减压贮藏对去壳黄甜竹笋的保鲜效果及其生理和分子机制
2021-01-28周成敏庞林江翁方荣
戴 丹 郑 剑 周成敏 庞林江 翁方荣
(1 浙江农林大学信息工程学院/林业感知技术与智能装备国家林业和草原局重点实验室/浙江省林业智能监测与信息技术研究重点实验室,浙江 杭州 311300;2 浙江农林大学农业与食品科学学院/浙江省农产品品质改良技术研究重点实验室,浙江 杭州 311300;3 浙江省丽水市农林科学研究院,浙江 丽水 323000;4浙江省丽水市遂昌县羽峰食品厂,浙江 遂昌 323300)
黄甜竹(Acidosasa edulis) 是禾本科酸竹属(Acidosasa)混生竹,广泛种植于浙西南地区。黄甜竹笋口感鲜甜、细腻,富含氨基酸、糖类、膳食纤维等营养素;因其于气温较高的4 ~5月份出笋,且其采后极易发生褐变、木质化和腐烂变质,因此保鲜难度大,严重制约其运输销售,影响产业发展。
减压贮藏能够通过降低贮藏环境的气压进而降低氧气含量,从而抑制果蔬呼吸,且减压贮藏可以快速去除呼吸热[1]。相关研究表明,减压贮藏能够显著抑制苹果[1]、芦笋[2]、甜樱桃[3]、草莓[4-5]、青辣椒[6]、绿皮西葫芦[7]、番茄[8-9]、水蜜桃[10]、鲜切西兰花[11]、双孢菇[12]、杏鲍菇[13]、松茸[14]和芒果[15]等果蔬和食用菌采后成熟、衰老以及品质劣变的速度,其作用机制主要是通过延缓果实硬度下降和叶绿素降解、抑制呼吸速率和失重率上升、减轻膜脂过氧化程度和延缓褐变发生、抑制维生素C 等营养成分损失等途径,有效延缓食用品质下降并延长贮藏期。郑先章等[16]提出,减压贮藏短期处理对生鲜果蔬具有后续保鲜效应,可在一定程度上弥补目前生鲜果蔬等冷链流通过程中断链的缺陷,为生鲜果蔬销售流通提供一定的技术支撑。
去壳净笋产业近些年来快速发展,去壳净笋以其加工烹饪方便以及无笋壳等固体废弃物污染等优点广受城市消费者青睐。然而,关于减压贮藏对去壳冷藏黄甜竹笋的褐变、笋肉木质化以及品质影响的研究尚鲜见报道。鉴于此,本试验以去壳黄甜竹笋为原料,研究减压贮藏对延缓黄甜竹笋冷藏期间木质化、褐变进程的影响效果及其内在调控机制,以期为黄甜竹净笋流通销售过程中保鲜方法的筛选以及保鲜包装盒的设计研发提供数据参考。
1 材料与方法
1.1 材料与试剂
黄甜竹笋,5月初上午6:00 左右采收于浙江省丽水市农林科学研究院的黄甜竹林。
