Resazurin as an indicator of reducing capacity for analyzing the physiologic status of deep-sea bacterium Photobacterium phosphoreum ANT-2200*
2021-02-22JieDAIHongzhiTANGXuegongLIClaireLiseSANTINWenpengCUINaLIUXiaoqingQIXuehuaCUIAlainGROSSIPhilippeNOTARESCHIWeiJiaZHANGLongFeiWU
Jie DAI , Hongzhi TANG , Xuegong LI , Claire-Lise SANTIN, , Wenpeng CUI , Na LIU , Xiaoqing QI , Xuehua CUI , Alain GROSSI , Philippe NOTARESCHI , Wei-Jia ZHANG , , Long-Fei WU , ,
1 Laboratory of Deep Sea Microbial Cell Biology, Institute of Deep Sea Science and Engineering, Chinese Academy of Sciences, Sanya 572000, China
2 University of Chinese Academy of Sciences, Beijing 100049, China
3 International Associated Laboratory of Evolution and Development of Magnetotactic Multicellular Organisms (LIA-MagMC), CNRS-CAS
4 Aix Marseille University, CNRS, LCB, IMM, FR3479, IM2B, Marseille 13402, France
5 Aix Marseille University, CNRS, IMM, FR3479, Marseille 13402, France
Abstract Resazurin (RZ) is a weakly fluorescent blue dye and can be reduced irreversibly to highly fluorescent pink resorufin (RF) that is reduced reversibly to colorless dihydroresorufin (hRF) by photodeoxygenation, chemical reaction and reductive organic compounds produced through cell metabolism. Because of the reliable and sensitive fluorescence-color change and noninvasive features, RZ has been used widely as a redox indicator in cell viability/proliferation assays for bacteria, yeast, and mammalian cells. However, RZ is used rarely for physiological characterization of marine microorganisms. Here, we developed a custom-made irradiation and absorption-analysis device to assess the reducing capacity and physiologic status of marine bacterial cultures. We measured the absorption spectra of RZ, RF, and hRF in the presence of the reducing compound Na 2 S and under visible-light irradiation. After establishing appropriate parameters, we monitored the color changes of RZ and its reduced derivatives to evaluate the coherence between reducing capacity, bioluminescence and growth of the deep-sea bacterium Photobacterium phosphoreum strain ANT-2200 under various conditions. Emission of bioluminescence is an oxidation process dependent upon cellular reducing capacity. Growth and bioluminescence of ANT-2200 cell cultures were impeded progressively with increasing concentrations of RZ, which suggested competition for reducing molecules between RZ at high concentration with reductive metabolism. Therefore, caution should be applied upon direct addition of RZ to growth media to monitor redox reactions in cell cultures. Analyses of the instantaneous reduction velocity of RZ in ANT-2200 cell cultures showed a detrimental effect of high hydrostatic pressure and high coherence between the reducing capacity and bioluminescence of cultures. These data clearly demonstrate the potential of using RZ to characterize the microbial metabolism and physiology of marine bacteria.
Keyword: oxic-reduction indicator; absorption spectra; bacterial growth; marine microorganism
1 INTRODUCTION

Fig.1 Molecular structure and absorption spectra of resazurin, resoruf in and dihydroresoruf in
Resazurin (RZ) has a chemical structure of 7-hydroxy-10-oxidophenoxazin-10-ium-3-one and is also referred to as a heterocyclic N-oxide or N-phenoxazine-3-one dye. RZ exhibits a blue color with weak fluorescence. As a donor in reactions involving transfer of oxygen atoms, RZ is reduced irreversibly to the highly fluorescent pink dye resorufin (RF). Further reduction of RF results in the production of colorless dihydroresorufin (hRF), which is a reversible process (Fig.1). RZ is soluble in water with low or non-cytotoxicity. Therefore, RZ and derivatives of N-phenoxzine-3-one dyes have been used widely as simple, rapid, reliable, sensitive, safe, and cost-effective agents for measurement of cell viability and proliferation, and for determination of bacterial and yeast contamination in milk (Neumann et al., 1996; Rampersad, 2012; Kowaltowski, 2019). The most widely used bioassay kits derived from RZ are provided by various companies under the product name of “Alamar Blue” (O’Brien et al., 2000). Moreover, the anionic dye RZ is an appropriate probe for analytical determination of Se(IV) and Pb(II) ions (Safavi et al., 1990; Afkhami et al., 1991). However, RZ dyes are rarely utilized in physiologic studies of marine microorganisms.
