婷婷[9]对‘张杂谷’饲草开展了谷草氨化、全株谷子青贮和收获籽实后谷秸黄贮3 种加工调制技术探究,通过营养成分含量变化和体外产气法评定其对‘张杂谷’饲草营养价值的影响,以确立‘张杂谷’饲草适宜的加工调制技术。结果表明,‘张杂谷’谷草氨化的适宜尿素用量为5%,水用量为50% ;‘张杂谷’全株青贮以添加4% 的糖蜜为宜;‘张杂谷’谷草黄贮以添加1‰ 菌剂效果最佳。确立了‘张杂谷’饲草的适宜氨化、青贮和黄贮加工调制技术。相比较成熟的禾本科和豆科牧草的青贮技术,饲草谷子的适时收获期、青贮技术和青贮品质等问题均有待研究。基于此,本研究分析饲草谷子不同生育期的营养价值,探究其青贮过程中的营养成分、发酵品质以及微生物数量的变化规律,旨在为调制优质饲草谷子青贮饲料、科学利用饲草谷子提供数据参考,以期为饲草谷子的加工利用技术提供理论依据。
1 材料与方法
1.1 试验地概况
试验地点位于内蒙古乌兰察布四子王旗吉生太镇老圈滩村,地处 111°21′-111°34′ E,41°34′-41°54′ N。平均海拔1 400 m,属丘陵地区。气候属温带大陆性气候。历年平均降水量在110~350 mm。
1.2 试验材料
供试品种‘118’于2018 年5 月17 日种植,由内蒙古自治区农业科学院提供,播种面积为0.067 hm2,播深3 cm,播量为2 kg·hm-2。‘118’品种是内蒙古清水河县的农家品种,发芽率为80%。在呼和浩特市生育期为127 d 左右,苗色绿色,植株半上冲,株高平均173.70 cm,抗倒伏能力强。‘118’品种灌浆期收获时间为2018 年8 月26 日,乳熟期为9 月4 日,蜡熟期为9 月19 日,上述生育期的产量分别为30.783、37.770 和25.747 t·hm-2(表1)。

