川西云杉幼苗非结构性碳水化合物对土壤温度和水分变化的响应
2021-02-06申静霞李迈和于飞海贺云龙袁秀锦雷静品
申静霞,李迈和,于飞海,贺云龙,袁秀锦,雷静品,*
1 中国林业科学研究院林业研究所 国家林业局林木培育重点实验室,北京 100091 2 南京林业大学南方现代林业协同创新中心,南京 210037 3 中国科学院沈阳应用生态研究所,沈阳 110016 4 瑞士联邦森林、雪和景观研究院,瑞士苏黎世 CH- 8903 5 台州学院 浙江省植物进化生态与保护重点实验室,台州 318000 6 中国林业科学研究院森林生态环境与保护研究所,北京 100091
全球气候变化对陆地生态系统植物的生长和分布产生的影响不仅是学术研究的热点领域,而且作为全球性问题得到国际社会广泛关注[1-3]。高山地区由于其生态较为脆弱且垂直变化大,低温、干旱、强风和积雪等环境胁迫均有可能限制植物的生长发育[4],因此相较于陆地其它地区,高海拔地区植物对气候变化更为敏感[5-7],尤其是高山林线交错带,被认为是研究全球气候变化的理想场所[8-10]。温度[11-14]和水分[15-16]被认为是高山树木分布上限形成的关键因子。因此,研究林线树木生长和生理对温度与水分变化的响应对于预测林线的动态变迁及山地森林恢复具有重要意义[17-18]。
生长季的土壤低温与植物存活和生长密切相关,很可能是导致高山林线形成的关键因子[14,19],尤其是生长季的土壤低温与高山林线的形成密切相关[12-14]。Körner[19]提出土壤低温限制了细胞分化抑制了树木对碳水化合物的利用,从而抑制了树木的生长发育导致垂直分布上限的形成。碳水化合物是植物生长发育必需的物质,对生长季植物维持自身生长,冬季植物维持正常呼吸、防止细胞间结冰安全越冬以及来年早春植物的复苏发挥着至关重要作用[20-22]。高海拔树木长期生活在高寒恶劣的环境中,碳水化合物生产、储存及供应对高山树木的生存与生长发育尤为重要。针对土壤低温下林线树木碳水化合物储存、分配和利用的研究,形成了以碳水化合物为基础的两个主要假说:“碳受限”假说[3,23-28]和“生长受限”假说[5,11,19-22,29-31]。提出的这两个假说迄今为止没有统一的结论,另外,由于将地上和地下温度严格分离较为困难,因此加强土壤温度变化与植物碳水化合物关系的研究,有利于进一步对这两个假说进行验证。
