不同浓度GnIH对鸭颗粒细胞周期和增殖的影响
2021-03-01陈世健刘文俊江丹莉欧阳宏佳黄运茂田允波
陈世健,刘文俊,杨 晨,江丹莉,欧阳宏佳,黄运茂*,田允波*
(1.仲恺农业工程学院,广州 510225; 2. 广东省水禽健康养殖重点实验室,广州 510225)
2000年,Tsutsui等[1]在鹌鹑下丘脑中发现了一种重要神经肽类激素,命名为促性腺激素抑制激素(gonadotropin inhibitory hormone,GnIH)。GnIH神经肽由12个氨基酸组成,其氨基酸序列为:Ser-Ile-Lys-Pro-Ser-Ala-Tyr-Leu-Pro-Leu-Arg-Phe,在中枢神经系统以及许多周边器官中均有分布[2]。动物的繁殖活动主要受到下丘脑-垂体-性腺轴(HPG轴)调控。研究认为,GnIH能直接或间接通过抑制下丘脑促性腺激素释放激素(GnRH)的分泌,抑制垂体促黄体激素(LH)和促卵泡激素(FSH)的合成与分泌,实现对动物繁殖的调控[3]。近年来研究发现,GnIH不仅可调控动物神经内分泌功能,抑制生殖行为,还影响和调控鸟类和哺乳动物的其他行为,比如性行为和攻击性行为[4-5]。
家禽的产卵受到卵泡发育的影响,而卵泡发育与颗粒细胞密切相关[6]。研究发现,颗粒细胞在FSH和LH的刺激下合成雌二醇和孕酮以调节卵泡的成熟和排卵[7-8],GnIH可抑制FSH和LH的合成和释放,调控卵泡的成熟和性腺的发育[9-10]。研究发现,GnIH及其受体GPR147在性腺组织也有表达,GnIH可直接与卵泡膜细胞和颗粒细胞上的结合位点相结合,进而调控细胞增殖、凋亡和类固醇的合成与分泌[11-13]。外周注射一定剂量的GnIH可降低小鼠的卵巢活性并抑制卵泡的发育,也可降低猪卵巢颗粒细胞中增殖相关蛋白ERK1/2、CyclinB1 和PCNA的表达量[14],进而促进颗粒细胞的凋亡和抑制颗粒细胞的增殖[15]。这表明,GnIH通过调节HPG轴来调控动物繁殖性能,还可直接影响卵泡颗粒细胞的增殖和激素的分泌[16]。而在家禽上,GnIH在性腺层面如何通过直接调控卵泡颗粒细胞来调控卵泡发育的机理尚不清楚。
本试验通过研究不同浓度GnIH对体外培养鸭颗粒细胞周期、增殖及相关基因表达的影响,以探究其对颗粒细胞的直接调控作用。研究结果有助于揭示GnIH在性腺层面对家禽繁殖的直接调控机制。
1 材料与方法
1.1 试验动物
试验动物为20只180日龄、体重1.5~1.6 kg的健康山麻鸭,来源于仲恺农业工程学院钟村教学科研基地。山麻鸭在实验室按动物福利原则处死后,取等级卵泡分离颗粒细胞用于细胞原代培养。
1.2 主要试剂
M199(C11150500BT)、胎牛血清(FBS)、胰酶(25200)、青链霉素(15140122)均购自美国Gibco公司;卵泡刺激素FSH(F4021)、雄烯二酮(A-084-1ML)均购自美国Sigma公司;4%多聚甲醛购自中国 Biosharp 公司;Trizol细胞裂解液(15596026)购自美国 Ambion公司;Ⅱ型胶原酶(17101-015)购自中国 Biosharp公司;细胞周期试剂盒(JX17-50T)、EdU细胞增殖检测试剂盒(C10310-1/-2/-3)均购自广州锐博生物科技有限公司。鸭GnIH多肽序列(SIKPIANMPLRF)由生工生物工程(上海)股份有限公司合成。