Reduction of the phenoxazine-3-one dyes RZ and RF occurs through chemical reactions and metabolic processes. In addition, irradiation in the presence of amines leads to deoxygenation of RZ and its conversion to RF (Neumann et al., 1996; Bueno et al., 2002). Similarly, reduction of RF is a photosensitive reaction. Zhao et al. (2011) reported that ultraviolet (UV) and visible light ( λ >300 nm) irradiation leads to one-electron reduction of RF and produces a semiquinoneimine type anion radical RF*−under anaerobic conditions with reduced nicotinamide adenine dinucleotide (NADH). Continuous irradiation reduces RF*−further to its colorless form of hRF (Fig.1).
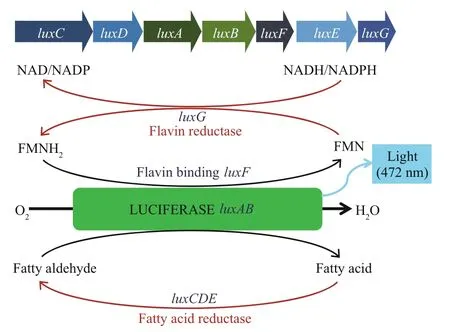
Fig.2 Bioluminescence of strain ANT-2200
Bioluminescent bacteria have the peculiar capacity of emitting blue-green light (470-490 nm) through the luciferase-catalyzed oxidation of reduced flavin mononucleotide (FMNH) and fatty aldehyde (Fig.2) (Dunlap and Kita-Tsukamoto, 2006). To continue bioluminescence, oxidized FMN must be reduced to FMNH by an enzyme using NADH as an electron donor (Fig.2). Most luminous bacteria dwell in marine environments from the shallow coast to the deep sea (Dunlap and Kita-Tsukamoto, 2006). The Photobacterium phosphoreum strain ANT-2200 (hereafter termed “ANT-2200”) has been isolated at a depth of 2 200 m from the Mediterranean Sea and can emit bioluminescence (Al Ali et al., 2010; Martini et al., 2013). Genomic and physiologic analyses have revealed a versatile potential for energy metabolism and growth capacity of ANT-2200 cells by: deriving energy from fermentation of glucose or maltose; respiration with formate as an electron donor and trimethlyamine N-oxide (TMAO), nitrate or fumarate as electron acceptors; by chemo-organo-heterotrophic growth in rich media (Zhang et al., 2016). Interestingly, the light emission of ANT-2200 cells seems to be proportional with their growth rate. Continuous emission of bioluminescence is dependent upon the reductive capacity of cells, and bio-luminous light might trigger the photo-reduction of RZ and RF. We wondered if it is feasible to use RZ dyes to assess the physiologic status of ANT-2200 cells by analyzing the correlation among reducing capacity, bioluminescence and growth. We set-up an irradiation and spectrum-scanning device and showed the reproducible relationship of RZ reduction with the growth and luminescence of ANT-2200 cells. We demonstrated potential usefulness of resazurin, at appropriate concentration, in monitoring growth and metabolism of marine bacteria.
2 MATERIAL AND METHOD
2.1 Strain and bacterial cultures
Cultures of the strain ANT-2200 were undertaken in YPG media in serum bottles at 27 °C (Supplementary Fig.S1), or in 5-mL syringes immersed in highpressure incubators, as described by Zhang et al. (2016). When indicated, hydrostatic pressure of 20 MPa was applied on the high-pressure incubators that contained the syringes. This medium was prepared with seawater and pH 8.9 allows fast growth and the highest bioluminescence. A stock solution of RZ at 20 mmol/L was prepared in Milli-Q water (Millipore, Bedford, MA, USA) and used at the final concentrations indicated. The sodium salt of RZ was purchased from Sangon Biotech (Shanghai, China).
To analyze the effect of irradiation, RZ solutions or bacterial cultures were exposed to tungsten lightbulbs or light-emitting diodes (LEDs) at 470 nm. Light densities were measured using a sensor (S120vc; Thorlabs, Newton, NJ, USA) connected to a power meter (Thorlabs).
Samples were taken at the indicated time. Absorbance at 600 nm was measured using a spectrophotometer and luminescence was analyzed using a microplate reader according to manufacturer instructions.
