干旱胁迫对杜仲叶片结构特征的影响
2021-03-02杜红岩刘攀峰杜兰英
何 凤 杜红岩 刘攀峰 王 璐 庆 军 杜兰英*
(1. 中国林业科学研究院经济林研究开发中心,国家林业和草原局杜仲工程技术研究中心,郑州 450003;2. 南京林业大学,南京 210037)
干旱是造成经济损失的主要自然灾害之一,对全球农作物生产的危害相当于其他自然灾害之和[1]。植物可通过形态结构和生理生化功能的变化以适应干旱胁迫。叶片是植物进行光合作用和呼吸作用的主要器官,随环境变化往往表现出其外部形态及内部解剖结构的差异,因此研究叶片的生态解剖学特征可以揭示干旱胁迫对植物的影响[2]。气孔是与叶片的功能有着密切关系的生理器官结构,它既是与外界进行气体交换的门户,又是水汽蒸腾的通道,与抗旱性能密切相关,植物在干旱胁迫下可通过气孔调节来适应[3~4]。
目前,已有诸多关于植物在干旱胁迫下叶片解剖结构和气孔响应的研究。曹林青等[2]研究发现,油茶在中重度干旱程度下,成熟叶片出现叶片变薄,栅栏组织和栅海比变小,海绵组织厚度变化不显著,气孔面积、气孔大小、气孔开度减小,气孔密度先减小后增大等解剖结构的变化,这些变化是油茶叶片对干旱逆境的适应特征进而维持生存和生长。黄绢等[5]研究发现,干旱胁迫下转基因杨ABJ01 和非转基因杨9#叶片厚度、上表皮厚度、海绵组织厚度、下表皮厚度显著降低,9#栅栏组织厚度显著降低,而ABJ01 则显著升高,表明ABJ01 具有更发达的栅栏组织,与很多植物相反,转基因杨ABJ01 通过栅栏组织厚度的升高来适应环境的变化。董晓民等[6]研究发现,随着干旱胁迫时间的延长,2 个扁桃品种叶片厚度均明显减小,叶肉上下表皮细胞变小,细胞间隙变小,海绵组织厚度降低,栅海比值逐渐增大,这些叶片结构朝着耐旱的方向改变,以减少水分蒸发适应环境变化。庞杰等[7]研究发现,随着土壤含水量下降,沙芥幼苗叶片厚度、上表皮厚度、下表皮厚度呈下降趋势,海绵组织厚度呈先下降后上升的趋势,栅栏组织和栅栏组织/海绵组织呈先上升后下降的趋势。高彦萍等[8]研究发现,经水分胁迫,大豆叶片气孔密度增加,气孔开口大小和单位叶面积气孔相对面积减小,不同抗旱类型大豆气孔密度变化幅度大小不同。刘世鹏等[9]研究发现,随着干旱胁迫程度的增加,5 个品种枣树叶片气孔密度先升后降,长度和宽度之间呈极显著线性关系,气孔大小先减小后增大,密度与长度、宽度之间均表现为极显著的负线性关系。Chartzoulakis K 等[10]研究发现,在干旱胁迫下橄榄叶片的栅栏组织、海绵组织的细胞间隙明显降低,细胞密度明显升高,这些变化导致单位叶面积增加进而促进CO2的吸收,从而在气孔导度低的干旱条件下保持光合速率。刘球等[11]综述了干旱胁迫对植物叶片解剖结构影响,为高等植物抗旱能力研究提供参考,文中介绍了不同植物叶片解剖对干旱胁迫响应不同。