表1 谷子收获时间及产量Table 1 Harvest time and yield of millet
1.3 试验设计
在供试品种‘118’进入灌浆期、乳熟期、蜡熟期时,随机选取3 个1 m2(1 m × 1 m)样方进行刈割收获,利用铡草机将谷子切短至2 cm,混合均匀后装入聚乙烯袋,每袋装入300 g,用真空封口机抽真空并密封,室温条件下贮存。在厌氧发酵60 d 后开袋,测定其营养成分(干物质、有机质、粗蛋白质、中性洗涤纤维、酸性洗涤纤维、可溶性糖、粗脂肪)、发酵品质(pH、乳酸、乙酸、丙酸、丁酸)和微生物数量,从而确定调制谷子青贮饲料的最适生育期。
1.4 测定指标
样品制备:将采集的样品进行鲜重的测定,然后置于烘箱中,先在105 ℃杀青30 min,再将烘箱温度设定为65 ℃烘干48 h 后,进行其风干重的测定。用粉碎机分别粉碎制备0.850 和0.425 mm 分析样品,用于营养成分的测定。采用烘干法[10]进行干物质含量的测定;采用GB6439-92 燃烧法测定有机质(organic matter, OM)的含量[10];利用全自动杜马斯定氮仪进行测定粗蛋白质(crude protein, CP)含量[11];采用 GB 6433-94 索氏脂肪提取法测定粗脂肪(ether extract, EE)含 量[12];利 用ANKOM A220 全 自动纤维仪分析品的中性洗涤纤维(neutral detergent fiber, NDF) 与酸性洗涤纤维(acid detergent fiber,ADF)含量[13];利用蒽酮—硫酸比色法进行可溶性碳水化合物(water soluble carbohydrate, WSC)含量的测定[14]。
将青贮饲料开袋后,称取10 g 左右的样品,利用剪刀将其剪碎,再向袋内加入90 mL 蒸馏水,静置1 min,然后放在拍打仪中拍打2 min。拍打完成后,首先利用4 层纱布进行初滤,将残渣与液体分离,然后利用滤纸进行过滤,制得青贮浸提液。用雷磁PHS-3C 精密pH 计(北京东南仪诚实验室设备有限公司)测定浸提液的pH[15];将青贮浸提液倒至离心管中,然后将其放置在离心机中离心15 min (转速为4 000 r·min-1),利用0.22 μm 的微孔滤膜对上清液进行过滤,备用进行有机酸的检测。所用设备为高效液相色谱仪(HPLC),其系统Waters 2695-2489(紫外),从而分析滤液中的有机酸的含量[16],有机酸分离采用的色谱柱:Rspak 系列,KC811 柱,8 mm ×300 mm;流动相:3 mmol·L-1优级纯高氯酸,流速1 mL·min-1,柱温40 ℃;检测波长210 nm,进样量5 μL。
取10 g 青贮样品置于青贮袋中,加入90 mL 无菌水,用拍打仪拍打2 min 使其均匀混合,用8.50 g· L-1无菌生理盐水稀释10-1、10-3、10-5。乳酸菌数量采用MRS (Difco)固体培养基,霉菌和酵母菌采用 PDA(Potato Dextrose Agar.Nissui Ltd.Tokyo.Japan)培养基,大肠杆菌采用BLB (Blue Light Broth,Nissui Ltd.Tokyo.Japan)培养基,一般好氧细菌采用营养琼脂(Nutrient Agar Nissui Ltd.Tokyo.Japan)培养基,乳酸菌30 ℃厌氧培养箱中培养48 h 后计数,酵母菌、霉菌、一般好氧细菌、大肠杆菌置于30 ℃恒温培养箱中培养48 h 后计数,所有微生物数据采用cfu· g-1表示。
1.5 青贮品质综合评价
采用模糊函数的隶属函数法对不同生育期的青贮品质进行综合性评价[17-19]。
隶属函数值计算公式:

反隶属函数值计算公式:

式中: µ(Xj)表示隶属函数值,Xj表示第j个指标测定值,Xmax和Xmin表示第j个指标的最大值和最小值,若其中某一指标与青贮品质负相关,则利用反隶属函数转换计算其抗性隶属函数值。

式中:Xj表示各生育期第j个指标平均值;Xij表示i生育期j指标的隶属函数值;Vj表示第j个指标标准差系数。

式中:Wj表示第j个指标的权重。
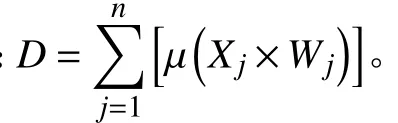
式中:D表示不同生育期的青贮品质综合评价值,D值越大表明品质越好。
1.6 数据处理与分析
采用Excel 2007 对数据进行预处理,应用SAS 9.0 软件对试验所得数据进行方差分析,并用 Duncan法对不同生育期营养指标的均值进行多重比较,数据以平均值 ± 标准差(Mean ± SD)表示。
2 结果与分析
2.1 青贮前后营养成分的变化
不同生育期饲草谷子各营养物质含量呈现出明显的规律变化(表2),DM 含量随着生育期的推进逐渐增加,到蜡熟期时,干物质含量在50%以上。随着籽粒成熟度的提高,CP 和EE 含量呈现降低趋势,而WSC 含量的变化则与之相反,随着生育期的推进而逐渐增加。NDF 含量在灌浆期达到最高,显著高于乳熟期和蜡熟期(P< 0.05),呈现先降低后增加的趋势。

