利用红外相机研究新龙县食肉目兽类及其猎物的时间生态位特征与重叠
2021-01-26陈超侯蓉吴蔚张志和古晓东齐敦武
陈超,侯蓉,吴蔚,张志和,古晓东,齐敦武*
(1.成都大熊猫繁育研究基地,四川省濒危野生动物保护生物学省部共建实验室,成都610081;2.四川省大熊猫科学研究院,成都610081;3.四川林业和草原局科研教育处,成都610081)
生态位是现代生态学发展起来的一种重要理论,以数学的方法测度物种在有限资源的多维空间中综合利用资源的能力、利用资源多样化的程度和竞争水平(黄英姿,1994;余世孝,Orlioci,1994;郭水良,李扬汉,1998;周丹等,1999;张林静等,2002;李等,2003),是评价种间和种内关系及其种群在群落中地位的重要手段(张继义等,2003;陈文业等,2010),已成为生态学中最活跃的研究领域之一。生物多样性的丧失将严重损害生态系统的生产能力,致使其无法维持高的质量且难以从小的变化中恢复(Wormetal.,2006)。同时,关键物种的丧失会严重损害生态系统的稳定性,破坏生态系统的功能(Hooperetal.,2012)。这种影响首先表现在物种的种群水平上,即种群的消失速度远快于物种的灭绝速度(Dirzo &Raven,2003)。世代较长的物种在栖息地破碎化和丧失过程中具有较强的灭绝延迟能力(Kuussaarietal.,2009)。物种对生态系统的影响程度最终是由其物种生态位决定(Cardinaleetal.,2006)。
物种生态位核心是其生态需求和在一个给定的群落中与其他物种的相互关系(如捕食关系;孙儒泳,2001)。捕食关系影响着捕食者与被捕食者的生境利用、种群密度、种群结构等(李博等,2000)。合适的生境能满足野生动物繁殖、取食、休息、隐蔽和扩散的需要,动物对不同生境的利用直接关系到物种和生境的保护和管理(魏辅文等,1999)。生境利用主要体现动物对资源的依赖性和选择性,可以体现在空间上和时间上的选择(尚玉昌,2001)。不同动物对资源的利用存在较大程度的趋同性,其生态位随着不同季节的变化出现明显的改变(付和平等,2005)。
青藏高原东缘山区是我国生物多样性较为丰富的区域之一,也是生态系统保存较完好的区域,四川省甘孜藏族自治州就是这一区域良好代表。因此,选择四川省甘孜藏族自治州新龙县,利用红外相机记录区内野生动物活动的时间,探讨新龙县食肉目Carnivora及其潜在猎物在时间生态位上的分化与重叠,以期进一步了解该区域内各物种的种间关系与共存机制,为后期的科学研究和野生动物保护工作提供参考与依据。
1 研究地区
新龙县位于四川省甘孜藏族自治州中部,是四川省三大林区县之一、长江上游雅砻江水系的重点林业县、拥有我国西南青藏高原最大的原始森林之一,森林覆盖率为26%。研究区域位于新龙县西南部,100.140 80°~100.387 70°E,30.504 04°~30.871 42°N,海拔3 136~4 265 m,总面积729.41 km2,属高原季风气候,表现出春秋连季、冬长无夏、雨热同季的特点(邓林,杨海真,2010;柴成秋,2013)。
2 研究方法
2.1 野外数据收集
2013年1月8日—2014年12月30日,结合当地前期监测和大型食肉动物生活习性及红外触发相机在野外的实际应用(王玉玢等,2013;王亮等,2014),选择100个监测点安装红外相机(Lti6210)(图1),设置为全年不间断工作,间隔 1 min 拍照1次。
根据新龙县气候特征,将1年划分为生长季和非生长季(生长季:5月1日—9月30日,非生长季:10月1日—4月30日)。
2.2 相对多度分析