水分同温度一样对植物的存活、生长及各项生理机能的正常运行也十分关键[15-16,32-34]。有研究表明,降水量随着海拔攀升而增加,并且由于气温和土壤温度降低,蒸发量变小且植物蒸腾速率显著降低,因此高海拔地区树木在生理上并不缺水[19,35]。但也有研究表明高海拔地区的树木仍处于生理缺水状态[16,36],这很可能是温度和水分共同作用的结果,低温限制了植物对水分的吸收及水分在树干中的转输[36-37],同时较低的蒸腾作用降低了水分拉力也限制了对水分的吸收利用[38]。因此,水分也可能是限制树木垂直分布的原因[5,15],并且研究温度与水分间的交互作用对高山树木生长和生理影响具有重要意义。
川西云杉(PiceabalfourianaRehd.et Wils)主要分布在我国西南地区海拔3300—4300m的森林垂直分布界限地带,是云杉属中分布海拔最高的树种,经常被选作高海拔森林更新荒山造林树种[39]。高山林线区严酷的环境可能并不足以导致树木死亡或生长受阻,却能强烈限制幼苗的存活和定居等森林更新过程[40-41],因此幼树能否成功定居对于高山林线的位置变化至关重要[42]。本研究以5年生川西云杉幼苗为研究材料,采用严格的室内控制实验,将土壤温度和土壤水分相结合,探究川西亚高山地区土壤温度和水分变化对高山树木碳水化合物生产、储存和供应生理过程的影响,以期验证川西云杉树线形成的“碳受限”假说和“生长受限”假说,完善对林线树木碳水化化合物的研究,并为全球气候变化下林线树木的生理生态响应和动态变迁研究提供基础数据。
1 材料与方法
1.1 植物材料与土壤采集
以四川省阿坝州金川县苗木基地(30°28′01″N,101°50′02″E)人工培育的5年生川西云杉幼苗(Piceabalfouriana)作为试验材料,试验用土取自该地区海拔3800m川西云杉160年天然林林下土壤,土壤为棕色针叶林土,pH值为5.92,土壤质量含水量为57%(土壤质量含水量=土壤水重量/干土重量×100%)。该树种分布区年平均气温7.8 ℃,生长季平均气温17 ℃;年平均降水量600—1000mm,6—9月降水量占全年降水量的80%,年平均相对湿度70%左右。于2016年6月初将川西云杉幼苗移栽至不透水的圆柱形塑料容器(直径10cm; 高20cm)中,移栽时土壤高度为15cm,并保持根系位置一致,为保持土壤温度和湿度在土壤表面撒约1cm厚的蛭石。
1.2 试验设计
选择135株长势良好且一致的川西云杉幼苗,于2016年6月上旬转移到3个环境条件完全相同的步入式人工气候室内,气候室内温度保持在17 ℃。每个气候室布设5个低温循环水箱(YHX-0510,宁波江南仪器厂,中国)控制土壤温度[43]。为检验Körner等[11,19]关于全球林线位置受生长季6.7 ℃左右土壤温度控制的假说,在整个试验期间设定从冰点以上2 ℃到7 ℃(6.7 ℃左右)到高于7 ℃的土壤温度梯度,即5个土壤温度处理:2,7,12,17,22 ℃。