表2 谷子青贮前后营养成分分析Table 2 Analysis of nutrient composition of millet before and after ensiling
青贮60 d 的CP 含量较原料有所降低,其中乳熟期和蜡熟期青贮前后的CP 含量有显著差异(P<0.05)。草谷子在调制青贮后,NDF 和ADF 含量均有所降低,其中蜡熟期青贮前后的NDF 存在显著差异(P< 0.05),从不同生育期青贮饲料中的NDF 和ADF 含量可知,乳熟期的NDF 含量显著低于灌浆期(P< 0.05)。乳熟期的WSC 含量高于灌浆期和蜡熟期,但不同生育期之间无显著差异(P> 0.05)。
2.2 不同生育期发酵品质的变化
青贮60 d 后,蜡熟期的pH 最高,其次为灌浆期、乳熟期,蜡熟期的pH 显著高于乳熟期(P< 0.05)(表3)。不同生育期的乳酸、乙酸、丙酸均无显著差异(P> 0.05),其中灌浆期的乳酸、丙酸含量最高。不同生育期的青贮饲料中均未检测到丁酸。

表3 谷子青贮饲料发酵品质分析Table 3 Analysis on fermentation quality of millet silage
2.3 不同生育期微生物的变化
原料和青贮60 d 的乳酸菌数量随着生育期的推进呈先增加后降低趋势(表4)。乳熟期原料附着的乳酸菌数量最多,与灌浆期和蜡熟期的乳酸菌数量存在显著差异(P< 0.05)。青贮后,乳酸菌的数量有所增加,不同生育期的乳酸菌数量无显著差异(P>0.05)。与原料相比,乳熟期和蜡熟期的乳酸菌数量显著增加(P< 0.05)。原料的一般好氧性细菌和大肠杆菌的数量随着生育期的推进逐渐增加,青贮60 d后均未检测到大肠杆菌。青贮前后,不同生育期的酵母菌和霉菌的数量无显著差异(P> 0.05)。厌氧发酵60 d 后,灌浆期、蜡熟期的霉菌和酵母菌的数量减少,但无显著差异(P> 0.05)。与原料相比,青贮饲料中的一般好氧性细菌的数量有所减少。

表4 青贮前后微生物数量的变化Table 4 Changes in microbial quantity before and after silage
2.4 隶属函数综合评价
利用隶属函数值法对饲用谷子不同生育期的青贮品质进行综合评价(表5)。不同生育期的青贮品质对比结果依次为乳熟期 > 蜡熟期 > 灌浆期。