因此植物适应干旱环境的对策是多种多样的,杜仲(Eucommia ulmoidesOliv.)属杜仲科(Eucommiaceae)植物,该科仅有1 属1 种,自古以取皮入药而著称,有关杜仲逆境胁迫的研究较少,且内容多集中在逆境生理指标[12~16]、光合特性等方面[17~19],关于干旱胁迫下杜仲叶片结构变化的研究较缺乏[20]。本研究选取2 年生‘华仲6 号’杜仲扦插苗为试验材料,干旱胁迫条件下观察并分析其叶片外部形态特征,叶片下表皮气孔特征、叶肉和叶脉显微结构,探究杜仲幼苗叶片结构在不同程度的干旱胁迫下的变化及规律,进一步了解其干旱适应机理。
1 材料与方法
1.1 试验材料
试验在国家林业和草原局泡桐研究开发中心原阳基地的大棚内进行。2018年12月将当年扦插生根的杜仲扦插苗‘华仲6号’幼苗移栽至高15 cm,入口直径15 cm 的营养钵中,钵底有4 个小孔以保持下层土壤通气性,每个营养钵中种植1 株,栽培土壤为基地内大田栽培土,土壤质地为壤土,全氮8.2 mg·L-1,全磷28.0 mg·L-1,全钾5.0 mg·L-1,全钙1 000.0 mg·L-1(全氮、全磷、全钾、全钙使用百灵达SKW500 仪器测定),每营养钵中土重约1.80 kg,田间持水量为40.11%,定植并浇透水放至大田中培养,保证土壤水分基本充足,确保苗木成活。
1.2 试验处理
于2019 年5 月1 日选取长势基本一致的营养钵苗(地径4.45±0.57 cm,苗高19.09±4.96 cm)共72 盆移至简易避雨棚中,防止雨水进入土壤水分控制区,每6 个营养钵苗放在1 个托盘里,托盘高5 cm,浇透水,托盘里盛一部分水,以保证每个营养钵的土壤含水量基本一致,之后每5 d浇透水1次。2019 年5 月16 日开始,对照(CK1、CK2、CK3)组每隔5 d 浇透水,干旱处理组采取自然干旱的方式进行,按停止浇水6 d(5 月22 日)、11 d(5 月27 日)、16 d(6月1日)作为水分干旱胁迫处理,3个时期分别编号为T1、T2、T3。各处理重复3次,每个重复4株,每次试验采集对照组和干旱处理组叶片、根系附近土壤。每个重复采集顶部第1~5片成熟叶片混匀备用。从基地气象站获取试验期间气象数据。
图1 为营养钵苗土壤质量含水量的变化和采样期间试验地气温变化。对照组幼苗土壤质量含水量为31.95%~34.29%,在干旱胁迫6 d、11 d、16 d 时,土壤质量含水量分别为24.54%、12.03%、2.60%。5月18日至22日和5月28日至6月1日的平均气温分别为22.32℃和22.37℃,这两个时间段的气温相似,5月23日至27日的气温最高,平均气温为27.58℃。
1.3 测定方法
1.3.1 土壤质量含水量的测定
采集扦插苗根部附近的土壤迅速放入铝盒中,每个重复挖取1 个扦插苗附近的土,3 次重复,带回实验室,采用烘干法测定,取其平均值。