L-苯丙氨酸、愈创木酚、三氯乙酸(trichloroacetic acid,TCA)、硫代巴比妥酸(thiobarbituric acid,TBA)、聚乙烯吡咯烷酮(polyvinyl pyrrolidone,PVP)、邻苯二酚、乙二胺四乙酸(ethylene diamine tetraacetic acid,EDTA)、TritonX-100、β-巯基乙醇,均为分析纯,国药集团化学试剂有限公司;松柏醇、烟酰胺腺嘌呤二核苷酸磷酸(氧化态),美国Sigma 公司;PureLink©plant RNA Extraction Kit、PrimeScriptTMⅡ1st Strand cDNA Synthesis Kit、SYBR©Premix Ex TaqTM(Tli Rnase Plus),日本TaKaRa Bio InC 公司。
1.2 仪器与设备
VSE-5 型减压贮藏小型实验设备,宁波象山食品设备有限公司;DDS-307 台式电导率仪,上海仪电科学仪器股份有限公司;3-30K 冷冻离心机,德国Sigma公司;TYI-3016F 便携式红外CO2分析仪,上海唐仪电子科技有限公司;UV-2355 紫外可见分光光度计,尤尼柯(上海)仪器有限公司;LHS-350SC 恒温恒湿箱,上海科辰实验设备有限公司;TA-XT2i 质构仪,英国Stable Micro Systems 公司;CHROMA METER CR-400 色差仪,日本柯尼卡公司;iQTM5 多重实时荧光定量PCR 仪,美国Bio-Rad 公司。
1.3 试验方法
1.3.1 预处理 现场采挖黄甜竹笋,拣选基部切口平整、笋体其他部位无机械损伤、无病虫害且粗细和长短相近的笋,置于采样盒中,采用空调车3 h 内运回实验室。切除自基部切口向上长度约3 ~4 cm 部分,剥去笋壳,自来水冲洗干净后于阴凉通风处晾干,随机取180 根笋,分成6 组,其中1 组用于测定贮藏初期(0 d)的各项指标;另外4 组置于减压贮藏设备的贮藏室(减压贮藏实验设备结构原理如图1 所示),进行预试验,分别于85±5、70±5、55±5 和40±5 kPa 减压环境下,6±1℃、相对湿度80%~85%条件下贮藏10 d;最后1 组在101 kPa 空气环境下同温湿度冷藏作为预试验对照(CK1)。通过测定硬度、L*值、木质子和纤维素含量筛选出最适处理参数。

图1 减压贮藏实验设备结构原理图Fig.1 Structure schematic diagram of the hypobaric storage test equipment
1.3.2 试验方法 将剩余360 根笋,分成2 组,一组贮藏于1.3.1 筛选出的最适气压条件,另一组贮藏于空气中(对照,CK),于6±1℃、80%~85%(相对湿度)条件下贮藏10 d。