山西省小麦苗期根系性状及抗旱特性分析
2021-02-05赵佳佳武棒棒乔麟轶张树伟闫素仙郑兴卫
赵佳佳 乔 玲 武棒棒 葛 川 乔麟轶 张树伟闫素仙 郑兴卫,* 郑 军,*
山西省小麦苗期根系性状及抗旱特性分析
赵佳佳1,2,**乔 玲2,**武棒棒2葛 川1乔麟轶1张树伟1闫素仙2郑兴卫2,*郑 军2,*
1山西农业大学有机旱作农业研究院 / 有机旱作山西省重点实验室, 山西太原 030006;2山西农业大学小麦研究所, 山西临汾 041000
小麦苗期根系形态是成株期根系分布的基础, 与抗逆和产量密切相关, 全面认识苗期根系及抗旱特性, 对于抗旱优异种质的利用和早期筛选具有重要意义。采用239份山西省小麦品种(系)在土培条件下, 研究了苗期根系性状及对水分胁迫的响应。结果表明, 正常生长下山西小麦苗期根系性状多样性丰富, 地方种变异最大; 不同年代品种中, 除最大根长随年代略下降外, 其他性状均呈先升后降的趋势; 不同根系性状对水分胁迫响应存在差异, 总根长对水分最敏感, 其次为根表面积、根体积和根生物量, 最大根长和平均根数不敏感。苗期根系综合抗旱能力随年代呈先降后升的趋势, 地方种和20世纪70年代品种多为中抗, 80和90年代的品种抗旱性较低, 2000年以后审定品种的抗性较高, 其中旱地品种抗性最好。苗期根系抗旱特性与产量性状相关分析发现, 最大根长、总根长、根体积和根生物量与雨养条件下的千粒重和产量显著正相关, 最大根长和根生物量与成株期抗旱性也显著正相关。因此苗期最大根长和根生物量可作为半干旱地区旱地育种过程中抗旱性和产量的早期筛选指标。
山西小麦; 苗期根系; 抗旱特性; 演变趋势
根系具有固着植物, 吸收、贮藏、运输水分和营养物质的功能。对于小麦而言, 根系形态和性能直接影响地上部分的生长、发育和产量表现, 是小麦具有节水、抗倒、耐热、落黄等良好特性的根本所在[1-3]。成株期根系发育易受到土壤和栽培措施影响, 常规取样破坏性大、工程复杂, 使得表型调查难以高效实施。目前, 小麦根系形态的研究方法主要有挖掘法、塑管土柱法、网袋法、根室观察法、三维坐标容器法、同位素示踪法和微根管系统等[3-4]。这些方法虽能直接或间接反映成株期根系生长状态, 但极少用于大规模群体取样, 导致小麦遗传改良研究主要关注在地上部分的重要农艺性状方面, 对“隐藏的另一半”——根系的研究相对欠缺。因此, 寻找准确、便捷的根系形态检测方法一直倍受重视。
小麦初生根是指种子萌发出1条胚根和另外4~6条种子根, 对根系建成和后期生长发育具有重要作用[5-7]。早期研究已证实发育良好的初生根有助于促进次生根的建成[8], 苗期根系形态可有效指示成株期的根群分布及特点[9-11]。此外, 初生根数量和活力对小麦产量也具有重要贡献, Xie等[12]发现苗期总根长和初生根数目与穗粒数、千粒重和地上生物量正相关; 初生根发育的深层根系可利用深土层储水, 直接关系到成株期抗旱性和产量[13-14]。An等[15]和Fan等[16]发现控制苗期根系的QTL同时调控营养吸收和抗逆; 且有些调控苗期根系生长的基因在干旱胁迫下高表达[5]。可见苗期根系发育与小麦的抗逆和产量密切相关。目前苗期根系性状调查常采用营养液法、凝胶室法、纸培法、砂培法和土培法等[17-21]。这些方法误差小、操作简便、通量高、损伤小, 取样后结合图像分析, 可完成对苗期根系形态的快速检测。肖永贵等[22]研究了骨干亲本‘京411’及14个衍生品种的苗期根系, 发现衍生品种的最大根长和根系生物量较京411显著改良; 景蕊莲等[23]对35个不同栽培类型的小麦品种(系)苗期根系与抗旱性关系研究, 发现中间型根系的品种反复干旱处理后存活率高, 抗旱性强; 马富举等[24]利用2个耐旱性不同的品种, 发现干旱胁迫下苗期根系通过较高的生物量和根长增加对水分的吸收; Dhanda等[25]利用30个品种探讨了早期抗旱评价指标, 认为根长和根冠比可作为预测耐旱的性状指标。用遗传群体研究苗期根系也有报道, Landjeva等[26]利用重组自交系群体检测根系性状与抗旱性的关系, 认为根长可作为苗期抗旱的指标; 周晓果等[27]利用双单倍体群体发现苗期根数和根干重与抗旱性显著相关。可见, 小麦苗期根系与抗旱性高度相关已受到普遍认可。我国西南、西北、黄土高原和长江中下游等麦区的小麦根系研究也有报道[28-31], 但大多通过遗传群体和个别品种进行, 并未建立起可用于抗旱育种的苗期筛选指标。因此, 系统研究干旱胁迫下苗期根系的形态特征和生理功能亟待进行。
山西省小麦种植历史悠久, 目前出土距今已有3000多年的小麦籽粒, 相同年代的面饼也有发现[32]。山西地理位置独特, 位于黄土高原东部, 属温带季风性气候, 北邻内蒙古草原, 南接中原腹地, 东依太行, 西界黄河, 外缘有山脉环绕, 不受海风气候的影响; 由于内蒙古冬季冷气团形成的大陆性季风性气候, 自南向北温差较大。全省年降水量在400~ 650 mm, 小麦种植区域为典型的半干旱地区, 使得山西小麦历来以抗旱闻名全国, 如晋麦33、长6878和晋麦47等品种被选为国家旱地小麦区试对照品种, 其中晋麦47从20世纪90年代至今一直作为黄淮北片旱地、山西南部旱地和陕西渭北旱地区试对照[33]。以山西省地方种和不同年代育成品种为材料探究半干旱地区的小麦苗期根系及抗旱特性具有较强的代表性。本文系统研究了山西省小麦品种(系)苗期根系性状及其对干旱胁迫响应, 鉴选出抗旱性评价的根系指标, 研究结果有助于半干旱地区小麦抗旱遗传改良。
1 材料与方法
1.1 试验材料
山西省小麦品种(系)共239份, 包括建国以来山西省育成品种198份, 占审定品种总数的90%, 其中旱地品种96个, 水地品种102个; 地方农家种41份, 为中国小麦核心种质中来源于山西省的地方种, 材料信息见附表1。育成品种来源于各育种单位和本课题保存, 地方种为中国农业科学院作物科学研究所郝晨阳副研究员提供。
所有材料于2017—2018年度和2018—2019年度种植于小麦研究所试验基地 (山西省临汾市36°2'N, 111°18'E), 10月中旬播种, 翌年6月中旬收获。每个材料播种2行, 行长1.5 m, 每行30粒。设旱地和水地2种生长条件, 旱地为雨养条件(生育期平均降雨量约为194 mm, 图1); 水地于越冬期、拔节期和灌浆期地下水灌溉, 灌溉量均为700 m3hm–2(70 mm)。成熟后各品种随机选取5株, 测量株高、小穗数、穗粒数和千粒重等性状。2个生育期内自然降水和常年相近, 未发生极端天气和严重自然灾害, 小麦生长情况良好。
1.2 苗期培养及干旱胁迫处理