每个低温循环水箱中有9株川西云杉幼苗,连同培养容器被安置在水箱中,水箱中水的高度设置为高出培养容器中土壤表面1cm,用泡沫板封闭水箱,使土壤温度保持在5个温度(2、7、12、17和22 ℃)。每个水箱随机选择3株植物材料作干旱处理,3株植物材料作正常水分含量处理(对照处理),其余3株作饱和水分含量处理,分别用Dry、Normal和Wet表示。本试验共135株川西云杉幼苗[5个温度处理×3个水分处理×3个气候室×3株川西云杉幼苗],试验设计如图1。选择土壤温湿度测试计(SS101,Dowdon technology Co.,Ltd.,深圳,中国)指示的3个土壤湿度水平直接作为试验需要的3个土壤水分处理:干旱处理(土壤质量含水量为35%—40%)、正常水分含量处理(土壤质量含水量为55%—60%)、饱和水分含量处理(土壤质量含水量为75%—80%)。使用土壤温湿度测试计监测土壤温度和湿度,发现大概在浇水后的第4至5天土壤湿度会降低至设置范围以外,于是试验期间每隔5d的下午5:00进行浇水,使土壤湿度保持在我们设定的范围。
试验期间,气候室内光照时间14h,空气相对湿度70%,白天/夜间温度17/17 ℃。2016年10月上旬生长季末期,将全部川西云杉幼苗收获。
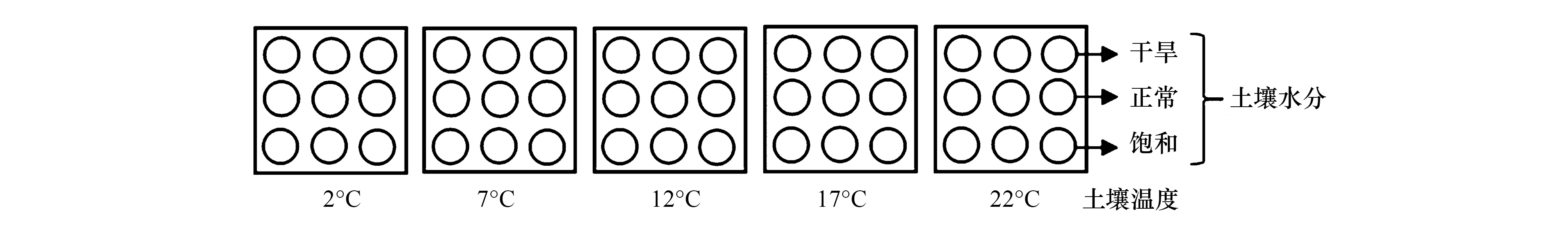
图1 试验设计简图Fig.1 Schematic diagram of experimental design
1.3 幼苗生长和生理指标测定
待收获后,将幼苗的各个器官分离(当年生叶、当年生枝、多年生叶、多年生枝、茎、根),放入烘箱于70 ℃烘至恒重,记录幼苗各器官干质量。
使用普通研磨仪(FW100,Test Instrument Co.,Ltd.,天津,中国)和球磨仪(MM400,Retsch,德国)将幼苗各器官分别研磨,然后通过孔径为0.3mm的标准分样筛,过筛后的植物样品用于可溶糖、淀粉和NSC浓度(可溶性糖浓度与淀粉浓度之和)的测定,含量为浓度与生物量的乘积。由于当年生叶和当年生枝的干物质含量过低,将同一个气候室内相同处理的3株幼苗的器官分别混合在一起研磨。
1.4 数据处理和分析
利用SPSS 22.0进行数据统计分析,以嵌套双因素方差(Two-way ANOVA)分析检验土壤温度、土壤水分、二者交互作用、气候室及同一个系统内的个体对生物量、可溶性糖、淀粉和NSC浓度的影响,差异显著性水平设定为0.05。同一个系统内的个体被作为随机因素嵌套在土壤水分处理中。当检测到土壤温度与土壤水分的交互作用有显著影响时,用Bonferroni校正进行事后检验,比较5个土壤温度处理下土壤水分处理之间生长和生理特性的差异以及同一土壤水分处理下5个土壤温度处理对植物生长和生理影响的差异。分析前,对所有数据进行正态性和方差同质性检验。利用SigmaPlot 12.5进行柱状图的绘制。