2.2 Absorption spectroscopy
Absorption spectroscopy of RZ and its reduced derivatives was undertaken in serum bottles directly as a solution or in growth media at the concentrations indicated. Alternatively, for post-growth scanning, 7−8-mL cultures were transferred to serum bottles and RZ was added at a final concentration of 0.01 mmol/L; the bottles were sealed and scanned for absorption spectroscopy every 5 min until the end of color change using a UV-Vis spectrophotometer (Varian 50, Varian, Pala Alto, CA, USA; Cary60, Agilent Technologies, Santa Clara, CA, USA). A bottle holder was designed and fabricated by threedimensional (3D) printing (Supplementary Fig.S1). 3 RESULT AND DISCUSSION
3.1 Spectral characteristics of RZ and its derivatives
3.1.1 Setup of a custom-made device to facilitate irradiation and absorption spectroscopy of cultures
RZ and RF are highly sensitive to oxidation, so tightly sealed containers should be used for bacterial cultivation and absorption spectroscopy. A serum bottle of 11 mL was ideal for our study for three main reasons. First, it could be sealed tightly and addition of chemical solutions or exchange of gases could be done using needles piercing rubber plugs. Second, culturing of marine bacteria such as the ANT-2200 strain with this bottle could provide suき cient biomass for further analyses. Third and most importantly, the bottle fitted in most standard spectrophotometers. To use this bottle with UV-Vis spectrophotometers (Varian 50 and Cary 60), we designed a 3D-printed bottle-holder for direct absorption spectroscopy (Supplementary Fig.S1). In addition, LED light bulbs could be mounted readily on the holders to irradiate the solution or bacterial cultures. The circular shape of the bottle could have introduced variations in absorbance readings, but the impact was minimal because absorbance values were used as empirical proxies.
Using our bottle in the Cary 60 UV-Vis spectrophotometer, we observed a principal absorption at 601 nm and a shoulder peak at 550 nm of RZ in water solution saturated with N2(Fig.1b). Under a highly controlled redox state based on Na2S concentrations, RZ was reduced irreversibly to RF by Na2S, which resulted in a shift of the spectrum, leading to a peak at 571 nm and shoulder peak at ~530 nm (Fig.1b). Further reduction to hRF led to disappearance of these absorption peaks. The absorption spectra of RZ, RF, and hRF were similar to those of previous reports, although a slight shift of absorption peaks (possibly caused by different media and spectrophotometers) was observed. Therefore, this device allowed easy monitoring of the reduction states of RZ, RF, and hRF in a sealed serum bottle, and peaks at 601 nm and 571 nm were used to represent RZ and RF, respectively, in the present study.
3.1.2 Effect of irradiation and Na2S on the absorption spectra of RZ and its derivatives
First, we assessed the color change of RZ under different redox conditions in artificial seawater in a fermenter at controlled redox conditions and pH. The solution was blue and remained unchanged after overnight bubbling with N2. Switching N2to CO2increased the redox potential from -90 mV to -53 mV and decreased the pH of the solution from 8.9 to 7.9, but had no effect on its blue color. Therefore, RZ solution was stable in the range of redox potential and pH variation tested, which was modified by supplementation of carbon dioxide. Decreasing the redox condition of the solution was not suき cient to convert RZ to RF.
Other than pH and redox conditions, irradiation has been reported to lead to deoxygenation of RZ and reduction of RF under certain conditions (Porcal et al., 2011). Consistently, we found that upon exposure to sunlight or illumination with a tungsten lightbulb, the solution became light-pink. This change implied RF formation from photochemical deoxygenation of RZ under these redox and pH conditions in artificial seawater solution.
Next, we evaluated the effect of irradiation and chemical reaction using our bottle-scanning device. The 11-mL serum bottles were filled with 8 mL of water containing 7.2 μmol/L RZ and bubbled with N2for 15 min. Then, Na2S was injected into the bottles to a final concentration of 72 μmol/L (Supplementary Fig.S2, bottles 1 and 6), 720 μmol/L (bottles 2 and 7), 3 600 μmol/L (bottles 3 and 8) and 7 200 μmol/L (bottles 4 and 9). As a control, bottles 5 and 10 contained only RZ without Na2S. The bottles were incubated in the dark (bottles 1-5) or with illumination from a tungsten lightbulb (bottles 6-10, at 25 mW/cm2). Visual observation showed that RZ remained blue in the absence of Na2S (Supplementary Fig.S2a). At a 10-fold molar ratio of Na2S:RZ (bottles 1 and 6), the solutions turned pink (RF), whereas all solutions became colorless (hRF) with a molar ratio of Na2S:RZ of >100-fold.