贮藏期间,每2 d 随机取样,每组每次取30 根笋,检测基部切面的色差、测定呼吸速率,选取自基部切口往上2 ~4 cm 的一段,检测该段笋肉组织的相对电导率、丙二醛(malondialdehyde,MDA)含量、硬度、木质素和纤维素含量以及苯丙氨酸解氨酶(phenylalanine ammonia lyase,PAL)、过氧化物酶(peroxidase,POD)、肉桂醇脱氢酶(cinnamyl alcohol dehydrogenase,CAD)、 多 酚 氧 化 酶( polyphenol oxidase,PPO)的活性及其编码基因的相对表达量,试验重复3 次。
1.3.3 呼吸速率和失重率的测定 取6 个密封良好且洁净的干燥器(其中3 个连接真空泵维持减压状态),分别在相应的贮藏温度(6 ±1℃)下,先用便携式红外线CO2分析器检测各自干燥器中CO2浓度(C0),然后将CK 和减压贮藏的去壳黄甜竹笋放入各自对应的干燥器中,密封后两组笋在各自对应的常压和减压环境下放置2 h,再检测干燥器中CO2浓度(C1),根据前后两次CO2浓度变化、干燥器的容积(V干)与笋的体积(V笋)以及笋的质量(m笋)计算呼吸速率。

失重率采用称重法测定。失重率=(贮藏前质量-贮藏后质量)/贮藏前质量×100%
1.3.4 色差的测定 采用色差仪测定黄甜竹笋基部切面的L*值,3 次重复,取平均值。
1.3.5 相对电导率和MDA 含量的测定 相对电导率的测定参照曹建康等[17]的方法稍作修改:测定样品为10 片直径1 cm、厚度1 mm 左右的笋肉组织圆片;MDA 含量的测定参照曹建康等[17]的方法。
1.3.6 硬度的测定 从黄甜竹笋基部切口往上2 ~4 cm 部位切下约1.0 ~1.4 cm 厚笋肉,切成约1 cm×1 cm 的正方形,然后用质构仪P/2 平头柱形探头检测笋肉的硬度,探头测试深度为4 mm,穿刺速度为0.5 mm·s-1,单位为N。
1.3.7 纤维素和木质素的测定 参照Zeng 等[18]的方法,结果以占笋肉鲜重质量百分比计。
1.3.8 PAL、POD、CAD、PPO 活性测定 PAL、POD、PPO 活性的测定参照曹建康等[17]的方法,CAD 活性的测定参照Zeng 等[18]的方法。
1.3.9 黄甜竹笋木质素代谢和褐变相关酶基因表达的测定 参照文献[19-20]的方法并稍作修改,用PrimeScriptTMⅡ1st Strand cDNA Synthesis Kit 进行逆转录试验。
Real-time PCR 检测:用多重实时荧光定量PCR仪进行扩增,引物及条件见表1,反应体系共16 μL,包括:6.6 μL ddH2O、8 μL SYBR Green qPCR mix(2×)、0.2 μL PCR-F(10 μmol·L-1)、0.2 μL PCR-R(10 μmol·L-1)、 1.0 μL 模板cDNA。