挑选籽粒饱满均一、无病虫害的种子, 腹沟朝下均匀的摆放在培养皿中, 用蒸馏水浸泡吸胀萌发; 种子露白后, 选取6粒萌发程度一致的种子, 点播在含有蛭石和100 mL水的培养杯中(直径为7 cm, 高8 cm), 于光强为500 µmol m−2s−1, 湿度为70%, 光温周期为20℃ 16 h/16℃ 8 h的人工气候室培养。设正常水分和19.2% PEG-6000 (polyethylene glycol, PEG)模拟干旱胁迫2种方式, 各设3次重复, 共6个处理。每天相同时间点播1个处理的材料(239份), 各处理出苗7 d后进行正常补水/胁迫处理; 继续生长4 d后从土壤中收集根系完整的植株, 清洗后, 分别测定根系形态。
1.3 根部性状测定
根状扫描仪(Microtek Scanmakeri 800 plus)扫描幼苗根部图片, 根数(average number of roots, RN)、总根长(total root length, TRL)、根表面积(root surface area, RSA)和根体积(root volume, RV)采用Win-RHIZO系统(Regent Instruments Inc., Quebec, Canada)测量获得, ImageJ软件测量最大根长(MRL), 每个品种测定5株。幼苗自根茎相接处分开, 称量地上部和地下部鲜重(shoot fresh weight, SFW和root fresh weight, RFW), 再分别放在烘箱105℃杀青20 min, 70℃烘干至恒重后, 称量地上部和地下部干重(shoot dry weight, SDW和root dry weight, RDW), 计算根冠比(root/shoot ratio, RSR)和地上、地下含水量(water content of shoot, SWC和water content of root, RWC), 每个品种测定5株取平均值。
露点渗透压仪(Wescor 5600, 美国)测定叶片及根部的渗透摩尔浓度, 按照公式w = –ic×R×T计算渗透势, R是气体常数, 通常取值为0.008314; T为凯氏温度, T = 273+t, t为室温(摄氏度); ic是被测样品的渗透摩尔浓度(mmol kg–1)。
1.4 抗旱性评价方法
参照文献[34]计算抗旱系数(drought-tolerance coefficient, DTC)、隶属函数、综合抗旱评价值()等指标, 抗旱系数为处理与对照的相对值, 公式为:
DTC=X()/CX()
式中,X()和CX()分别表示处理和对照性状测定值。
对所有性状DTC进行主成分分析, 再计算其隶属函数值(X), 公式为:
(X) = [CI− CImin]/[CImax− CImin]
式中,(X)表示第个主成分的隶属函数值, CImin表示所有材料该主成分最小值, CImax表示所有材料该主成分的最大值。
各综合指标权重W:
W=P/∑(= 1)P(= 1, 2, …,)
式中,W表示第个主成分在所有主成分中的重要程度,P表示各材料第个综合指标贡献率,为主成分数量。
D为综合抗旱能力评价值,D越高, 表示该材料的综合抗旱能力越大。
D= ∑(= 1)(X)W(= 1, 2, …,)
为了综合考虑苗期根系特性, 对根系性状进行归一化处理, 与性状相对抗旱系数结合评价苗期的抗旱性。
X′() = [X() −min]/[max−min]
DTC′ =X′() DTC
X′()为数据归一化处理结果, DTC′为苗期根系抗旱特征值。
1.5 数据分析
用Microsoft Excel 2010软件对数据进行常规统计分析, SPSS 24.0软件进行方差、相关性和主成分分析。
2 结果与分析
2.1 小麦苗期根系性状变异
正常水分条件下, 山西小麦苗期根部性状总体变异幅度较大, 除地上和地下含水量(SWC和RWC)的变异系数较小外, 其他性状均在11%以上, 其中总根长(TRL)、根系表面积(RSA)、根体积(RV)和根干、鲜重(RFW, RDW)等性状变异系数在17%以上, 说明山西小麦苗期根系表型丰富, 品种间根系性状差异明显(图2-A和表1)。品种类型中以育成品种的根系性状值较大, 除旱地品种地上干、鲜量(SFW, SDW)变异系数较大外, 其他性状均以地方种变异丰富; 育成品种中旱地品种根系变异丰富, 但水地品种根系性状值较大。
2.2 干旱胁迫下根系性状变异
PEG处理抑制了苗期根系生长, 有些品种敏感, 胁迫前后根系差异大, 如长麦6973; 有些品种胁迫前后根系差异小, 如运旱21-30 (图2-B)。胁迫条件下苗期根系形态性状整体受到抑制, 其中TRL的降幅最大, 为33.94%~39.37%, 其余依次为RSA、RV、最大根长(TRL)、RFW和RN (表2); 各性状变异系数变小, 表明缺水条件下根系形态性状变异性低。采用抗旱系数(DTC)分析不同性状的相对抗性表明, 山西小麦苗期根系性状的DTC变异丰富(图3), 不同性状对胁迫的响应存在差异, 总根长的抗旱系数大值为1.5; 除根冠比外, 其他性状的抗旱系数也较低, 多分布在1.0以下。胁迫下根系生物量(RFW和RDW)和地上生物量(SFW和SDW)均降低, 但SFW和SDW降幅大, 根冠比(FRSR和DRSR)增大, 抗旱系数分布在1.0~1.5之间, 表明干旱胁迫对地上部的抑制作用大于根系。整体而言, TRL、RSR、SFW和SDW对水分胁迫最敏感, 其次为RSA、RV、RFW和RDW, MRL和RN对水分敏感性低(表2和图3)。
A: 正常条件下小麦苗期根系多样性。1: 晋春16; 2: 玉兰麦; 3: 晋麦79; 4: 临优145; 5: 晋麦73; 6: 晋麦86。B: 正常(CK)和胁迫(PEG)处理下苗期根系表型, YH 21-30: 运旱21-30; CM 6973: 长麦6973。
A: diversity of seedling root with five plants under control in wheat; 1: Jinchun 16; 2: Yulanmai; 3: Jinmai 79; 4: Linyou 145; 5: Jinmai 73; 6: Jinmai 86. B: difference of root traits at seedling stage between control and drought stress. YH 21-30: Yunhan 21-30; CM 6973: Changmai 6973.