We further quantified the effect of reductants and irradiation by monitoring the time-lapse of absorption spectra. The absorption spectra of RZ remained unchanged during 24 h in the absence of Na2S and light (Fig.3a). However, the absorption spectra diminished slightly or substantially after 12 h and 24 h of irradiation with a tungsten lightbulb, respectively (Fig.3b). Notably, the absorption spectra revealed changes that were unperceivable by the naked eye. Therefore, irradiation led to photobleaching and photo-deoxygenation of RZ, as reported by Porcal et al. (2011).
When adding Na2S, a decrease of RZ spectra was concomitant with rise of RF spectra. With the concentration increasing, the latter began decline after reaching the maximum. The spectral change was related to the concentration of Na2S (Fig.3c-f). Upon incubation in the dark, addition of a 10-fold amount of Na2S led to a decrease in absorption at 601 nm from 0.78 to 0.33 and increase of absorption at 571 nm from 0.54 to 0.82 within 96 min (Fig.3c & g). Notably, absorption at 571 nm continued to increase under this condition (Fig.3g, dash-red curve), indicating RF was not reduced further to hRF within this analysis time. The reduction of RZ→RF→hRF could be accelerated by irradiation. Upon irradiation exposure, the peak at 601 nm decreased rapidly to the baseline level at 15 min (Fig.3d & g, blue curve), then absorption at 571 nm reached its maximum and began to decrease gradually (Fig.3g, red curve). By the end of analyses at 40 min, RF was partially reduced to transparent hRF, which led to lower absorption at 571 nm and a lighter pink color of the solution (Fig.3d and Supplementary Fig.S2, bottle 6).
As demonstrated above, the amount of reductant had a considerable influence on RZ reduction. When excess Na2S (molar ratio of Na2S:RZ=500) was added, absorption at 601 nm decreased rapidly to the baseline level in ~1 min (Fig.3e, f and blue curves in h). Absorption at 571 nm increased to a maximal level at 30 s, and then decreased to the approximate baseline level at 2.5 min after addition of Na2S, respectively. Irradiation did not change velocity of reduction (Fig.3h), which indicated a dominant effect of chemical reduction over the effect of irradiation upon addition of excess Na2S.
Taken together, these results show that Na2S chemically reduced RZ to RF and hRF, and irradiation with visible light led to photo-deoxygenation of RZ and accelerated reduction in our bottle-scanning system. Evolution of the absorption peak at 571 nm resulted from reduction of RZ in combination with further reduction of RF to hRF. In contrast, the irreversible reduction of RZ was truly reflected by a decrease in absorption at 601 nm and thus could be an eき cient indicator of reducing capacity in the reaction system. To quantify and compare the reducing capacity between different samples, the velocity of instantaneous reduction was calculated using the following equation:

Fig.3 Effect of Na 2 S and irradiation upon the absorption spectra of resazurin, resoruf in and dihydroresoruf in

Fig.4 Influence of RZ on ANT-2200 the growth and bioluminescence of ANT-2200 cells
Vn=( A601_n+1- A601_n)/( t_n+1- t_n), where t_nis a point in time series and A601_nis the absorption at 601 nm at that time.
The velocity of instantaneous reduction increased from 0.005 to 0.08 A601/min upon illumination at a limited amount of Na2S (Fig.3g), whereas it reached a maximum of 1.04 and 1.05 A601/min at excessive Na2S independently of illumination (Fig.3h). Vnwas used to describe the reducing capacity of the cultures in the present study.
3.2 Using RZ as an indicator of reducing capacity to evaluate the physiologic status of ANT-2200 cells
3.2.1 Effect of RZ on the growth of ANT-2200 cells
Bioluminescent bacteria emit light by a highly conserved FMNH oxidation reaction (Fig.2). To continue such emission, the oxidized FMN must be reduced back to FMNH by NADH or NADPH, which is catalyzed by the flavin reductase LuxG in marine luminous bacteria (Nijvipakul et al., 2008). In parallel, RZ is converted to RF by the reductants NADPH or NADH in the presence of the enzyme NADPH dehydrogenase or NADH dehydrogenase in living cells (De Jong and Woodlief, 1977; Barnes and Spenney, 1980). Therefore, RZ might compete with LuxG for the reductant NADPH/NADH and affect bioluminescence. To test this hypothesis, we analyzed the effect of addition of RZ on ANT-2200 cell cultures. Interestingly, the growth of ANT-2200 cells was hampered progressively with increasing quantities of RZ in the growth media (Fig.4a). Moreover, the bioluminescence was decreased in parallel (Fig.4b).