1.3.2 叶片相对含水量的测定
取样后将叶片表面擦拭干净,立即称叶片的鲜质量(Wf),后将叶片浸入蒸馏水中24 h,取出用吸水纸吸干叶片表面水分,称饱和质量(Wt),最后将叶片置于105℃烘箱中杀青15 min,于80℃下烘干至恒量,称干质量(Wd),计算叶片相对含水量(RWC),每个处理3次重复,结果取平均值。
1.3.3 叶片细胞显微结构观测
每个重复选取1 片叶,3 次重复,取叶片中部连着中脉剪取1.5 cm×1.5 cm 的方块,置于FAA 固定液(70%乙醇90 mL+甲醛5 mL+乙酸5 mL)中固定,于4℃冰箱中保存,用于做常规石蜡切片,材料经FAA 固定24 h 以上,乙醇和二甲苯系列脱水透明,浸蜡,包埋。切片(切片厚度8 μm)用番红-固绿染色,中性树胶封片后制成永久装片,用显微镜观察并照相,每个处理观测6 个视野,用ImageJ 软件观察分析测定叶片栅栏组织、海绵组织、上表皮、下表皮、叶脉等结构参数,结果取平均值。
1.3.4 叶片下表皮气孔结构扫描电镜观测
每个重复选取1 片叶,3 次重复,取叶片中部主脉一侧剪取1 cm×1 cm 的方块,置于电镜固定液(2.5%的戊二醛)中固定,于4℃冰箱中保存备用做电镜扫描,每个处理在扫描电子显微镜下观察15个视野并拍照。从中选取1 张100 倍镜的视野统计气孔数目和闭合气孔的数目,计算气孔密度(个/mm2)和气孔闭合百分比;从中选取2 张400 倍镜视野用ImageJ 软件测量每个气孔的面积、长度、宽度,气孔长度为平行于气孔器的最长值,气孔宽度为垂直于气孔器的最宽值,求取2个视野下气孔面积、长度、宽度的平均值。每个处理3次重复,结果取平均值。
1.4 统计分析
利用Microsoft Excel 2016 和SPSS 21.0 软件对数据分析、显著性检验和相关性分析。
2 结果与分析
2.1 杜仲叶片解剖结构和下表皮气孔特征
‘华仲6号’叶片形态结构为长椭圆形,叶缘锯齿,叶尖尾尖,基部圆形,叶片嫩绿显光泽,与一般双子叶植物的叶相同(见图2A)。叶片解剖结构可分为表皮、叶肉、叶脉,为背腹型叶。上下表皮均由1层表皮细胞组成,排列紧密。上表皮细胞较厚,由类椭圆形细胞排成1层;下表皮较上表皮薄,细胞体积较小,类长方形。叶肉有明显的栅栏组织和海绵组织的分化,栅栏组织紧靠上表皮,排列紧密,细胞呈长圆柱形,内含类似小泡样结构;海绵组织靠近下表皮,细胞较大,近椭圆形,靠近栅栏组织的叶肉细胞排列较为紧密,靠近下表皮的叶肉细胞排列较为松散无序,胞间隙较大,便于气孔与叶肉之间的联系(见图2B)。
主脉由表皮、厚角组织、薄壁组织、韧皮部、形成层、木质部构成,为外韧型,略呈凹槽状,木质部导管径向排列紧密,韧皮部细胞较小呈密集无规则分布,形成层较明显。韧皮部的外侧有薄壁组织和厚角组织,共11~16 层细胞,厚角组织3~4层细胞,较为发达(见图2C)。