表1 正常水分条件下山西苗期小麦根部性状分析
MRL: maximum root length; TRL: total root length; RSA: root surface-area; RV: root volume; RN: root number; SFW: shoot fresh weight; SDW: shoot dry weight; RFW: root fresh weight; RDW: root dry weight; FRSR: fresh root/shoot ratio; DRSR: dry root/shoot ratio; SWC: water content of shoot; RWC: water content of root; CV: coefficient of variation.
地方种地上部受抑制程度大于育成品种, 地上鲜、干重的抗旱系数(DTCSFW和DTCSDW)小, 根系生物量和根冠比的抗旱系数(DTCRFW、DTCRDW和DTCFRSR、DTCDRSR)则表现为地方种大于育成品种; 胁迫下旱地品种的TRL、MRL、RSA和RDW降幅较小, 抗旱系数除DTCRN外, 其他性状的DTC以旱地品种最大, 地方种和水地品种较小。整体而言, 旱地品种根系对干旱胁迫的敏感性低(表2和图3)。
2.3 小麦苗期根系性状演变趋势
对农艺性状演变趋势分析, 可以归纳出品种适应自然和人工选择的规律。山西小麦苗期根系性状随年代变化有3种趋势, 其中MRL随年代略降, 以地方种最高; TRL、RSA、RV、RN及RFW、RDW呈先增后降趋势, 以20世纪80和90年代品种最大; FRSR和DRSR随年代变化不明显(图4)。各性状抗旱系数随年代变化趋势与根系性状演变不同, DTCMRL和DTCRDW随年代缓慢增大; DTCTRL、DTCRSA、DTCRV、DTCRFW、DTCRFW和DTCRDW先降后增。整体上随着人工选择与品种改良, 地方种到育成品种根系性状先增后降, 80、90年代根系性状最大, 90年代之后随着育种水平的提高, 抗旱能力不断提高。
2.4 苗期抗旱性综合评价
抗旱性受多因素控制, 利用单一指标鉴定品种抗旱性存在局限性, 将较多指标转换为综合抗旱值评价抗旱性已得到认可。山西小麦苗期值变化范围为0.273~0.803, 可分为高抗(≥ 0.611)、中抗(0.566 ≤< 0.611)、一般(0.477 ≤< 0.566)、较敏感(0.432 ≤< 0.477)和敏感(< 0.432) 5级。各级材料份数依次为38、36、80、46和39份, 分别占总数的15.89%、15.06%、33.47%、19.25%和16.32%。高抗品种包括: 长麦6135、晋麦88、长6878和运旱21-30等旱地品种20份, 长6452、临优2018、晋太114和晋麦74等水地品种11份, 以及白秃麦、齐穗麦和绛州红等地方种7份。
各年代均以旱地品种的值较高, 抗旱性最好, 20世纪80和90年代的品种抗旱能力变弱, 大多为一般和较敏感类型, 地方种和20世纪70年代的品种抗性一般, 2000年以后审定品种的抗旱性整体表现较好,值主要分布在中抗和高抗之间(图5), 表明随着育种水平的不断提高, 根系抵抗旱胁迫的能力增强, 能更好的适应缺水环境。
2.5 渗透势监测
干旱环境下, 保持良好的水分状态是植物抗旱的主要方式, 监测胁迫前后渗透势变化可有效反映出植株在干旱条件下的渗透调节能力[35-36]。依据苗期值分级, 随机选取抗旱型和敏感型的小麦品种各5份, 测定正常和胁迫下叶片和根系渗透势, 其中晋麦88、晋麦82、晋麦83、运旱21-30和晋麦90等品种抗性好, 晋春16、晋春14、运旱20410、长麦5973和玉兰麦等抗性较差。与正常生长相比, 干旱胁迫下各品种的叶片和根系渗透势均降低, 根系的降幅较叶片小; 抗旱性好的品种根系和叶片的渗透势下降程度大于抗旱性弱的品种, 渗透调节能力较强(图6)。此外, 胁迫处理下根系与叶片的渗透势差增大, 地上部分对干旱胁迫较根系更敏感。
2.6 苗期根系与产量性状的关系
评价粮食作物抗旱性要兼顾丰产性, 通过水旱地产量性状计算抗旱系数求得成株期值。苗期根系性状抗旱特性与成株期产量相关性分析表明, 苗期DTC′MRL、DTC′SFW、DTC′SDW、DTC′RFW和DTC′RDW与成株期值显著正相关, 其中DTC′SDW和DTC′RDW与成株期值极显著相关, DTC′RN与成株期值显著负相关(表3)。DTC′MRL与雨养条件下千粒重和产量显著正相关, 与灌溉下穗粒数显著负相关; DTC′TRL、DTC′RSA、DTC′RV、DTC′RFW和DTC′RDW均与雨养和灌溉条件下千粒重显著相关, 仅与雨养条件下产量显著相关(表3)。说明苗期根系性状抗旱性与雨养条件下产量性状的相关性较高, 可以将苗期最大根长和根生物量作为旱地育种工作中筛选产量和抗旱性的指标。
缩略同表1。Abbreviations are the same as those given in Table 1.