2 结果与分析
2.1 土壤温度和土壤水分对生物量的影响
土壤温度对茎生物量和根冠比影响显著,对总生物量和其它部分生物量均无显著影响(表1)。在正常水分含量处理下,土壤温度对根生物量和根冠比有显著影响(图2),根生物量和根冠比随土壤温度降低显著降低,可能是由于低温会降低根系对土壤养分和水分的吸收速率,导致根的生长受到抑制。土壤水分对总生物量、多年生叶生物量、根生物量和根冠比影响显著(表1)。干旱处理显著降低了12 ℃时的总生物量和茎生物量;干旱处理和饱和水分含量处理对根生物量和根冠比有较显著的影响(图2),干旱处理和饱和水分含量处理显著降低了17和22 ℃时的根生物量和根冠比,同时也显著降低了12 ℃时的根生物量。土壤温度和土壤水分对根冠比的影响存在显著的交互作用(表1),在不同的土壤温度处理下,干旱和饱和水分含量处理对根冠比的影响有不同表现,在12和22 ℃时显著降低了根冠比,而随着温度降低影响不再显著;土壤温度降低显著减小了正常水分含量处理下的根冠比,而对干旱和饱和水分含量处理下的根冠比无显著影响(图2)。
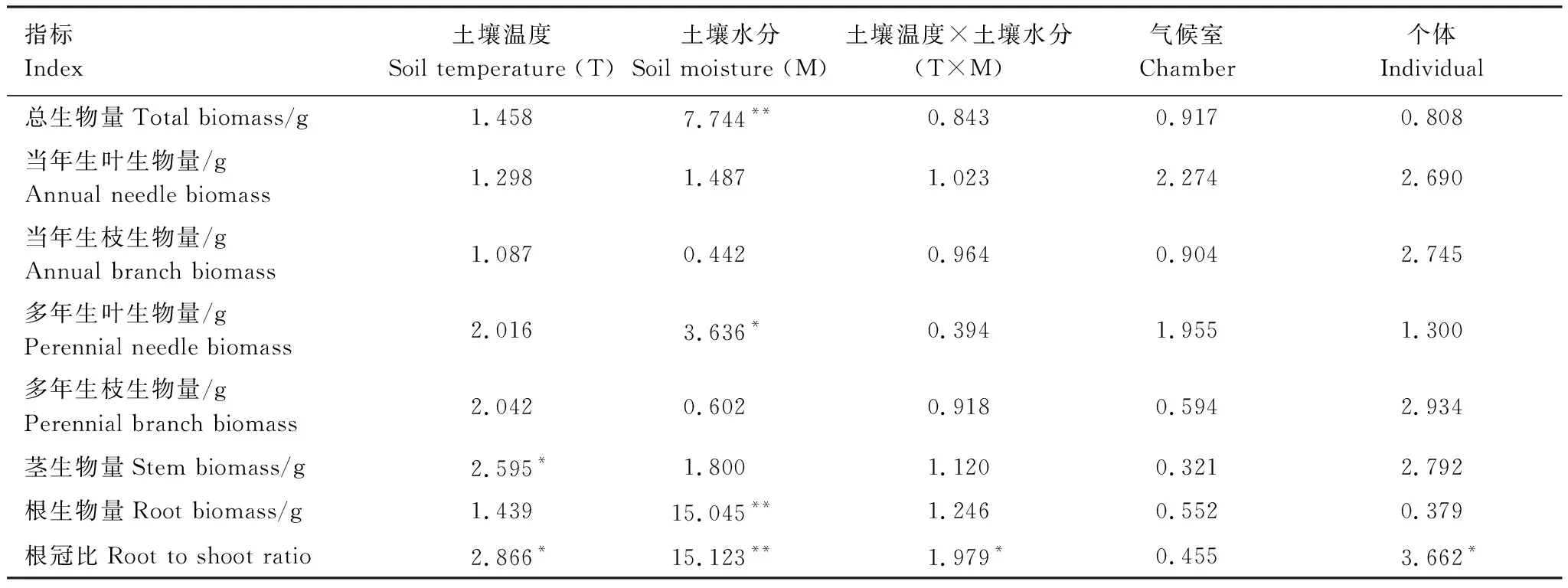
表1 土壤温度和土壤水分对幼苗各器官生物量的影响 (F)

图2 不同土壤温度和水分处理下川西云杉幼苗各器官生物量及分配(平均值±标准误差)Fig.2 Biomass and root to shoot ratio of Picea balfouriana seedlings under different soil temperature and moisture treatments at the end of the experiment不同大写字母表示不同土壤水分处理间差异显著;不同小写字母表示不同土壤温度处理间差异显著(P<0.05)
2.2 土壤温度和土壤水分对非结构性碳水化合物的影响
土壤温度对当年生叶中NSC浓度、淀粉浓度以及可溶性糖/淀粉比值影响显著,但对可溶性糖浓度无显著影响(表2)。正常水分含量处理下,当年生叶中淀粉和NSC浓度随土壤温度降低显著升高(图3);饱和水分含量处理下,淀粉浓度在22 ℃时显著低于其它4个温度处理,且这4个温度处理间无显著差异,NSC浓度在22 ℃时显著低于7和12 ℃(图3);干旱处理下,低温显著增大了可溶性糖/淀粉比值;饱和水分含量处理下,可溶性糖/淀粉比值在22 ℃时显著高于其它4个温度处理,且4个温度处理间无显著差异(图4)。土壤温度对茎和根中可溶性糖、淀粉和NSC浓度影响显著(表2)。在正常水分含量处理下,茎中可溶性糖浓度在7 ℃显著高于17 ℃;在干旱处理下,茎中可溶性糖和NSC浓度在7 ℃时最高,且显著高于17和22 ℃(图3)。