1.3 颗粒细胞的分离与培养
采集山麻鸭的等级卵泡,置于装有PBS缓冲液(含2%双抗)的无菌烧杯中,然后转移到细胞房超净台内,剥净卵泡表面结缔组织,用镊子撕开膜层,倒置在培养皿上剥离颗粒层。将颗粒层收集到15 mL离心管后加入5 mL培养基反复吹打,随后600 r·min-1离心3 min, 收集上清液备用。再向离心管内加入5 mL 0.1% Ⅱ型胶原酶,重悬沉淀,置于37 ℃恒温水浴锅中消化20 min,每5 min震荡1次。消化结束后加入5 mL M199完全培养基(含10% FBS),用70 μm 细胞筛过滤(同时过滤上清液)至50 mL离心管,1 000 r·min-1离心10 min,弃上清,沉淀用M199 洗1次,1 000 r·min-1室温离心10 min后加入3 mL的M199完全培养基(含10% FBS 及2%双抗),重悬后以每孔1×106个细胞的密度接种于6孔 细胞培养板,置于39 ℃、5% CO2的培养箱内静置培养。培养液含不同浓度GnIH(0、0.1、1、10和100 ng·mL-1)、5% FBS、2%双抗、终浓度为1 ng·mL-1的FSH、终浓度为0.1 μmol·mL-1的雄烯二酮。
1.4 细胞周期检测
将分离到的颗粒细胞以每孔5×105个细胞的密度接种于12孔板,用不同浓度GnIH(0、0.1、1、10和100 ng·mL-1)处理24 h后进行细胞周期检测。用胰酶消化吹打细胞,制备单细胞悬液,用1.5 mL 离心管收集细胞,1 000 r·min-1离心5 min后弃上清,细胞沉淀用1 mL预冷的PBS重悬后1 000 r·min-1离心5 min,弃上清,细胞沉淀用1 mL 预冷的75%乙醇轻轻混匀,-20 ℃固定过夜。固定细胞后加入1 mL PBS室温水合15 min。离心收集细胞,弃上清,每管加入500 μL碘化丙啶染色液,37 ℃避光条件下孵育30 min。以标准程序用流式细胞仪检测,一般计数2万~3万个细胞,结果用FlowJo 7.6软件分析细胞周期。
1.5 EdU细胞增殖检测
将分离到的颗粒细胞以每孔5×105个细胞的密度接种于12孔板,用不同浓度GnIH(0、0.1、1、10和100 ng·mL-1)处理24 h后进行EdU细胞增殖检测。每孔加入1 mL 50 μmol·L-1EdU培养基孵育2 h,弃培养基,PBS清洗细胞2次,每次5 min。然后加入500 μL细胞固定液室温孵育30 min,弃固定液。再加入500 μL甘氨酸(2 mg·mL-1),脱色摇床孵育5 min后用PBS清洗5 min。添加1 mL渗透剂(0.5% TritonX-100的PBS)脱色摇床孵育10 min后用PBS清洗5 min,每孔加入1 mL的Apollo®染色反应液,避光、室温、脱色摇床孵育30 min。然后,加入1 mL渗透剂脱色摇床清洗3次,每次10 min。 每孔加入1 mL Hoechst33342反应液,避光、室温、脱色摇床孵育30 min后用PBS清洗2次。最后,每孔加入1 mL PBS,在倒置荧光显微镜下观察细胞。