3 讨论
3.1 苗期根系性状与抗旱性密切相关
根系是作物吸收水分和养分的主要器官, 形态特征直接影响水分的吸收和利用。苗期根系遗传力高, 受环境影响较小, 正常生长条件下可以反映成株期根系形态和根群分布, 与成株期抗旱性息息相关[5,9-13]。玉米的苗期根长、根重和根系水导等与抗旱性极显著相关, 可作为生产中筛选抗旱品种的指标[37]。王贺正等[38]证明水稻根长也可作为苗期抗旱性的形态指标; 旱育秧苗一直强化育根, 苗期壮根可保证生育后期单位株高干物质重[39], 因此, 研究旱胁迫下苗期根系的形态特征和生理功能是揭示种质抗旱性的有效方法。小麦是干旱与半干旱地区的主要作物, 发达的早期根系有利于干旱环境下水分和养分的吸收[5]。Dhanda等[26]发现小麦幼苗干旱胁迫下, 根长、根冠比和膜渗透性表现出较大变异和较高遗传力, 可用于预测植株的耐旱性。本文利用PEG模拟旱胁迫探索小麦苗期根系性状的响应, 发现总根长(TRL)、根冠比(FRSR和DRSR)和地上部生物量(SFW和SDW)对水分胁迫敏感, 其次为根表面积(RSA)、根体积(RV)和根生物量(RFW和RDW), 最大根长(MRL)和平均根数(RN)对水分胁迫敏感性较低。目前本文考察了不同品种根系对水分胁迫的形态响应, 后期将从遗传途径和吸水效率进行研究, 挖掘调控根系性状的显著位点, 进一步解析小麦根系抗旱特性的机理。
A: 叶片渗透势; B: 根系渗透势。JM88: 晋麦88; JM83: 晋麦83; JM82: 晋麦82; YH 21-30: 运旱21-30; JM90: 晋麦90; JC16: 晋春16; JC14: 晋春14; YH 20410: 运旱20410; CM 5973: 长麦5973; Yulanmai: 玉兰麦。
A: osmotic potential of leaf; B: osmotic potential of root. JM88: Jinmai 88; JM83: Jinmai 83; JM82: Jinmai 82; YH 21-30: Yunhan 21-30; JM90: Jinmai 90; JC16: Jinchun 16; JC14: Jinchun 14; YH 20410: Yunhan 20410; CM 5973: Changmai 5973.

表3 苗期根部性状抗旱性与产量相关性分析
**与*分别表示在 0.01和0.05水平差异显著。缩写同表1。
**and*indicate different significances at the 0.01 and 0.05 probability levels, respectively. Abbreviations in the first column are the same as those in Table 2. PH: plant weight; GNPS: grain number per spike; SNPP: spikelet number per spike; EPT: effective plant tiller; TKW: 1000-kernel weight; YPP: yield per plant. Abbreviations the same as those in Table 1.
3.2 苗期根系形态影响产量性状
苗期初生根在整个生育期均发挥重要作用, 形成的深层根系在干旱环境下能有效利用深土层养分和水分[13,40], 进而影响产量性状。半干旱地区小麦苗期多处于干旱少雨的秋冬季节[41], 土壤下层充足的水分能保障初生根的生长, 水分不足则会减少次生根的生成, 影响根系分布。此外, 苗期根系活力直接关系到成株期地上部的生长发育[5-6,42], 这是小麦苗期的根系形态建成对产量贡献的关键[43]。目前苗期主要用反复干旱存活率或PEG模拟胁迫法鉴定种质的抗旱性, 这类方法能体现苗期生理上抗旱性, 也是苗期高抗品种含有水地品种的原因。在农业生产中, 作物抗旱性的评价一般以稳产为前提, 牺牲产量提高存活力的种质是不可取的, 人为设定的成株期抗旱性评价指标往往重点关注产量。李龙等[44]发现‘晋麦47’等旱地高产稳产品种的苗期抗旱性一般, 得出苗期抗旱性与成株期抗旱性不相关结论, 可能是苗期反复干旱法仅考虑了胁迫下存活率, 忽略了根系在生长发育后期对籽粒形成的贡献。本文分析了苗期根系性状归一化后的抗旱系数与产量性状的关系, 发现最大根长(MRL)、总根长(TRL)、根体积(RV)和根生物量(RFW和RDW)的抗旱特性与雨养下千粒重和产量呈显著正相关, 且最大根长(MRL)和根生物量(RFW和RDW)与成株期抗旱性显著正相关, 可以将苗期最大根长、根生物量等作为旱地育种中筛选抗性和产量的早期指示指标。目前应用单株产量进行了研究, 生产条件下群体产量与稀播下个体产量有时存在差异, 对筛选指标的效应会造成偏差, 随着人们对个体-群体及个体竞争对产量形成影响机制的认识, 这种偏差将会更好的解决。
3.3 山西小麦根系变化揭示半干旱地区根系演变趋势
小麦农艺性状演变规律研究历来备受重视。自20世纪40年代以来, 在全国范围内实现了6~9次小麦品种更换, 每次品种更换不仅产量有了很大提高, 而且品种综合性状也会改善。目前不同省份关于株高、叶片、抽穗期、产量和品质等性状的演变规律已较为清楚, 根系是小麦地上部分的根本, 其形态和生理特征的研究较为欠缺, 关于根系演变趋势和特点尚有不同看法。Zhu等[29]选择中国西北地区广泛种植的8个品种, 证明现代品种的根系比早期品种窄, 具有扎根深和横向分布少的特点。张荣等[30]研究黄土高原5 个春麦品种的演变趋势是根系不断减小。田中伟等[31]利用5个品种研究了长江中下游地区根系更替, 发现根系总根长、表面积和根体积显著增大。不同研究结论存在差异, 一是利用的材料份数少导致偶然性过大, 另一方面是采用长江中下游麦区和西南麦区的小麦品种, 这些地区水分充足, 无法客观反映根系对水分胁迫响应。
本文采用可代表半干旱地区的山西小麦为材料,系统研究了苗期根系的演变趋势, 发现苗期最大根长随年代略降低, 而最大根长的生理抗旱性却一直增加; 苗期总根长、根表面积、根体积和根生物量等性状随年代基本表现出先增后降趋势, 但根系性状的抗旱系数则表现为先降后升, 以20世纪80年代和90年代的品种性状值最大。可能是90年代以前品种改良主要以提高根系生物量, 增加水肥吸收, 进而提高产量, 在这个过程中忽略了品种生理抗旱性; 而90年代后产量和地下生物量达到一定平衡, 产量的提高依赖地下根系冗余减少、构型优化、水肥利用效率和抗旱性的提高。根冠比过大会影响地上部生物学产量和经济产量的形成, 造成根系冗余, 因此适宜的根系将会是半干旱区小麦育种的重要指标, 这与Sun等[14]提出的干旱与半干旱地区小麦根系的生态预测一致。可见在今后干旱与半干旱地区的品种选育过程中应人为调控根系的大小, 通过提高水分利用率培育抗旱节水的优良品种, 从而促进小麦抗旱节水的遗传改良。
4 结论
山西小麦苗期根系性状多样性丰富, 总根长、根表面积、根体积、平均根数及根生物量随年代表现出明显的先增后降现象, 各性状抗旱系数整体上呈先降后增的趋势。干旱胁迫抑制了小麦苗期根系生长, 不同根系性状对旱胁迫的响应存在差异, 总根长对水分胁迫最敏感, 其次为根体积、根表面积和根生物量, 最大根长和平均根数敏感性最低。最大根长和根生物量与雨养条件下成株期产量和抗性显著相关, 可作为旱地育种中抗旱高产优异种质的早期筛选指标。
致谢:在材料征集的过程中得到了山西省农业科学院柴永峰、任杰成、张晓军、张东旭、闫金龙、卜斌和任永康等老师, 以及中国农业科学院作物科学研究所景蕊莲研究员的大力协助, 谨致热忱!