气孔器是指气孔与两个保卫细胞组成的复合体。植物体通过保卫细胞来调控气孔的开张度,从而调节气体的出入和水分蒸腾。从电镜扫描照片看(见图2D),下表皮向表面隆起,有大量气孔器分布,气孔器由两个肾形的保卫细胞组成,呈椭圆形,无副卫细胞,两个肾形细胞对称排列,气孔呈纺锤形。保卫细胞由几个普通的表皮细胞不规则地围绕,属于无规则型。下表皮具角质层,呈棱状突起往往沿着某一方向延伸。气孔器的两个保卫细胞表面光滑,在气孔器周围,常由4~5 条棱状突起角质层环绕,个别的气孔器外由光滑的突起包围。气孔与下表皮细胞处于相似的水平。
2.2 干旱胁迫下杜仲叶片相对含水量和表型特征变化
随着干旱胁迫时间的延长,对照组的叶片相对含水量维持在74.88%~76.59%,干旱处理组叶片相对含水量持续下降;干旱胁迫6 d、11 d、16 d时,叶片相对含水量分别为75.63%、73.58%、56.11%(见图3A)。处理组,干旱胁迫6 d 时,叶片相对含水量与对照无显著差异,叶片长势良好,无旱害症状(见图3A,B);干旱胁迫11 d 时,叶片相对含水量与对照无显著差异,叶片出现微卷曲特征(见图3A,C);在干旱胁迫16 d 时,叶片相对含水量极显著低于对照组,叶片明显萎蔫卷曲下垂(见图3A,D)。
2.3 干旱胁迫下杜仲叶片解剖结构特征变化
由图4 可见,从叶片的横切面可以看出,对照组叶片解剖结构细胞充盈,结构清晰,细胞间隙小,排列整齐(见图4 CK1,CK2,CK3),叶片上、下表皮厚度分别为14.43~14.48 μm、12.64~12.72 μm,栅栏组织、海绵组织厚度分别为54.19~54.36 μm、85.20~85.88 μm(见表1)。由表1 可知,随着干旱胁迫时间的延长,处理组栅栏组织的厚度与对照组相比变化不显著;在干旱胁迫6 d、11 d 时,分别与对照CK1、CK2 相比,叶片的上、下表皮细胞厚度、海绵组织厚度均变化不显著;在干旱胁迫16 d时,与对照CK3 相比,栅栏组织厚度、上表皮细胞厚度均变化不显著,叶片海绵组织厚度、下表皮细胞厚度均显著降低,分别降低了40.66%、27.04%,细胞之间相互挤压变形;在干旱胁迫6 d、11 d 时,叶片的栅海比为0.64,与对照一致;在干旱胁迫16 d 时栅海比为1.07,显著大于对照。由此说明,杜仲扦插幼苗叶片能忍受短期的干旱胁迫,叶片内部结构在短期的干旱胁迫下几乎不受影响,超过一定长时间的干旱胁迫,叶片的内部结构会发生显著变化。
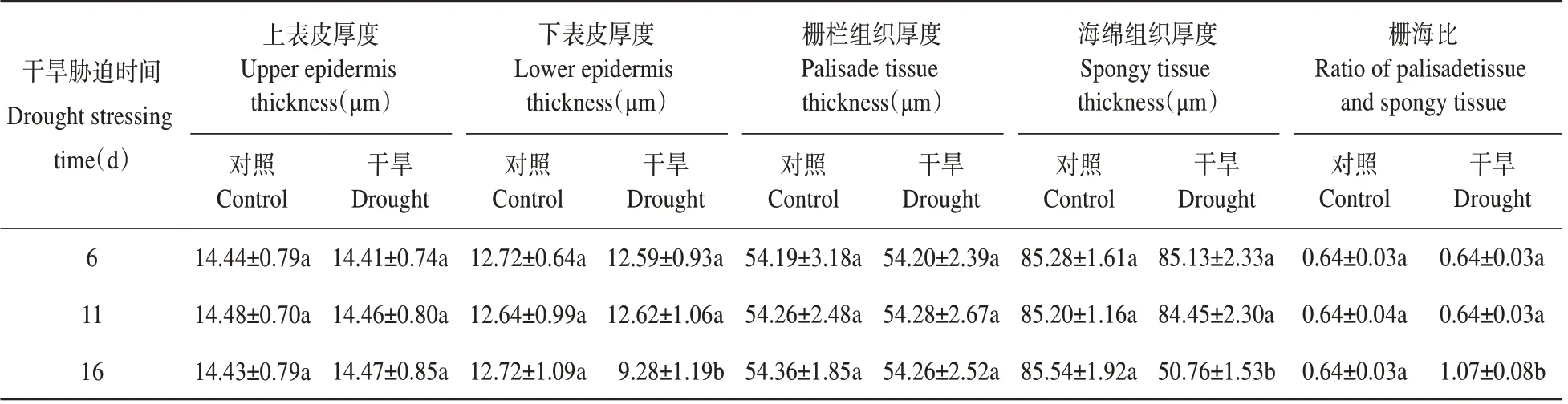
表1 干旱胁迫下杜仲叶肉结构参数Table 1 Leaf mesophyll structure parameters of E.ulmoides under drought stress