1.6 增殖相关基因表达的检测
取不同浓度GnIH(0、0.1、1、10和100 ng·mL-1)处理24 h后的6孔板细胞,弃培养液后用Trizol试剂从颗粒细胞中提取总RNA,根据东洋纺反转录试剂盒ReverTra Ace qPCR RT Master Mix with gDNA Remover说明书进行反转录,获得颗粒细胞cDNA。荧光定量检测增殖相关基因IGF-2、IGFBP-2、CyclinD1、CDK6、p27kip1的基因序列从NCBI 获得,利用软件Primer 5.0设计引物,引物序列见表1,由生工生物工程(上海)股份有限公司合成。荧光定量使用Power UPTM SYBR®Green Master Mix 试剂,反应体系为20 μL:Mix 10 μL,水 8.6 μL,上、下游引物各0.2 μL,cDNA 1 μL。反应条件:95 ℃预变性10 min;95 ℃变性15 s,60 ℃退火1 min,共40个 循环。以GAPDH为内参基因,每个样品进行3次 重复,检测结果用2-△△CT法进行统计分析。

表1 qRT-PCR 相关引物Table 1 The related primers of qRT-PCR
1.7 统计分析
数据处理采用GraphPad Prism 7.0的单因素方差分析,不同处理间的显著性检验采用Tukey式多重比较。数据结果采用“Mean±SEM”表示(n=3),P<0.05表示差异显著,P>0.05表示差异不显著。
2 结 果
2.1 不同浓度GnIH对颗粒细胞生长状态的影响
在倒置荧光显微镜下观察不同浓度GnIH处理后颗粒细胞的生长状态。结果显示(图1),各GnIH处理组细胞形态正常,细胞轮廓清晰,视野内有极少量细胞碎片化,呈漂浮死亡状态;随着GnIH处理浓度的升高,死亡细胞的数量增加,但组间差异不显著(P>0.05)。
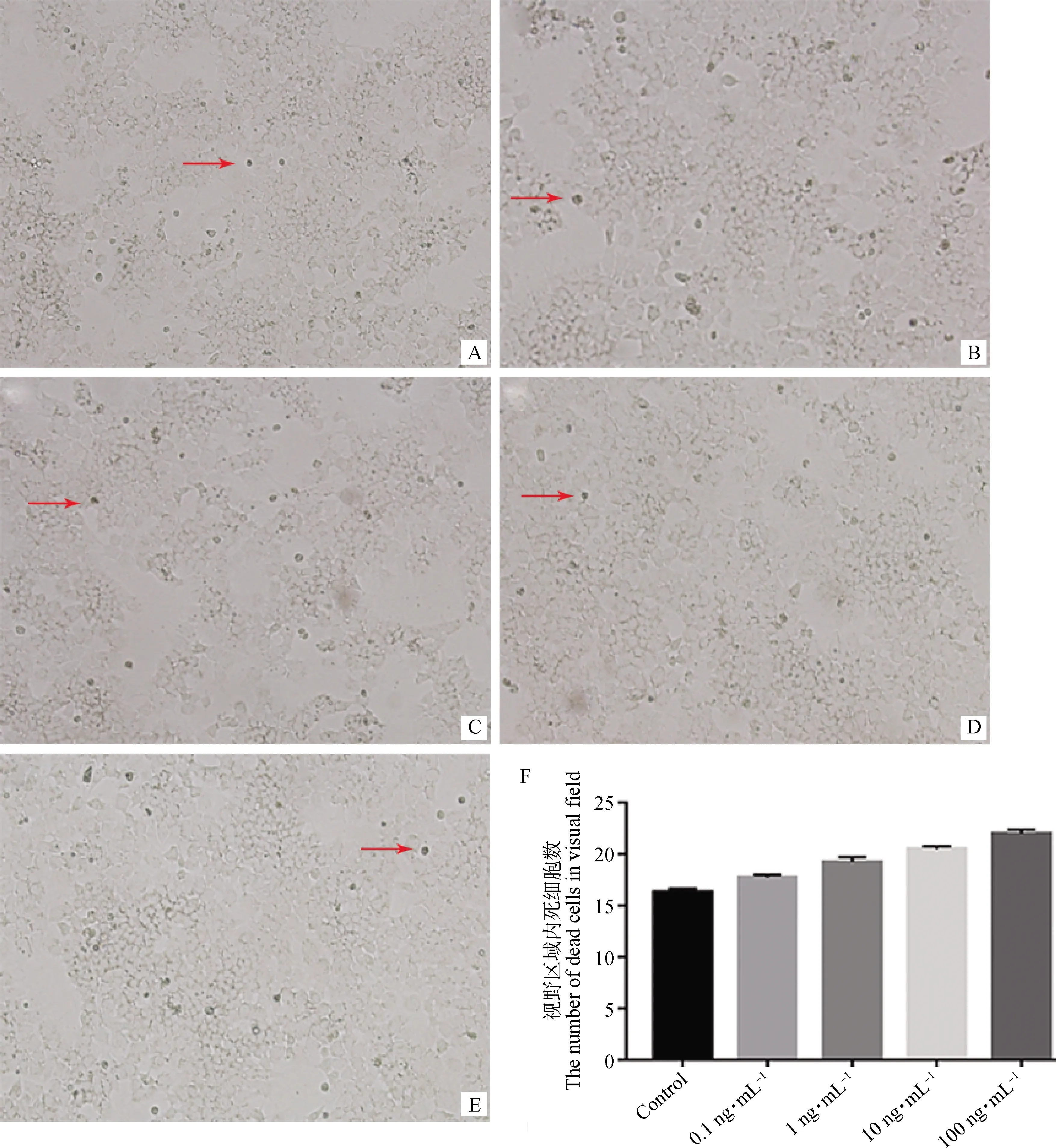
A. 对照组(400×);B. 0.1 ng·mL-1 GnIH处理组(400×);C. 1 ng·mL-1 GnIH处理组(400×);D. 10 ng·mL-1 GnIH处理组(400×);E. 100 ng·mL-1 GnIH处理组(400×);F. 视野区域内死细胞个数。红色箭头指死细胞A. Control group (400×); B. 0.1 ng·mL-1 GnIH treatment group (400×); C. 1 ng·mL-1 GnIH treatment group (400×); D. 10 ng·mL-1 GnIH treatment group (400×); E. 100 ng·mL-1 GnIH treatment group (400×); F. The number of dead cells in visual field. The red arrows indicate dead cells图1 不同浓度GnIH对颗粒细胞生长状态的影响Fig.1 Effects of different concentrations of GnIH on growth of granulosa cells