反应条件为:95℃预变性1 min;50 个循环(95℃变性10 s,60℃退火25 s,收集荧光);在55~95℃之间进行熔解曲线分析。
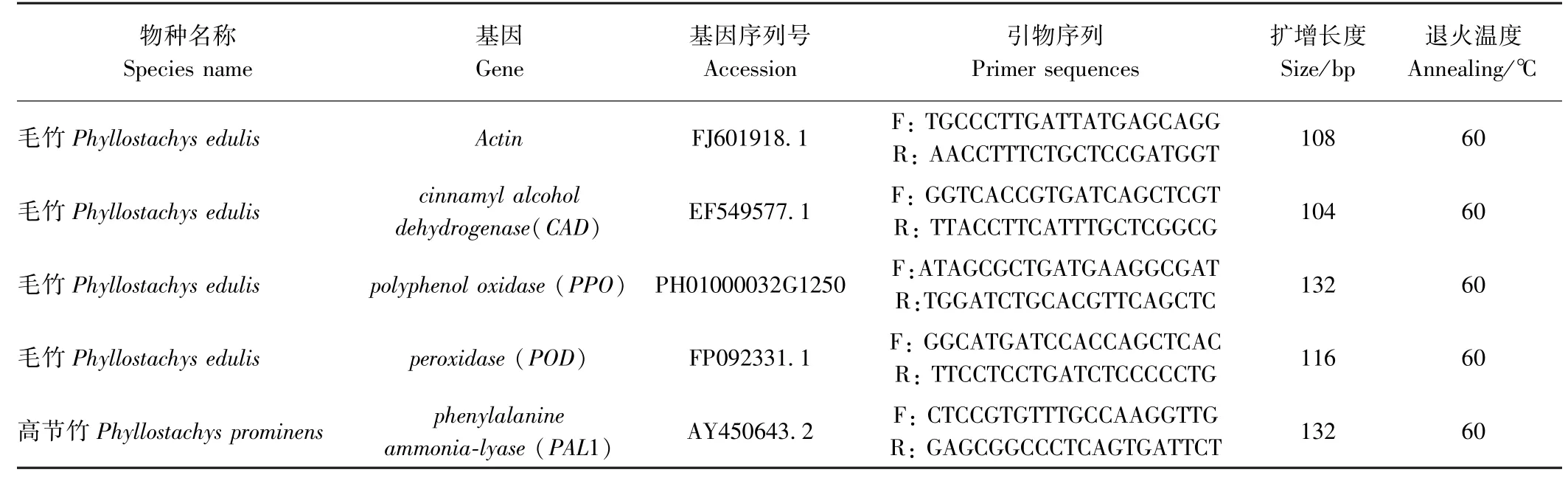
表1 定量PCR 引物序列及反应条件Table 1 Real-time PCR primers and conditions
1.4 数据分析
采用SPSS 16.0 软件中的one-way ANOVA 方法对数据进行显著性检验分析,用Duncan 进行多重比较分析(P<0.05),用Excel 2010 软件作图。
2 结果与分析
2.1 黄甜竹笋减压贮藏参数的预试验筛选
由表2 可知,与0 d 的去壳黄甜竹笋相比,常压处理(101 kPa)和各减压处理组竹笋的硬度和笋肉木质素含量均明显上升。与常压处理组相比,4 组减压处理均显著抑制了竹笋硬度和木质素含量的上升,然而85 和70 kPa 减压贮藏去壳黄甜竹笋的失重率显著升高;另外70、55 和40 kPa 减压贮藏显著抑制了笋肉组织中纤维素含量的上升和L*值的下降,而85 kPa 减压贮藏虽然抑制了笋肉组织中纤维素含量含量上升和L*值下降,但未达到显著水平;此外,55 kPa 减压贮藏在抑制笋肉硬度和失重率上升方面略好于40 kPa 减压贮藏,但差异不显著,同时在木质素和纤维素含量以及L*值方面差异也不显著。综合考虑以上结果以及减压设备运营等因素,确定后续以55 kPa 减压贮藏开展试验。
2.2 减压贮藏对黄甜竹笋呼吸速率和L*值的影响
由图2-A 可知,黄甜竹笋呼吸速率随着贮藏时间的延长逐渐升高,贮藏第2 天时,减压贮藏组黄甜竹笋的呼吸速率略低于CK 但差异不显著,贮藏4 ~10 d内,减压贮藏组的呼吸速率均显著低于CK。
由图2-B 可知,随着贮藏时间的延长,黄甜竹笋切面的L*值逐渐降低,在贮藏第2 天,减压贮藏组黄甜竹笋的L*值略高于CK 但差异不显著,而贮藏4~10 d内,减压冷藏组黄甜竹笋基部切面的L*值均显著高于CK。
由图3 可知,贮藏第10 天时,CK 组黄甜竹笋的切面和笋体发生较明显褐变,笋肉和外表面呈明显的棕褐色且有明显酸腐味;而减压贮藏组黄甜竹笋的笋体颜色大部分仍呈淡黄色,切面颜色为淡棕黄色、轻微褐变、笋体比较饱满且仍然有黄甜竹笋特有的鲜甜味。表明减压贮藏能够有效抑制去壳黄甜竹笋贮藏期间呼吸速率的升高并延缓褐变。

表2 不同减压贮藏参数对黄甜竹笋硬度、L*值、木质素和纤维素含量以及失重率的影响Table 2 Effects of different hypobaric conditions on firmness, L*,lignin and cellulose contents and weight loss of bamboo shoot