土壤温度对根中可溶性糖、淀粉和NSC浓度均有显著影响(表2)。在正常水分含量处理下,随着土壤温度降低,根中可溶性糖浓度呈显著增高趋势,可溶性糖浓度在2和7 ℃显著高于17 ℃;在饱和水分含量处理下,根中可溶性糖浓度在7 ℃最高,且显著高于22 ℃(图3);正常水分含量处理下,根中淀粉浓度在12 ℃最高,且显著高于22 ℃;干旱处理下,随着土壤温度降低,根中淀粉和NSC浓度有增大趋势,在7 ℃时显著高于17 ℃(图3)。正常水分含量处理下,根中NSC浓度在2和12 ℃显著高于22 ℃;饱和水分含量处理下,根中NSC浓度在7 ℃时最高,显著高于2和22 ℃(图3)。因此,土壤低温并没有导致碳受限,反而显著增大了当年生叶和根中NSC浓度,这可能是由于在相对较低的温度下,植物会增加NSC浓度来避免细胞间和细胞内结冰。
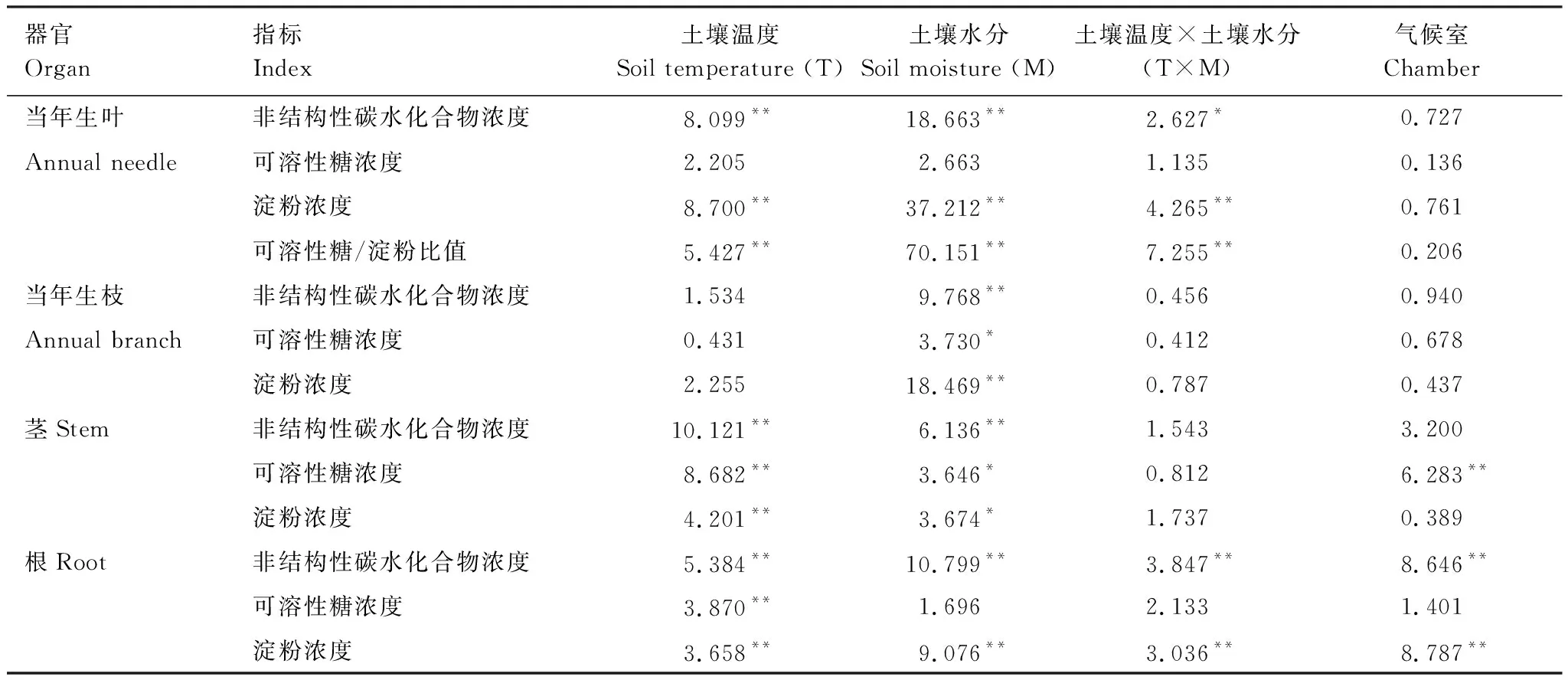
表2 土壤温度和土壤水分对各器官可溶性糖、淀粉及NSC浓度的影响 (F)/%

图3 不同土壤温度和水分处理下川西云杉幼苗各器官可溶性糖、淀粉和NSC浓度Fig.3 Soluble sugar,starch and NSC concentration in different parts of Picea balfouriana seedlings under different soil temperature and moisture treatments at the end of the experiment不同大写字母表示不同土壤水分处理间差异显著;不同小写字母表示不同土壤温度处理间差异显著(P<0.05)
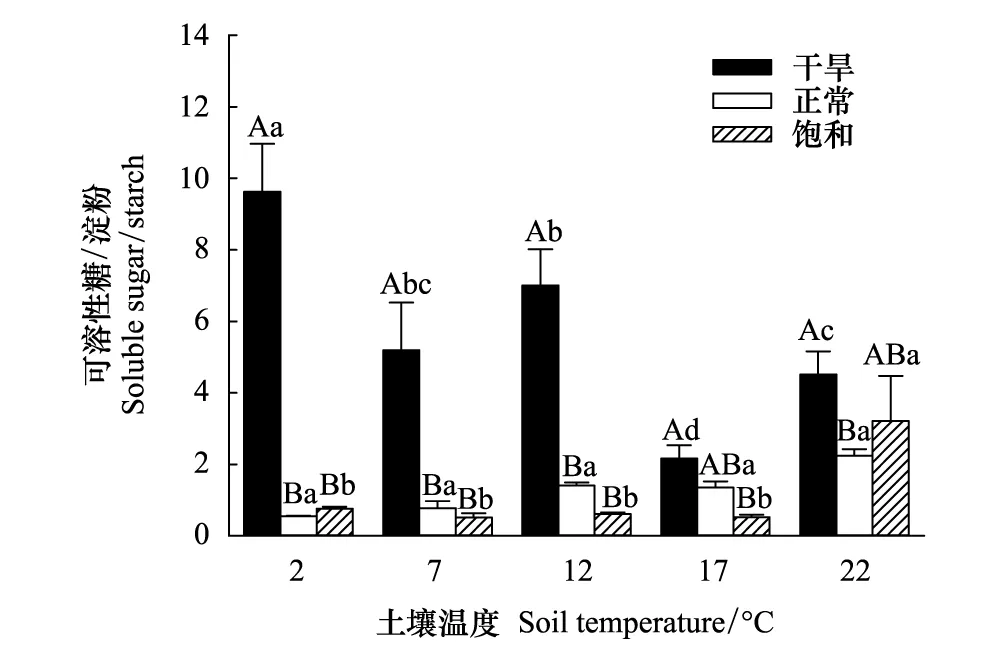
图4 不同土壤温度和水分处理下川西云杉幼苗当年生叶中可溶性糖与淀粉的比值Fig.4 The ratio of soluble sugar to starch in annual needles of Picea balfouriana seedlings under different soil temperature and moisture treatments at the end of the experiment不同大写字母表示不同土壤水分处理间差异显著;不同小写字母表示不同土壤温度处理间差异显著(P<0.05)