2.2 不同浓度GnIH对颗粒细胞周期的影响
用流式细胞术检测不同浓度GnIH处理对颗粒细胞周期的影响。结果显示(图2),与对照组相比,各GnIH处理组细胞在G1期的比例有所下降,其中,1 ng·mL-1GnIH处理组细胞在G1期的比例显著降低(P<0.05);在0.1 和1 ng·mL-1GnIH处理组,细胞在G2期的比例显著升高(P<0.05);各GnIH处理组细胞在S期的比例均无显著差异(P>0.05)。

A. 对照组流式细胞术结果图;B. 0.1 ng·mL-1 GnIH处理组流式细胞术结果图;C. 1 ng·mL-1 GnIH处理组流式细胞术结果图;D. 10 ng·mL-1 GnIH处理组流式细胞术结果图;E. 100 ng·mL-1 GnIH处理组流式细胞术结果图;F. GnIH处理组处于不同周期细胞所占百分比。不同小写字母表示差异显著(P<0.05),相同小写字母表示差异不显著(P>0.05),下同A. The flow cytometry results of control group; B. The flow cytometry results of 0.1 ng·mL-1 GnIH treatment group; C. The flow cytometry results of 1 ng·mL-1 GnIH treatment group; D. The flow cytometry results of 10 ng·mL-1 GnIH treatment group; E. The flow cytometry results of 100 ng·mL-1 GnIH treatment group; F. The percentage of cells in different cell cycles in GnIH treatment groups. Different lowercase letters indicate significant differences (P<0.05), and the same lowercase letter indicates no significant differences (P>0.05), the same as below图2 不同浓度GnIH对颗粒细胞周期的影响Fig.2 Effects of different concentrations of GnIH on cycle of granulosa cells
2.3 不同浓度GnIH对颗粒细胞增殖的影响
用EdU方法检测不同浓度GnIH处理对颗粒细胞增殖的影响。结果显示(图3),与对照组相比,随着GnIH处理浓度的升高,EdU阳性细胞所占的百分比逐步降低;在1、10和100 ng·mL-1GnIH处理组中EdU阳性细胞率显著降低(P<0.05)。