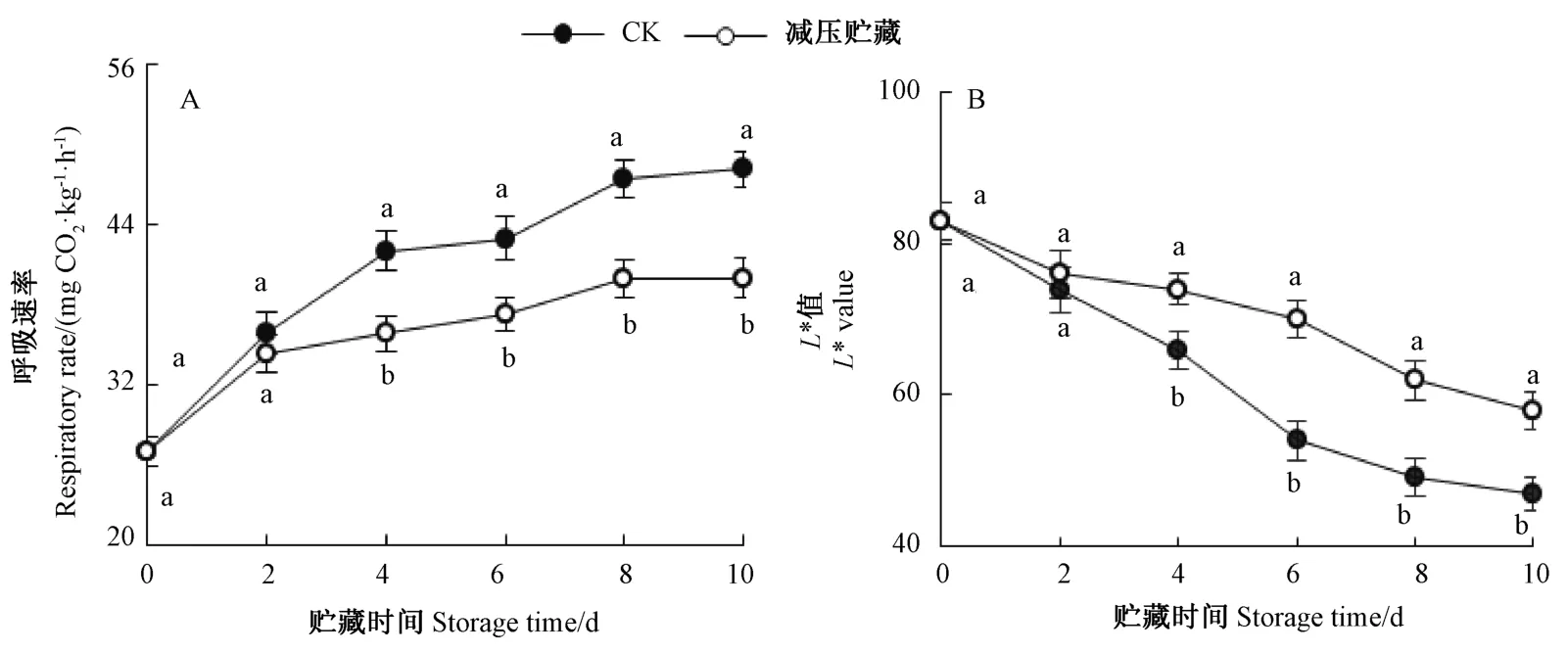
图2 减压贮藏对黄甜竹笋呼吸速率和L*值的影响Fig.2 Effects of hypobaric storage on respiratory rate and L* value of bamboo shoots
2.3 减压贮藏对黄甜竹笋MDA 含量和相对电导率的影响
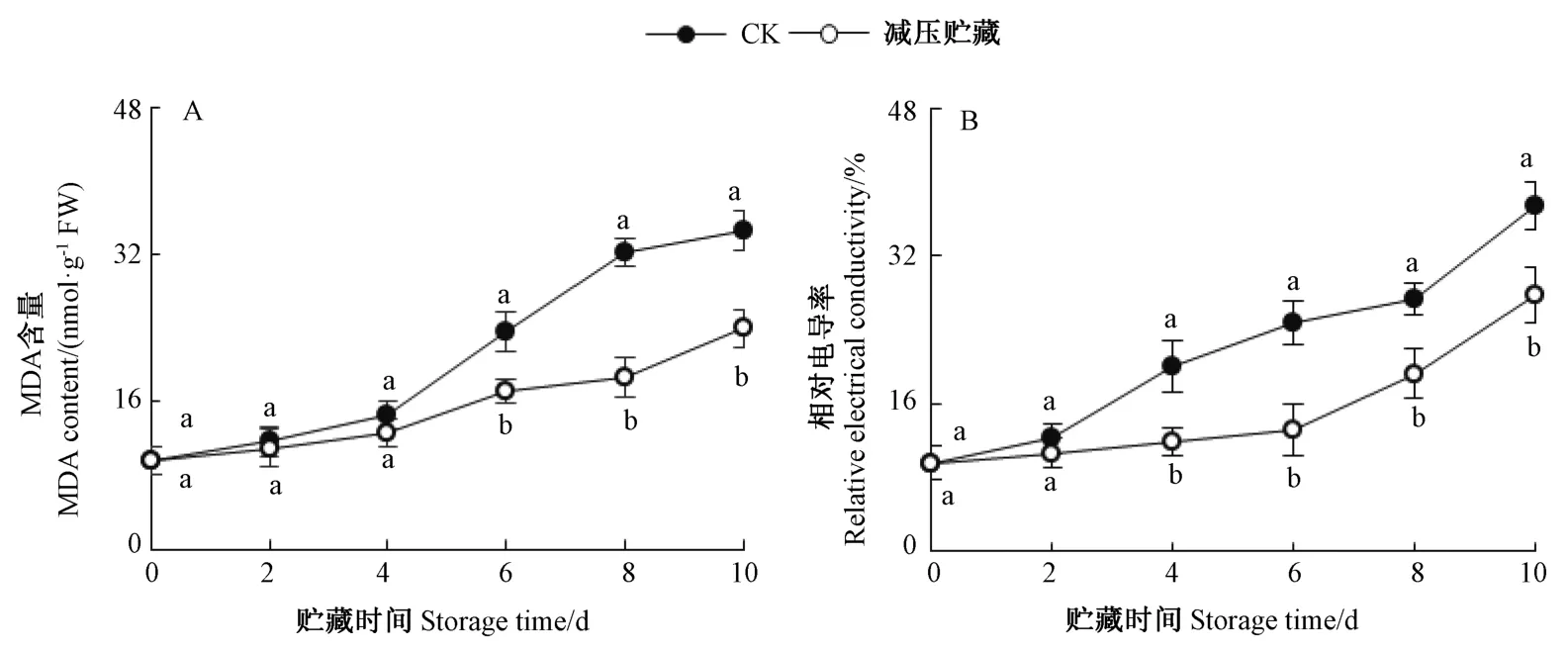
图4 减压贮藏对黄甜竹笋MDA 含量和相对电导率的影响Fig.4 Effects of hypobaric storage on MDA content and relative electrical conductivity of bamboo shoots
由图4-A 可知,黄甜竹笋MDA 含量随着减压贮藏时间的延长呈上升趋势,在贮藏0 ~4 d 内,减压贮藏组黄甜竹笋的MDA 含量略低于CK 但差异不明显,而贮藏6~10 d 时其含量显著低于CK。
由图4-B 可知,黄甜竹笋的相对电导率随着减压贮藏时间的延长逐渐升高。贮藏第2 天时,减压贮藏组黄甜竹笋的相对电导率略低于CK 但差异不显著,贮藏4~10 d 内,显著低于CK。表明减压贮藏能够有效抑制去壳黄甜竹笋组织MDA 的累积和相对电导率的升高,有助于保持细胞膜的完整性。
2.4 减压贮藏对黄甜竹笋硬度、木质素和纤维素含量的影响
由图5-A、B 可知,黄甜竹笋笋肉组织的硬度和木质素含量均随着减压贮藏时间的延长呈逐渐上升趋势,在贮藏0~4 d 内,减压贮藏抑制笋肉硬度和木质素含量上升的效果不显著,而贮藏6 ~10 d,减压贮藏显著抑制了笋肉组织硬度和木质素含量的上升。同时,贮藏4~10 d 时,减压贮藏在也显著抑制了笋肉组织中纤维素的累积(图5-C)。表明减压贮藏能够有效抑制去壳黄甜竹笋组织中木质素和纤维素的累积,并延缓笋肉硬度的上升。

图5 减压贮藏对黄甜竹笋硬度、木质素和纤维素含量的影响Fig.5 Effects of hypobaric storage on firmness and contents of lignin and cellulose of bamboo shoots
2.5 减压贮藏对PAL、POD、CAD、PPO 活性的影响
由图6 可知,CK 与减压贮藏黄甜竹笋的PAL 和CAD 活性在贮藏期内均呈上升趋势,减压贮藏组黄甜竹笋的PAL 和CAD 活性在贮藏4 ~10 d 内显著低于CK。CK 与减压贮藏黄甜竹笋的POD 和PPO 活性变化规律相似,在贮藏2 ~10 d 内减压贮藏显著抑制了笋肉组织POD 和PPO 活性。表明减压贮藏能够有效抑制去壳黄甜竹笋中木质素合成关键酶和褐变关键酶活性的上升。