2.4 干旱胁迫对杜仲叶片下表皮气孔特征变化
图5 是不同干旱胁迫时间下杜仲叶片下表皮气孔器分布与气孔特征。由表2可知,随着干旱胁迫时间的延长,杜仲叶片气孔密度呈现先升高后下降的变化趋势;在干旱胁迫11 d 时,叶片气孔密度最大,为308.03 个·mm-2,显著大于对照;在干旱胁迫6 d 和16 d 的叶片气孔密度,与对照相比均差异不显著,分别为207.28、210.74 个·mm-2;叶片气孔面积、气孔长度、气孔宽度随着干旱胁迫时间的延长变化趋势一致,均呈先下降后升高的趋势;在干旱胁迫11 d 时,叶片气孔面积、气孔长度、气孔宽度为最小,与干旱胁迫6 d 相比,分别下降了59.22%,32.41%,28.40%,此时气孔的闭合百分比最高,显著高于对照。
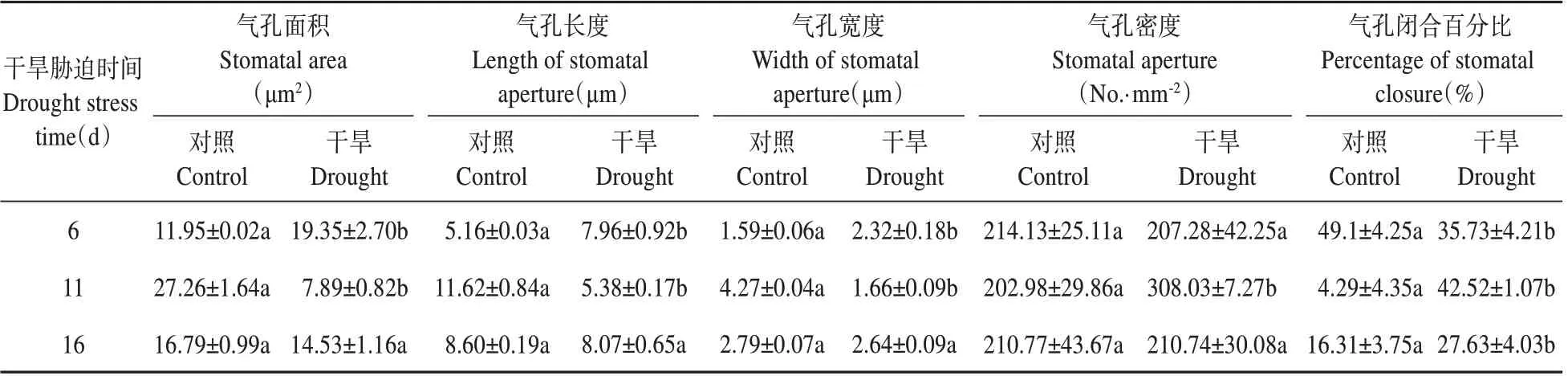
表2 干旱胁迫下杜仲叶片下表皮气孔特征值Table 2 Leaf stomatal structure eigenvalue of lower epidermis of E.ulmoides under drought stress
由表2 可知,在干旱胁迫6d 时,叶片气孔面积、气孔长度、气孔宽度均显著大于对照CK1,气孔密度差异不显著;在干旱胁迫11 d 时,叶片气孔面积、气孔长度、气孔宽度均显著低于CK2;在干旱胁迫16 d 时,叶片气孔面积、气孔长度、气孔宽度均与CK3无显著差异。
由表2 可知,对照组之间相比,叶片气孔密度相近,无大的变化;CK2 的叶片气孔面积、气孔长度、气孔宽度均明显大于CK1、CK3。
由表3可知,干旱胁迫6 d、11 d、16 d杜仲扦插幼苗叶片气孔的长度、宽度、密度相关性及线性回归方程,气孔长度与宽度呈显著正相关(R=0.896),气孔长度、宽度与气孔密度均呈显著负相关(R=0.774,R=0.795)。
3 讨论
作为直接接触外界环境的器官,叶片是植物进化过程中对环境变化比较敏感、可塑性较强的器官,其外部形态和解剖结构特征的变化能较好地反映出植物对干旱等逆境的适应性[21~23]。叶片表皮上的气孔,通过控制植物与外界的气体和水分交换来适应复杂多变的环境,当土壤持续缺水、水分高度蒸发时,气孔控制是植物限制水分丢失,防止植物受害的一种重要生理机制[24]。在研究干旱胁迫对植物生长的影响时,常用叶片气孔密度和大小的变化来进行分析[9]。