A.从各GnIH处理组中随机选择5张合并图像(400×);B. 从荧光图像中获得EdU阳性细胞数,计算EdU相对阳性率A.Five merged images randomly selected from each GnIH treanment group(400×); B. The number of EdU-positive cells obtained from the fluorescence image, and the relative positive rate of EdU was calculated图3 不同浓度GnIH对颗粒细胞增殖的影响Fig.3 Effects of different concentrations of GnIH on proliferation of granulosa cells
2.4 不同浓度GnIH对颗粒细胞增殖相关基因表达的影响
用qRT-PCR方法检测不同浓度GnIH处理对颗粒细胞增殖相关基因表达的影响。结果显示(图4),与对照组相比,在0.1和1 ng·mL-1GnIH处理组中,CDK6、CyclinD1、IGF-2、IGFBP-2和p27kip1基因的相对表达量均下降,但差异不显著(P>0.05);在10 和100 ng·mL-1GnIH处理组中,CyclinD1、IGFBP-2和p27kip1基因的相对表达量均显著上升(P<0.05)。
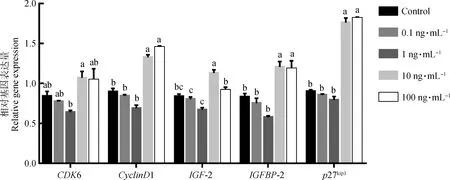
图4 不同浓度GnIH对颗粒细胞增殖相关基因表达的影响Fig.4 Effects of different concentrations of GnIH on proliferation-associated genes expression in granulosa cells
3 讨 论
GnIH及其同源物RFRP-3是一种重要的神经肽类激素,可抑制性腺激素的产生和影响生殖细胞的分化与成熟[17-19]。研究表明,GnIH可直接或间接通过下丘脑GnRH调控垂体促性腺激素的分泌,对禽类繁殖性能发挥重要调控作用,并且在下丘脑能与褪黑素、kisspeptin等生殖相关神经肽互作共同调节动物的生殖活性[20-21]。除了在垂体层面,GnIH可降低垂体前叶LH和FSH的分泌[22],有研究显示,在性腺层面,GnIH可直接抑制卵泡颗粒细胞中类固醇激素E2和T的分泌水平,对动物卵泡发育、闭锁及排卵等发挥重要作用[23-25]。但目前在家禽上,关于GnIH直接在性腺层面调控卵泡发育及闭锁的机理仍不清楚。因此,本试验在体外培养条件下,研究了不同浓度GnIH对卵泡发育紧密相关的颗粒细胞周期、增殖及相关基因表达的影响,所得结果一定程度上揭示了GnIH通过调节颗粒细胞影响卵泡发育的机理。
细胞周期进程对家禽卵泡颗粒细胞的增殖有至关重要的影响。研究表明,在细胞增殖过程中,周期蛋白依赖性蛋白激酶(CDKs)对细胞周期产生重要的调控作用[26],其中CyclinD1与CDK6 在G1期形成复合物促使细胞由G1期进入S期,进而促进细胞的增殖[27]。CKI家族因子p27kip1对CDK则具有广谱的抑制作用,与CDK-Cyclin复合物结合来调控细胞周期进程[28]。本研究中,用外源性GnIH处理鸭颗粒细胞后,EdU阳性细胞所占百分比下降,细胞死亡数增多,G2期细胞所占百分比增加,结果表明,GnIH处理导致鸭颗粒细胞增殖受到抑制,且细胞凋亡增多,其中主要原因是GnIH处理将细胞周期阻滞在G2期。