[1] Manschadi A M, Christopher J, Devoil P, Hammer G L. The role of root architectural traits in adaptation of wheat to water-limited environments., 2006, 33: 823–837.
[2] Berry P M, Sylvester-Bradly R, Berry S. Ideotype design for lodging-resistant wheat., 2007, 154: 165–179.
[3] Atkinson J A, Wingen L U, Griffiths M, Pound M P, Gaju O, Foulkes M J, Gouis J L, Griffiths S, Bennett M J, King J, Wells D M. Phenotyping pipeline reveals major seedling root growth QTL in hexaploid wheat., 2015, 66: 2283–2292.
[4] Veronica M R, Jorge B V, Luis L B, Rafael J L B. Monitoring wheat root development in a rainfed vertisol: tillage effect., 2010, 33: 182–187.
[5] Ibrahim S E, Schubert A, Pillen K, Léon J. QTL analysis of drought tolerance for seedling root morphological traits in an advanced backcross population of spring wheat., 2012, 2: 619–629.
[6] Placido D F, Campbell M T, Folsom J J, Cui X, Kruger G R, Stephen B P, Walia H. Introgression of novel traits from a wild wheat relative improves drought adaptation in wheat., 2013, 161: 1806–1819.
[7] Feng S W, Gu S B, Zhang H B, Wang D. Root vertical distribution is important to improve water use efficiency and grain yield of wheat., 2017, 214: 131–141.
[8] Zobel R W, Wright S F. Primary and secondary root systems. In: Roots and Soil Management: Interactions between Roots and the Soil. ASA, CSSA, and SSSA, Madison, WI, 2005. pp 3–14.
[9] Cane M A, Maccaferri M, Nazemi G, Salvi S, Francia R, Colalonga C, Roberto T. Association mapping for root architectural traits in durum wheat seedlings as related to agronomic performance., 2014, 34: 1629–1645.
[10] Liu X L, Li R Z, Chang X P, Jing R L. Mapping QTLs for seedling root traits in a doubled haploid wheat population under different water regimes., 2013, 189: 51–66.
[11] Kabir M R, Liu G, Guan P F, Wang F, Khan A A, Ni Z F, Yao Y Y, Hu Z R, Xin M M, Peng H R, Sun Q X. Mapping QTLs associated with root traits using two different populations in wheat (L.)., 2015, 206: 175–190.
[12] Xie Q, Fernando K M C, Mayes S, Sparkes D L. Identifying seedling root architectural traits associated with yield and yield components in wheat., 2017, 119: 1115–1129.
[13] 苗青霞, 方燕, 陈应龙. 小麦根系特征对干旱胁迫的响应. 植物学报, 2019, 54: 652–661. Miao Q X, Fang Y, Chen Y L. Studies in the responses of wheat root traits to drought stress., 2019, 54: 652–661 (in Chinese with English abstract).
[14] Sun Y Y, Zhang S Q, Chen W. Root traits of dryland winter wheat (L.) from the 1940s to the 2010s in Shaanxi province, China., 2020, 10: 5328.
[15] An D G, Su J Y, Liu Q Y, Zhu Y G, Tong Y P, Li J M, Jing R L, Li B, Li Z S. Mapping QTLs for nitrogen uptake in relation to the early growth of wheat (L.)., 2006, 284: 73–84.
[16] Fan X L, Zhang W, Zhang N, Chen M, Zheng S S, Zhao C H, Han J, Liu J J, Zhang X L, Song L Q, Ji J, Liu X G, Ling H Q, Tong Y P, Cui F, Wang T, Li J M. Identification of QTL regions for seedling root traits and their effect on nitrogen use efficiency in wheat (L.)., 2018, 131: 2677–2698.
[17] Ayalew H, Ma X, Yan G. Screening wheat (spp.) genotypes for root length under contrasting water regimes: potential sources of variability for drought resistance breeding., 2015, 201: 189–194.
[18] Christopher J, Christopher M J, Jennings R, Jones S, Fletcher S, Borrell A, Manschadi A M, Jordan D, Mace E, Hammer G. QTL for root angle and number in a population developed from bread wheats () with contrasting adaptation to water- limited environments., 2013, 126: 1563–1574.
[19] Bai C H, Liang Y L, Hawkesford M J. Identification of QTLs associated with seedling root traits and their correlation with plant height in wheat., 2013, 64: 1745–1753.
[20] Hamada A, Nitta M, Nasuda S, Kato K, Fujita M, Matsunaka H, Okumoto Y. Novel QTLs for growth angle of seminal roots in wheat (L.)., 2012, 354: 395–405.
[21] Cao P, Ren Y Z, Zhang K P, Teng W, Zhao X Q, Dong Z Y, Liu X, Qin H J, Li Z S, Wang D W, Tong Y P. Further genetic analysis of a major quantitative trait locus controlling root length and related traits in common wheat., 2014, 33: 975–985.
[22] 肖永贵, 路亚明, 闻伟锷, 陈新民, 夏先春, 王德森, 李思敏, 童依平, 何中虎. 小麦骨干亲本京411及衍生品种苗期根部性状的遗传. 中国农业科学, 2014, 47: 2916–2926. Xiao Y G, Lu Y M, Wen W E, Chen X M, Xia X C, Wang D S, Li S M, Tong Y P, He Z H. Genetic contribution of seedling root traits among elite wheat parent Jing 411 to its derivatives., 2014, 47: 2916–2926 (in Chinese with English abstract).