土壤水分对当年生叶中淀粉浓度、NSC浓度和可溶性糖/淀粉比值影响显著(表2)。在2和7 ℃时,干旱显著降低了当年生叶淀粉和NSC浓度;饱和水分含量处理显著增大了17 ℃时当年生叶中淀粉和NSC浓度,同时显著增大了12 ℃时当年生叶中淀粉浓度(图3);在2,7,12和22 ℃时,干旱显著增大了可溶性糖/淀粉比值(图4)。土壤水分对当年生枝中可溶性糖、淀粉和NSC浓度均有显著影响(表2)。在2 ℃时,干旱处理下当年生枝中淀粉和NSC浓度显著低于饱和水分含量处理;在7 ℃时,淀粉浓度在干旱处理下也显著低于饱和水分含量处理(图3)。土壤水分对茎中可溶性糖、淀粉和NSC浓度影响显著(表2),干旱显著降低了22 ℃时茎中NSC浓度(图3)。土壤水分对根中淀粉和NSC浓度影响显著(表2),在2,12和17 ℃时,干旱处理显著降低了根中淀粉和NSC浓度,在2 ℃时,水分饱和含量处理也同样显著降低了根中淀粉和NSC浓度(图3)。由以上结果可知,干旱对各器官NSC浓度的负面影响较大,尤其是对当年生叶中淀粉和NSC浓度影响最为显著。
土壤温度和水分对当年生叶和根中淀粉浓度、NSC浓度和可溶性糖/淀粉比值的影响存在显著的交互作用(表2),不同水分处理下,土壤温度的影响有不同表现。
3 讨论
在本研究中,土壤温度降低对川西云杉幼苗总生物量无显著影响,但显著降低了根生物量和根冠比,这与Domisch等[44]和Lahti等[45]的研究结果是一致的,可能是由于在低温环境下,低温会严重制约根系对土壤养分和水分的吸收速率,导致根的生长受到抑制[36]。刘新圣等[46]研究也发现随着海拔的升高,为了应对低温胁迫植物会将更多的生物量分配给光合器官,非光合器官的生物量相对减小。但也有研究表明土壤温度降低导致生长速率降低,植物各器官的生物量均会呈现出显著减小的趋势[47-48]。本研究结果表明,干旱和饱和水分胁迫显著降低了根生物量和根冠比,且在土壤温度较高时影响显著。但有研究表明,幼苗的根冠比随着土壤含水量的降低显著提高[33,49],植物受到干旱胁迫时,根系首先做出感应,根比重增加,在严重土壤水分亏缺条件下幼树光合产物倾向于向地下分布转移,增大根系竞争水分和营养成分的能力[49-50]。而本研究结果与此恰相反,很可能是水分和温度共同作用的结果。此外,本研究发现土壤温度与水分胁迫存在明显的交互作用,随着土壤温度的降低,水分胁迫对根生物量和根冠比的影响由显著变得不再显著,这一结果表明,此时低温对植物的影响要远大于水分。
本研究结果表明,川西云杉幼苗叶片中NSC浓度随着土壤温度的降低并没有降低,反而有显著升高的趋势,这可能是由于在相对较低的温度下,植物会增加NSC浓度来避免细胞间和细胞内结冰[21-22],但NSC浓度随着温度的降低之所以呈现出升高的趋势也可能是由于植物的生长在低温条件下受到了抑制,因此产生了浓缩效应,但在本研究中,随着温度降低川西云杉的生长并未受到显著抑制,且当年生叶的生物量也未发生显著变化,即土壤温度降低对NSC含量也无显著影响,因此随着土壤温度降低川西云杉并没有表现出碳受限,这也从侧面支持了“生长受限”假说。