发达的栅栏组织可避免植物叶肉细胞受强光灼伤的同时,充分利用衍射光进行光合作用,有利于植物保持水分以及更好地保护内部组织,从而提高植物的存活率并有助于植物生长[25]。叶肉栅栏组织发达、海绵组织厚度相对减少及叶片组织紧密度增大等特征,有助于CO2从孔下室传导至光合作用场所,在气孔导度较低的情况下维持植物较高的光合速率,提高植物的保水能力和抗旱能力[26]。随着水分亏缺的严重,栅栏组织与海绵组织的比值会逐渐增大,叶片紧密程度不断提高,表示植株抗旱能力在增强[27]。本研究中,‘华仲6号’杜仲扦插幼苗叶片的上表皮细胞厚度较下表皮细胞厚,海绵组织厚度厚于栅栏组织,栅栏组织表现出较强的干旱适应能力其厚度受干旱胁迫的影响不大。从干旱胁迫11~16 d,杜仲叶片的相对含水量显著下降,叶片严重失水从而使海绵组织厚度显著降低,细胞之间相互挤压变形,栅栏组织与海绵组织的比值显著增大且比值大于1,叶片紧密程度提高以此来提高杜仲叶片的抗旱能力。由杨秀平[28]等人研究得知杜仲叶片上表皮细胞无气孔,具角质层,下表皮角质层薄,本研究中由于染色的原因,下表皮的角质层未看清楚。
本研究结果表明,随着干旱胁迫时间的延长,土壤含水率的降低,杜仲扦插幼苗叶片气孔密度表现为先增加后减少的趋势,而气孔面积、气孔长度和气孔宽度则表现为增加—减少—增加的趋势,且气孔长宽与气孔密度均呈显著的负线性相关,干旱胁迫下气孔的长度和宽度减少和增大具有同步性。在干旱胁迫6 d 时,可能是因为适度的干旱胁迫有助于杜仲叶片气孔的开张,对气孔密度无影响;在干旱胁迫11 d 时,叶片气孔密度显著变大可能是由于水分胁迫影响了细胞的伸长,使叶面积变小,从而提高了单位叶面积上的气孔个数,有研究表明气孔密度越大,越有利于蒸腾作用快速地将热量散掉,从而叶片可以避免因温度过高使叶绿体及原生质变性[29],气孔大小减少能降低杜仲叶片水分蒸腾,使叶片具有保水作用;在干旱胁迫16 d 时,气孔密度下降到对照水平,气孔大小变大,可能是因为叶片受到严重水分胁迫,光合作用受到严重影响,减少了植株的能量供应,植株为了维持基本的生理过程,通过增大气孔使光合作用速率不至于降到过低的程度[9]。气温也能影响气孔的大小、密度[30],本研究中,在干旱胁迫的第7~11 d,气温相对于其他两个采样时间均较高,可能是因为温度适宜叶片的生长,对照CK2 的气孔面积、气孔长度、气孔宽度均明显大于CK1 和CK3。
综上所述,随着干旱胁迫时间的延长,杜仲扦插幼苗叶片会出现严重的干枯卷曲,因此在栽培种植过程中扦插幼苗可以忍受短时间的干旱,在叶片出现萎蔫时要及时浇水。杜仲扦插幼苗厚度变薄的主要原因是海绵组织厚度变小,在干旱胁迫11 d 时,叶片气孔产生小而密、气孔闭合增多的生态适应特征,在干旱胁迫16 d 时气孔呈现大而疏的适应特征。植物抗旱性是复合性状,体现在生长形状、生理特征、光合特性、解剖结构特征、分子基因表达等方面的变化[31],本试验重点关注‘华仲6 号’扦插幼苗的叶片结构对抗旱性的响应,存在一定局限性。干旱胁迫对‘华仲6 号’幼苗叶面积的影响,以及叶面积和气孔密度之间的关联,干旱胁迫下气孔性状对叶片光合作用、蒸腾作用等重要生理过程的调节功能,温度、植物激素等与气孔开闭的关系,其他生理生化方面和分子水平方面抗旱机制有待进一步研究。