[23] 景蕊莲, 昌小平, 朱志华, 胡荣海. 小麦幼苗根系形态与反复干旱存活率的关系. 西北植物学报, 2002, 22: 243–249. Jing R L, Chang X P, Zhu Z H, Hu R H. Relationship between root morphology of wheat () and survival percentage under repeated drought condition., 2002, 22: 243–249 (in Chinese with English abstract).
[24] 马富举, 李丹丹, 蔡剑, 姜东, 曹卫星, 戴廷波. 干旱胁迫对小麦幼苗根系生长和叶片光合作用的影响. 应用生态学报, 2012, 23: 724–730. Ma F J, Li D D, Cai J, Jiang D, Cao W X, Dai T B. Responses of wheat seedlings root growth and leaf photosynthesis to drought stress., 2012, 23: 724–730 (in Chinese with English abstract).
[25] Dhanda S S, Sethi G S, Behl R K. Indices of drought tolerance in wheat genotypes at early stages of plant growth., 2004, 190: 6–12.
[26] Landjeva S, Neumann K, Lohwasser U, Börner A. Molecular mapping of genomic regions associated with wheat seedling growth under osmotic stress., 2008, 52: 259–266.
[27] 周晓果, 景蕊莲, 郝转芳, 昌小平, 张正斌. 小麦幼苗根系性状的QTL分析. 中国农业科学, 2005, 38: 1951–1957. Zhou X G, Jing R L, Hao Z F, Chang X P, Zhang Z B. Mapping QTL for seedling root traits in common wheat., 2005, 38: 1951–1957 (in Chinese with English abstract).
[28] 胡雯媚, 王思宇, 樊高琼, 刘运军, 郑文, 王强生, 马宏亮. 西南麦区小麦品种苗期抗旱性鉴定及其指标筛选. 麦类作物学报, 2016, 36: 182–193. Hu W M, Wang S Y, Fan G Q, Liu Y J, Zheng W, Wang Q S, Ma H L. Analysis on the drought resistance and screening of drought resistance appraisal indexes of wheat cultivars in seedling stage in southwest area, 2016, 36: 182–189 (in Chinese with English abstract).
[29] Zhu Y H, Weiner J, Yu M X, Li F M. Evolutionary agroecology: trends in root architecture during wheat breeding., 2019, 12: 733–743.
[30] 张荣, 张大勇. 半干旱区春小麦不同年代品种根系生长冗余的比较实验研究. 植物生态学报, 2000, 24: 298–303. Zhang R, Zhang D Y. A comparative study on root redundancy in spring wheat varieties released in different years in semi-arid area., 2000, 24: 298–303 (in Chinese with English abstract).
[31] 田中伟, 樊永惠, 殷美, 王方瑞, 蔡剑, 姜东, 戴廷波. 长江中下游小麦品种根系改良特征及其与产量的关系. 作物学报, 2015, 41: 613–622. Tian Z W, Fan Y H, Yin M, Wang F R, Cai J, Jiang D, Dai T B. Genetic improvement of root growth and its relationship with grain yield of wheat cultivars in the middle-lower Yangtze river., 2015, 41: 613–622 (in Chinese with English abstract).
[32] 唐淼, 王晓毅, 侯侃, 侯亮亮. 山西晋中小南庄墓地人骨的C、N稳定同位素:试析小麦在山西的推广. 人类学学报, 2018, 37: 318–330. Tang M, Wang X Y, Hou K, Hou L L. Carbon and nitrogen stable isotope of the human bones from the Xiaonanzhuang cemetery, Jinzhong, Shanxi: A preliminary study on the expansion of wheat in ancient Shanxi, China., 2018, 37: 318–330 (in Chinese with English abstract).
[33] 赵佳佳, 乔玲, 郑兴卫, 李晓华, 曹勇, 马小飞, 杨斌, 姬虎太, 乔麟轶, 郑军, 张建诚. 山西小麦育成品种品质性状及HMW-GS组成演变分析. 植物遗传资源学报, 2018, 19: 1126–1137. Zhao J J, Qiao L, Zheng X W, Li X H, Cao Y, Ma X F, Yang B, Ji H T, Qiao L Y, Zheng J, Zhang J C. Variation of quality-related traits and HMW-GS of wheat varieties in Shanxi province., 2018, 19: 1126–1137 (in Chinese with English abstract).
[34] 吕学莲, 白海波, 惠建, 田小燕, 杨宸刚, 马斯霜, 蔡正云, 李树华. 籼粳稻杂交衍生RIL系的苗期抗旱性评价. 植物遗传资源学报, 2019, 20: 556–563 Lyu X L, Bai H B, Hui J, Tian X Y, Yang C G, Ma S S, Cai Z Y, Li SH. Evaluation of seedling drought resistance of RIL derived fromrice andrice., 2019, 20: 556–563 (in Chinese with English abstract).
[35] Ming D F, Pei Z F, Naeem M S, Gong H J, Zhou W J. Silicon alleviates PEG-induced water-deficit stress in upland rice seedlings by enhancing osmotic adjustment., 2012, 198: 14–26.
[36] Blum A. Osmotic adjustment is a prime drought stress adaptive engine in support of plant production., 2017, 40: 4–10.
[37] 慕自新, 张岁岐, 梁爱华, 梁宗锁. 玉米整株根系水导与其表型抗旱性的关系. 作物学报, 2005, 31: 203–208. Mu Z X, Zhang S Q, Liang A H, Liang Z S. Relationship between maize root hydraulic conductivity and drought resistance., 2005, 31: 203–208 (in Chinese with English abstract).
[38] 王贺正, 李艳, 马均, 张荣萍, 李旭毅, 汪仁全. 水稻苗期抗旱性指标的筛选. 作物学报, 2007, 33: 1523–1529. Wang H Z, Li Y, Ma J, Zhang R P, Li X Y, Wang R Q. Screening indexes of drought resistance during seedling stage in rice., 2007, 33: 1523–1529 (in Chinese with English abstract).
[39] 赵言文, 丁艳锋, 陈留根, 黄丕生. 水稻旱育秧苗抗旱生理特性研究. 中国农业科学, 2001, 34: 283–291. Zhao Y W, Ding Y F, Chen L G, Huang P S. Physiological characteristics of drought resistance of rice dry nursery seedlings., 2001, 34: 283–291 (in Chinese with English abstract).
[40] Liao M, Fillery I, Palta J. Early vigorous growth is a major factor influencing nitrogen uptake in wheat., 2004, 31: 121–129.