这一结果说明,随着土壤温度降低,叶片仍能维持正常水平的光合作用或者仍能保持一定水平的光合产量。对可溶性糖/淀粉比值的分析发现,正常水分条件下,土壤温度对可溶性糖/淀粉比值无显著影响,但以往的研究表明,可溶性糖/淀粉比值在植物抵御低温胁迫中发挥着重要作用[51],生长在高海拔的树木不仅依赖于NSC,而且需要足够高的可溶性糖/淀粉比值才能顺利越冬[52],植物组织中的可溶性糖/淀粉比值随温度降低会呈现出显著升高的趋势[31,53],休眠期的树木为抵御严寒,组织中的可溶性糖/淀粉比值也显著高于生长季[3]。
干旱对碳同化、碳利用和碳储存过程均存在负面影响[54],研究表明干旱胁迫对NSC浓度影响显著,干旱显著降低了当年生叶中NSC浓度,这可能是由于干旱胁迫下植物气孔关闭限制了植物的光合作用,导致光合产物减少,长期下去植物开始调动和利用植物体内储存的可溶性碳水化合物维持植物生长[33,55],但本研究还发现土壤低温显著加重了干旱胁迫对植物NSC浓度的影响,这与Adams等[56]的研究结果是相反的,Adams研究发现,在相同的干旱胁迫条件下,植物在较高的温度对碳水化合物有更大的消耗,同时呼吸作用增强也会加剧对碳水化合物的消耗,而温度较低时,植物本身对碳的需求减少,呼吸作用也相对减弱,因此干旱胁迫下可溶性碳水化合物会呈现出随着温度升高而降低的趋势。本研究发现,土壤低温下,干旱显著提高了当年生叶中可溶性糖/淀粉比值,这意味着适度的干旱可能会有利于川西云杉幼苗抵御低温胁迫,但是在低温条件下,干旱也显著降低了当年生叶中NSC浓度,即很可能会导致植物的碳饥饿,不利于植物抵御低温胁迫,土壤低温和干旱对川西云杉非结构性碳水化合物影响的交互作用有待进一步研究。
4 结论
土壤温度对川西云杉幼苗地上和地下生物量的分配有显著影响,土壤低温显著降低了根冠比。川西云杉幼苗在土壤低温下并未表现出碳受限,NSC浓度反而有显著升高的趋势,这也从侧面支持了林线形成的“生长抑制”假说。这一结果说明,川西云杉幼苗在土壤低温下仍然能维持正常水平的光合作用或者叶片仍能保持一定水平的光合产量。此外,土壤温度和水分对非结构性碳水化合物的影响存在显著的交互作用,干旱在低温处理下对川西云杉幼苗的NSC浓度有显著影响,因此在土壤低温下干旱很可能会导致川西云杉的“碳饥饿”。这些结果表明未来在模拟全球气候变化影响的模型中应该考虑土壤温度的影响,以便更准确地预测全球气候变化的影响。
由于川西云杉幼苗针叶较粗硬,为光合指标的测定带来了困难,因此建议下一步着重将光合作用与养分元素、碳水化合物三者结合起来进行探究,以期完善我们对高山树木碳水化合物供应与积累生理过程的理解。