表5 饲用谷子不同生育期的青贮品质综合评价Table 5 Comprehensive evaluation of silage quality of forage millet at different growth stages
3 讨论
干物质含量是青贮成败的关键。青贮饲料水分含量的高低决定着青贮发酵过程和青贮饲料的发酵品质且影响细菌总数和发酵速率[21]。由于谷子开花至成熟后水分流失,随生育期的延迟水分含量逐渐减少,干物质积累逐渐增多,导致蜡熟期干物质含量过高[22-23]。过高的干物质含量会抑制微生物的生长,减缓或者停止青贮进程,而过低的干物质含量会导致梭菌的大量生长,产生大量丁酸,不仅使青贮饲料营养成分损失还会产生臭气从而影响采食量。由于在青贮饲料早期发酵阶段存在一些产纤维素酶的微生物[24],青贮中的营养物质被其消耗产生纤维素酶导致酸性洗涤纤维下降较快且伴随有中性洗涤纤维的降低,下降到发酵中期基本稳定。随着籽粒成熟度的提高,‘118’品种的中性洗涤纤维和酸性洗涤纤维含量逐渐降低。研究表明, 随着植株的成熟,牧草中细胞壁成分增加, 纤维素类物质含量也会增加[25]。纤维素类物质含量先增加后减少,开花期之后,乳熟期、蜡熟期有更低的纤维类物质含量,这可能是谷子籽粒形成后,在灌浆过程中,籽粒中淀粉不断沉积使纤维素含量相对下降,这与Crovetto 等[26]研究的全株小麦的中性洗涤纤维和酸性洗涤纤维含量在抽穗期至乳熟期相对稳定, 在蜡熟期减少的结果相近。
Nordkvist 和Aman[27]的研究表明,牧草的生长时期对牧草积累的粗蛋白质含量有一定的影响,生长阶段与粗蛋白质含量两者有较高的关联性。‘118’品种的粗蛋白质含量随着生育期的延长逐渐降低,在青贮发酵过程中蛋白质有所降解,乳熟期的蛋白损失率比灌浆期和蜡熟期低。由于牧草自身的特性,不同品种的谷子其粗蛋白质、中性洗涤纤维、酸性洗涤纤维和可溶性糖含量也不相同。青贮发酵时微生物需要能量来源以保证自身生长,可溶性糖作为乳酸菌生长碳源摄取的主要底物,在青贮过程中被乳酸菌代谢消化,其含量可以间接反映乳酸菌生长情况。随着生育期的推进,供试品种的可溶性糖含量逐渐增加。青贮60 d 后,不同生育期的可溶性糖含量呈先增加后降低趋势,这与Wang 等[28]以马鞭草(Verbena officinalis)作为青贮原料,厌氧发酵过程中可溶性糖含量有所降低的研究结果相同。这是因为在青贮饲料发酵过程中需要消耗可溶性糖,以便为乳酸菌提供良好的生长环境和营养[29],从而导致消耗量增加和乳酸产量增加。
青贮发酵主要在厌氧条件下进行,而厌氧环境有利于有机酸的产生,青贮发酵中主要以乳酸为主。发酵过程中乳酸的含量越高则青贮品质越好,因此在一定程度上可以通过增加青贮饲料的乳酸含量来提高青贮品质。乙酸主要来源于醋酸菌、异型乳酸菌和肠内细菌对糖类的分解发酵[30],‘118’品种乳熟期的乳酸含量高于乙酸,表明青贮饲料中同型发酵乳酸菌较多,能够更充分地利用营养成分,减少营养物质损失。
青贮发酵程度的增加伴随着酸性产物的蔓延,其他微生物活动受到抑制,乳酸菌逐渐抑制其他微生物的活动[31],从而占据主导地位,产生大量乳酸,‘118’品种不同生育期的乳酸菌数量均有所增加。酵母菌和霉菌是青贮饲料中主要存在的真菌,在发酵底物充足且pH 降低的条件下,乳酸菌快速繁殖后自身被抑制,进而导致乳熟期的酵母菌大量繁殖,而酵母菌通过分解糖和乳酸,造成干物质的损失和pH 上升,引起其他杂菌的生长。pH 是反映青贮饲料品质优劣的一个重要指标, 常规成功青贮要求的pH 应低于4.2[32]。‘118’饲草谷子3 个处理组的pH 较高,可能是由于青贮饲料中附着的酵母菌和霉菌较多,与乳酸菌竞争发酵底物,消耗了部分可溶性糖,无法产生大量的乳酸,抑制有害微生物的生长繁殖。张慧杰[33]指出,植物原料上的微生物数量很少,不良微生物占据绝大部分,植物表面上乳酸菌数量在105cfu·g-1以上,才能达到青贮标准。在不同种类的作物表面上附着的微生物种类和数量也有很大的差异。其中,乳酸菌的数量对饲料的青贮发酵品质和饲料的保存有重要的影响等[34]。‘118’饲草谷子单独青贮时,不同生育期的青贮品质存在差异,但由于原料附着的乳酸菌数量均不足105cfu·g-1,pH 偏高,乳酸含量较低,无法抑制青贮饲料中的有害微生物的生长繁殖,导致青贮饲料品质较低。青贮中使用青贮添加剂,可以有效抑制不良微生物的繁殖,增加乳酸菌的数量,提高青贮的发酵品质[35]。因此‘118’饲草谷子在调制青贮饲料时,可通过添加青贮添加剂抑制有害微生物的生长繁殖,减少营养物质的流失,提高其青贮品质。
4 结论
不同生育期处理的谷子青贮60 d 后,其青贮品质和微生物数量均发生了变化。乳熟期的pH 和中性洗涤纤维含量最低,且乳酸菌数量、乳酸、可溶性糖和粗脂肪含量最高,是‘118’饲草谷子调制青贮饲料的最适生育期。