Fig.5 Coherence of reducing capacity with growth and bioluminescence of ANT-2200 cell cultures
Next, we analyzed the reducing capacity of these cultures. RZ in the growth media was converted irreversibly to RF during the growth of ANT-2200 cells, so we followed the changes in the characteristic absorption peaks of RF at 571 nm instead. Upon shaking the culture bottles, hRF generated during the growth of ANT-2200 cells was re-oxidized to RF in the air phase in the culture bottles. Then, RF was reduced to hRF by ANT-2200 cells. By measuring time-lapse absorption spectra, we compared the rate and extent of RF reduction that reflect the physiologic status of these cultures. Absorption at 571 nm of the culture with 0.01 mmol/L RZ decreased to the baseline level 25 min after shaking (Fig.4c). However, it took 30 min for the RF peak to reduce to ~50% with cells cultured with 0.05 mmol/L RZ (Fig.4d). These results suggest that 0.05 mmol/L RZ might be at excessive amount to be reduced completely by ANT-2200 cells. The excessive amount of RZ in the media might also have accounted for its inhibitory effect upon the growth and bioluminescence of ANT-2200 cells. Therefore, caution should be taken if RZ is added to media to monitor the redox reaction of cultures.
3.2.2 Coherence of using bioluminescence and instantaneous reducing velocity to analyze the physiologic status of ANT-2200 cell cultures
The deep sea luminous strain ANT-2200 has versatile potential in terms of energy metabolism (Zhang et al., 2016) and its light emission seems to be proportional to its growth rate. We evaluated the reducing capacity of cultures of marine bacteria by inspection of growth, bioluminescence and reducing velocity of RZ under different growth conditions.
The growth of ANT-2200 cells under high hydrostatic pressure was slightly slower than that at atmospheric pressure, and cultures at both pressure conditions reached a stationary phase after ~10 h of incubation (Fig.5a). However, the cultures at atmospheric pressure exhibited a second growth phase and attained maximal absorbance of 1.32 at 24 h, and retained this absorbance after 50 h of incubation (Fig.5a). Consistently, bioluminescence increased during the exponential growth phase and decreased in the stationary growth phase, and the cultures at atmospheric pressure were more luminous than those under high hydrostatic pressure (Fig.5b). Notably, the instantaneous reducing velocity of RZ increased during the exponential growth phase and decreased at the stationary growth phase, and the instantaneous reducing velocity of RZ of the cultures at atmospheric pressure was higher than that under high pressure (Fig.5c), which indicates degeneration of piezophile property of ANT-2200. Taken together, these results showed that the instantaneous reducing velocity truly reflected the physiologic status of cultures, and was suitable for evaluating the metabolism of marine bacteria.
The proof-of-concept experiment showed the great potential of using RZ for physiologic and metabolic studies of marine microorganisms. Moreover, we used the RZ→RF reaction to monitor the redox state of growth media at light-blue/pink to inoculate microaerobic marine bacteria, of which the color-redox correlation was pre-established in a fermenter. Similarly, the color change between RF and hRF allowed adjustment to a suitable condition for the growth of a marine archaea by adding Na2S and changing the gas composition of culture media. Besides equilibrating the redox reaction in media before inoculation, we also used the reducing capacity (i.e., the incubation time required for change of pink RF to colorless hRF) to screen for optimal culture-media and incubation conditions (temperature and hydrostatic pressure) for deep-sea microorganisms.
4 CONCLUSION
RZ is a sensitive indicator of reducing capacity. We assessed the feasibility of using RZ as an indicator of reducing capacity to evaluate the physiologic state of marine luminous bacteria. RZ exhibits cytotoxicity at high concentrations, and its use in cultures should be undertaken with extreme caution. However, we found an obvious coherency between the reducing capacity of the cultures with their growth and bioluminescence. RZ can be used in physiologic studies of marine microorganisms.
5 DATA AVAILABILITY STATEMENT
All data generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
杂志排行

Journal of Oceanology and Limnology的其它文章
- Influence of sequential tropical cyclones on phytoplankton blooms in the northwestern South China Sea*
- Simulated perturbation in the sea-to-air flux of dimethylsulfide and the impact on polar climate
- Performance of ecological restoration in an impaired coral reef in the Wuzhizhou Island, Sanya, China*
- Investigating factors driving phytoplankton growth and grazing loss rates in waters around Peninsular Malaysia
- Effects of oxytetracycline dihydrate and sulfamethoxazole on Microcystis aeruginosa and Chlamydomonas microsphaera*
- Reproductive cycle of Ophiopholis mirabilis (Echinodermata: Ophiuroidea) in Zhangzi Island area, northern Yellow Sea*