图6 减压贮藏对黄甜竹笋的PAL、POD、CAD、PPO 活性的影响Fig.6 Effects of hypobaric storage on activities of PAL,POD,CAD and PPO of bamboo shoots
2.6 减压贮藏对黄甜竹笋PAL1、POD、CAD 和PPO基因表达的影响
CK 与减压贮藏组黄甜竹笋中PAL1 和CAD基因相对表达水平在贮藏期内总体呈下降趋势,且伴有一定的波动,减压贮藏组黄甜竹笋的PAL1 基因相对表达水平在贮藏第2 天和贮藏6 ~8 d 时显著低于CK,CAD基因相对表达水平在贮藏2、6、10 d 显著低于CK。CK 与减压贮藏组黄甜竹笋的POD基因相对表达水平在贮藏期内呈先上升而后下降再上升的波动变化,减压贮藏组黄甜竹笋的POD基因相对表达水平在贮藏6~10 d 时显著低于CK。CK 与减压贮藏组黄甜竹笋的PPO基因相对表达水平在贮藏期内也呈先上升而后下降再较平缓波动的变化趋势,减压贮藏组黄甜竹笋PPO基因相对表达水平在贮藏第2 天和贮藏6~10 d 时显著低于CK。表明减压贮藏能够在大部分贮藏期间有效下调去壳黄甜竹笋中木质素合成关键酶和褐变关键酶的基因表达水平。
3 讨论
呼吸会导致果蔬水分和干物质的损失,引起采后果蔬失重失鲜等[18-19]。因此,降低呼吸速率对于延缓采后黄甜竹笋的品质下降至关重要。本研究发现,55 kPa 减压贮藏条件下能够显著抑制去壳黄甜竹笋冷藏过程中呼吸速率的上升,与An 等[4]和程曦等[12]减压贮藏草莓和皱叶莴苣以及双孢菇的研究结果一致。说明减压贮藏能够通过抑制黄甜竹笋的呼吸速率来保持品质。

图7 减压贮藏对黄甜竹笋PAL1、CAD、POD、PPO 基因表达的影响Fig.7 Effects of hypobaric storage on relative gene expression levels of PAL1、CAD、POD and PPO in bamboo shoots
有学者指出,采收期间的机械损伤会诱导竹笋笋肉组织中木质素合成代谢加速,促进纤维素、木质素持续快速合成并导致笋肉硬度快速增加,使竹笋采后食用品质迅速下降[18-19,21]。木质素是植物苯丙烷类代谢中一系列酶促反应的产物,其中PAL 是该代谢途径第一个关键限速酶,CAD 是中间步骤的关键酶,而POD 是最后一个关键酶,能催化木质素单体聚合生成木质素大分子[18,21]。