检测与细胞周期紧密相关基因的表达发现,GnIH处理导致细胞周期促进因子CDK6和CyclinD1的表达降低,细胞周期抑制因子p27kip1的表达升高。这揭示了GnIH处理是通过影响与细胞周期紧密相关因子CDK6、CyclinD1和p27kip1的表达来抑制鸭颗粒细胞的细胞周期以及增殖。本结果在相关研究中得到进一步验证,有研究表明,GnIH处理猪卵巢颗粒细胞后降低了细胞周期相关基因的表达,并导致细胞周期阻滞在G2/M期[29];也有研究证实,GnIH能降低颗粒细胞活力,并调控周期蛋白CyclinB1、PCNA以及其他多种周期因子的表达来阻滞细胞周期进程,影响颗粒细胞的增殖和分化[30-31]。此外,细胞增殖相关基因IGFBP-2属于IGFBPs家族的重要成员之一,能直接或间接地影响IGF-2的功能,进而影响细胞的有丝分裂和增殖[32]。有研究报道,在猪卵泡颗粒细胞上,IGF-2和IGFBP-2基因表达的上调会促进颗粒细胞的增殖[33]。本研究中,GnIH处理下调了鸭颗粒细胞中IGF-2和IGFBP-2的表达,且变化趋势与CDK6和CyclinD1一致,结果与国内外相关的研究报道一致。这进一步揭示,IGF-2和IGFBP-2参与了颗粒细胞增殖的调控。另外,在本研究中,高浓度GnIH处理组会导致鸭颗粒细胞中部分增殖相关基因表达的上升,其中原因还不得而知。一方面可能是与本研究在体外培养条件下开展有关,另一方面可能与这种高剂量外源性GnIH所引起的一些细胞内非正常作用有关,具体原因还需进一步研究。
动物的繁殖行为和能力受多种环境因素的影响,但环境因子都需要通过神经-内分泌信号传导影响HPG轴来调控动物繁殖活性。而GnIH作为下丘脑重要的内分泌激素,通过生殖轴对禽类繁殖行为发挥重要调控作用[34]。GnIH不仅在生殖轴上游的下丘脑和垂体通过抑制促性腺激素的分泌调控动物繁殖活性,而且还可以在性腺层面直接调控禽类卵泡发育。大量研究表明,外周注射GnIH可抑制成年鸡的性腺活性,诱导睾丸细胞凋亡,降低睾丸的生精活性[35-36];在鹅的繁殖调控中,GnIH在繁殖后期维持高水平表达,促进鹅由繁殖期向休产期转变[37-38];在鹌鹑繁殖期中,GnIH与褪黑素具有协同作用,能降低血浆睾酮水平,影响繁殖性能[39-40];GnIH处理还可引起成年小鼠精子发生的剂量依赖性组织学变化,如生殖细胞增殖、存活标志物下降、睾丸凋亡标志物增加[41-42]。本研究结果也表明,在体外培养条件下,GnIH处理能通过影响周期相关因子的表达来抑制鸭颗粒细胞周期和增殖。因此,GnIH作为禽类生殖调控的重要因子,系统揭示了GnIH在生殖轴3个层面的作用机制,对深入了解禽类繁殖机理和开发提高禽类繁殖性能的技术均具有积极意义。
4 结 论
在体外培养的鸭颗粒细胞中,添加外源性GnIH能降低细胞周期促进因子CDK6、CyclinD1、IGF-2、IGFBP-2的表达,提高细胞周期抑制因子p27kip1的表达,并且将细胞周期阻滞在G2期,降低EdU阳性细胞百分率,进而抑制颗粒细胞的增殖。本结果对GnIH在性腺层面通过影响颗粒细胞调控禽类繁殖性能机理的研究具有积极意义。