[41] 陈成升, 谢志霞, 刘小京. 旱盐互作对冬小麦幼苗生长及其抗逆生理特性的影响. 应用生态学报, 2009, 20: 811–816. Chen C S, Xie Z X, Liu X J. Interactive effects of drought and salt stresses on winter wheat seedling growth and physio logical characteristics of stress resistance., 2009, 20: 811–816 (in Chinese with English abstract).
[42] 魏道智, 宁书菊, 林文雄. 小麦根系活力变化与叶片衰老的研究. 应用生态学报, 2004, 15: 1565–1569. Wei D Z, Ning S J, Lin W X. Relationship between wheat root activity and leaf senescence., 2004, 15: 1565–1569 (in Chinese with English abstract).
[43] Sandhu N, Subedi S R, Singh V K, Sinha P, Kumar S, Singh S P, Ghimire S K, Pandey M, Yadaw R B, Varshney R K, Kumar A. Deciphering the genetic basis of root morphology, nutrient uptake, yield, and yield-related traits in rice under dry direct-seeded cultivation systems., 2019, 9: 9334.
[44] 李龙, 毛新国, 王景一, 昌小平, 柳玉平, 景蕊莲. 小麦种质资源抗旱性鉴定评价. 作物学报, 2018, 44: 988–999. Li L, Mao X G, Wang J Y, Chang X P, Liu Y P, Jing R L. Drought tolerance evaluation of wheat germplasm resources., 2018, 44: 988–999 (in Chinese with English abstract).
附表1 山西小麦品种(系)及其审定年限
Table S1 Wheat cultivars bred in Shanxi province and the approval of the year
(续附表1)
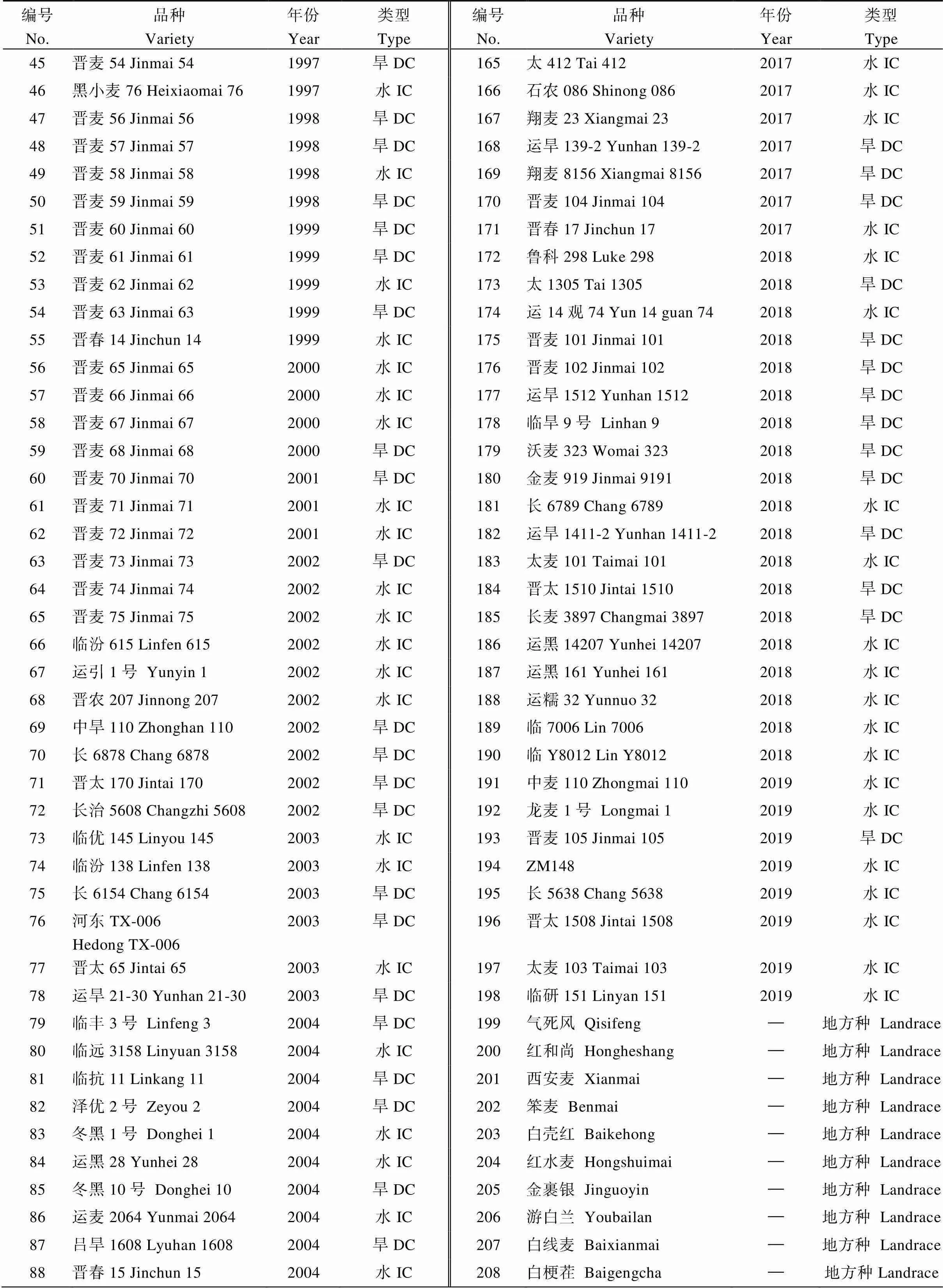
编号No.品种Variety年份Year类型Type编号No.品种Variety年份Year类型Type 45晋麦54 Jinmai 541997旱DC165太412 Tai 4122017水IC 46黑小麦76 Heixiaomai 761997水IC166石农086 Shinong 0862017水IC 47晋麦56 Jinmai 561998旱DC167翔麦23 Xiangmai 232017水IC 48晋麦57 Jinmai 571998旱DC168运旱139-2 Yunhan 139-22017旱DC 49晋麦58 Jinmai 581998水IC169翔麦8156 Xiangmai 81562017旱DC 50晋麦59 Jinmai 591998旱DC170晋麦104 Jinmai 1042017旱DC 51晋麦60 Jinmai 601999旱DC171晋春17 Jinchun 172017水IC 52晋麦61 Jinmai 611999旱DC172鲁科298 Luke 2982018水IC 53晋麦62 Jinmai 621999水IC173太1305 Tai 13052018旱DC 54晋麦63 Jinmai 631999旱DC174运14观74 Yun 14 guan 742018水IC 55晋春14 Jinchun 141999水IC175晋麦101 Jinmai 1012018旱DC 56晋麦65 Jinmai 652000水IC176晋麦102 Jinmai 1022018旱DC 57晋麦66 Jinmai 662000水IC177运旱1512 Yunhan 15122018旱DC 58晋麦67 Jinmai 672000水IC178临旱9号 Linhan 92018旱DC 59晋麦68 Jinmai 682000旱DC179沃麦323 Womai 3232018旱DC 60晋麦70 Jinmai 702001旱DC180金麦919 Jinmai 91912018旱DC 61晋麦71 Jinmai 712001水IC181长6789 Chang 67892018水IC 62晋麦72 Jinmai 722001水IC182运旱1411-2 Yunhan 1411-22018旱DC 63晋麦73 Jinmai 732002旱DC183太麦101 Taimai 1012018水IC 64晋麦74 Jinmai 742002水IC184晋太1510 Jintai 15102018旱DC 65晋麦75 Jinmai 752002水IC185长麦3897 Changmai 38972018旱DC 66临汾615 Linfen 6152002水IC186运黑14207 Yunhei 142072018水IC 67运引1号 Yunyin 12002水IC187运黑161 Yunhei 1612018水IC 68晋农207 Jinnong 2072002水IC188运糯32 Yunnuo 322018水IC 69中旱110 Zhonghan 1102002旱DC189临7006 Lin 70062018水IC 70长6878 Chang 68782002旱DC190临Y8012 Lin Y80122018水IC 71晋太170 Jintai 1702002旱DC191中麦110 Zhongmai 1102019水IC 72长治5608 Changzhi 56082002旱DC192龙麦1号 Longmai 12019水IC 73临优145 Linyou 1452003水IC193晋麦105 Jinmai 1052019旱DC 74临汾138 Linfen 1382003水IC194ZM1482019水IC 75长6154 Chang 61542003旱DC195长5638 Chang 56382019水IC 76河东TX-006 Hedong TX-0062003旱DC196晋太1508 Jintai 15082019水IC 77晋太65 Jintai 652003水IC197太麦103 Taimai 1032019水IC 78运旱21-30 Yunhan 21-302003旱DC198临研151 Linyan 1512019水IC 79临丰3号 Linfeng 32004旱DC199气死风 Qisifeng—地方种 Landrace 80临远3158 Linyuan 31582004水IC200红和尚 Hongheshang—地方种 Landrace 81临抗11 Linkang 112004旱DC201西安麦 Xianmai—地方种 Landrace 82泽优2号 Zeyou 22004旱DC202笨麦 Benmai—地方种 Landrace 83冬黑1号 Donghei 12004水IC203白壳红 Baikehong—地方种 Landrace 84运黑28 Yunhei 282004水IC204红水麦 Hongshuimai—地方种 Landrace 85冬黑10号 Donghei 102004旱DC205金裹银 Jinguoyin—地方种 Landrace 86运麦2064 Yunmai 20642004水IC206游白兰 Youbailan—地方种 Landrace 87吕旱1608 Lyuhan 16082004旱DC207白线麦 Baixianmai—地方种 Landrace 88晋春15 Jinchun 152004水IC208白梗茬 Baigengcha—地方种Landrace