Zheng 等[21]研究发现,外源草酸处理能够通过抑制去壳高节笋中PAL、CAD 和POD等酶的活性进而抑制笋肉组织硬度的上升以及木质素和纤维素的累积,与Luo 等[22]研究热处理抑制竹笋组织中木质素累积的结果和所阐述的调控机制一致。本研究结果表明,与CK 相比,55 kPa 减压贮藏显著抑制了去壳黄甜竹笋笋肉PAL、CAD 和POD 活性的上升,同时抑制了笋肉硬度的增加以及纤维素和木质素的累积,与Gao 等[23]研究减压贮藏延缓低温冷藏下枇杷果肉木质化的结果以及所揭示的机制一致。前人研究发现,果蔬中木质素代谢关键酶的基因表达也参与调控木质素的累积,如EjCAD1、EjPOD的表达与枇杷的木质化进程密切相关[24],氯化钙处理能够通过下调梨果肉中PpCAD1 和PpCAD2 的基因表达抑制木质素的合成[25]。本研究结果表明,与CK 相比,减压贮藏中后期显著下调了PAL1、POD和CAD基因的相对表达量。说明,减压贮藏能够抑制木质素合成关键酶的活性及其基因相对表达量进而抑制去壳黄甜竹笋笋肉组织的木质化进程,从而保持其冷藏期间较好的品质。
诸多学者研究发现,果蔬和鲜切果蔬贮藏期间的褐变主要是由于细胞膜受损后,有氧条件下酚类底物在PPO 和POD 催化下发生的酶促氧化反应所致,褐变严重降低了果蔬的品质和商品价值[26-27]。研究发现,采后一些预处理措施辅助冷藏能够有效抑制竹笋的褐变,如,采后水杨酸和外源草酸处理均能有效抑制细胞膜的损伤并延缓PPO 和POD 活性上升,进而有效抑制了竹笋冷藏期间的褐变[21,28]。本研究发现,减压贮藏中后期显著抑制了去壳黄甜竹笋中MDA 的累积和相对电导率的上升,同时迟滞了PPO 和POD 活性上升,并有效延缓了切面L*值下降,抑制了切面褐变,与Song 等[29]在减压贮藏水蜜桃中的研究结果相似。果蔬中褐变关键酶的基因表达也参与调控其褐变进程,如,UV-C 处理能够通过下调双胞蘑菇的蘑菇柄和菌盖中AbPPO(AbPPO2,AbPPO3 和AbPPO4)基因的相对表达量抑制蘑菇柄和菌盖的褐变[30]。本研究中,与CK 相比,减压贮藏在大部分贮藏时间内显著下调了POD和PPO基因的相对表达量。综上可知,减压贮藏能够有效抑制去壳黄甜竹笋冷藏期间褐变进程,延缓其品质下降。
4 结论
本研究结果表明,55 kPa 减压贮藏显著抑制了去壳黄甜竹笋冷藏期间的呼吸速率,有效维持了细胞膜的完整性并通过抑制POD 和PPO 活性以及下调编码基因的表达量显著抑制了黄甜竹笋基部切面的褐变,通过抑制PAL、POD 和CAD 的活性及下调其编码基因的相对表达量显著抑制了笋肉组织中纤维素和木质素含量以及硬度的上升,进而延缓了产品感官和食用品质的下降。表明减压贮藏可以作为去壳黄甜竹净笋采后保鲜的一种新方法。