(续附表1)
IC: irrigated cultivar; DC: dryland cultivar.
Seedling root characteristics and drought resistance of wheat in Shanxi province
ZHAO Jia-Jia1,2,**, QIAO Ling2,**, WU Bang-Bang2, GE Chuan1, QIAO Lin-Yi1, ZHANG Shu-Wei1, YANSu-Xian2, ZHENG Xing-Wei2,*, and ZHENG Jun2,*
1Academy of Organic Dry Farming Agricultural Research, Shanxi Agriculture University, the Key Laboratory of Organic Dry Farming of Shanxi Province, Taiyuan 030006, Shanxi, China;2Institute of Wheat Research, Shanxi Agriculture University, Linfen 041000, Shanxi, China
Wheat root morphology at the seedling stage is the basis of root distribution at the adult stage. It is closely related to stress resistance and yield. A comprehensive understanding of the root system characteristics at seedling stage and drought resistance is of great significance for the excellent germplasm utilization and early screening of drought resistance. Using 239 wheat varieties (lines) from Shanxi province, the root traits at seedling stage and their response to water stress were evaluated. The results showed that under normal growth, Shanxi wheat had a great variation in root traits at seedling stage, with the greatest variation in landraces. The maximum root length (MRL) tended to decrease slightly with the years, while other root traits were first increased and then decreased. There were differences among the root traits in response to water stress. The total root length (TRL) is the most sensitive to water, followed by root surface area (RSA), root volume (RV) and root biomass. The maximum root length (MRL) and the average number of roots (RN) were insensitive. The drought resistance of seedling roots showed a trend of decreasing first and then increasing with the years. Landraces and varieties released from 1970 to 1979 had moderately resistance to water stress, varieties released from 1980 to 1999 had lower drought resistance, and varieties released after 2000 had the better resistance, of which the dryland varieties were the best resistance. Correlation analysis of the seedling root characteristics and yield related traits suggested that the maximum root length, total root length, root volume and root biomass were significantly positively correlated with 1000-kernel weight and yield under rain-fed conditions, and the maximum root length and root biomass were also significantly positively correlated with adult plant drought resistance. The present study suggests that the maximum root length and root biomass at the seedling stage can be used as early generation selection parameters for drought resistance and yield in dryland breeding in semi-arid areas.
Shanxi wheat; seedling root system; drought resistance characteristics; evolution trend.
10.3724/SP.J.1006.2021.01048
本研究由有机旱作山西省重点实验室开放基金项目(201805D111015-2), 山西省自然科学基金项目(201901D211567), 山西省农业科学院科技创新项目(YCX2018413)和山西省重点研发计划项目(201803D421021)资助。
This study was supported by the Key Laboratory of Organic Dry Farming of Shanxi Province (201805D111015-2), the Natural Science Foundation of Shanxi Province (201901D211567), the Agricultural Science and Technology Project (YCX2018413), and the Shanxi Province Key Research and Development Program (201803D421021).
郑军, E-mail: sxnkyzj@126.com; 郑兴卫, E-mail: smilezxw@126.com
**同等贡献(Contributed equally to this work)
赵佳佳, E-mail: jjzh1990@163.com
2020-06-08;
2020-09-13;
2020-09-25.
URL: https://kns.cnki.net/kcms/detail/11.1809.S.20200925.1646.004.html