将一天平均分为8个时间段:00∶01—03∶00、03∶01—06∶00、06∶01—09∶00、09∶01—12∶00、12∶01—15∶00、15∶01—18∶00、18∶01—21∶00和21∶01—24∶00。记录每个物种的出现时间点,若同一地点重复拍摄同一动物,则间隔30 min记录第二次,30 min内为1张独立有效照片。
使用独立有效照片和相机工作日,统计食肉目和非食肉目在不同季节的相对多度指数(relative abundance index,RAI)(李晟等,2016;陈立军等,2019):
RAI=Ni/T×1 000。
式中,T为各季节所有相机的工作日总和,Ni为i物种在所有相机位点拍摄到的独立有效照片数。
2.3 时间生态位宽度
时间生态位宽度(Bi)以Shannon-Wiener多样性指数为基础(孙儒泳,2001;李德志等,2006;杨春文等,2008):
式中,j为时间段,Nij为i物种在j时段出现的独立有效照片数,r为生态位的时间段总数,即24 h被划分的时间段数(r=8)。0≤B≤1,其中,0表示该物种在所有时间段未出现,1表示该物种在所有时间段都出现。
2.4 时间生态位重叠指数
时间生态位重叠指数(Cih)采用Schoener(1968)的公式计算(孙儒泳,2001;杨春文等,2008):
Cih=1-1/2Σ︱Nij/Ni-Nhj/Nh︱。
式中,Ni为i物种在所有时间段出现的次数,Nhj为h物种在j时间段的出现次数,Nh为h物种在所有时间段出现的次数。0≤C≤1(0表示2个物种的时间生态位完全没有重叠,1表示2个物种的时间生态位完全重叠)。
3 结果
3.1 物种相对多度
共记录到哺乳动物17种,另有未能鉴定出具体物种的鼠类,统归为鼠科Muridae。选择食肉目的9个物种(狼Canislupus、赤狐Vulpesvulpes、豹猫Prionailurusbengalensis、金钱豹Pantherapardus、黑熊Ursusthibetanus、黄喉貂Martesflavigula、黄鼬Mustelasibirica、猪獾Arctonyxcollaris和香鼬Mustelaaltaica)作为主要研究对象,选择非食肉目的9个物种作为潜在猎物(灰尾兔Lepusoiostolus、中华鬣羚Capricornismilneedwardsii、林麝Moschusberezovskii、毛冠鹿Elaphoduscephalophus、猕猴Macacamulatta、马来水鹿Cervusequinus、野猪Susscrofa、隐纹花松鼠Tamiopsswinhoei和鼠科Muridae)。
生长季相机总工作日为8 876 d,非生长季相机总工作日为18 227 d,共收集到独立有效照片4 148 张。其中,生长季食肉目230张,非食肉目1 157张;非生长季食肉目469张,非食肉目2 292张。18种兽类生长季相对多度为156.26,其中,食肉目的猪獾和黄鼬相对多度较高(分别为8.22和5.75),非食肉目的毛冠鹿相对多度最高(38.19),其次是灰尾兔(26.14);非生长季的相对多度为151.48,其中,食肉目的赤狐和狼相对多度较高(分别为6.80和5.16),非食肉目的毛冠鹿相对多度最高(31.11),其次是马来水鹿(28.91)(表1)。

表1 不同季节18个物种的相对多度指数Table 1 Relative abundance index of 18 species in different seasons
3.2 时间生态位宽度
食肉目中,狼和金钱豹在非生长季的时间生态位宽度较宽,分别为0.990和0.929;黑熊在生长季的时间生态位宽度为0.832,而非生长季的仅为0.306;豹猫、金钱豹和黄鼬在非生长季的生态位宽度大于生长季,赤狐、黄喉貂、猪獾和香鼬在2个季节的时间生态位宽度相近。非食肉目中,马来水鹿、毛冠鹿和中华鬣羚在非生长季的时间生态位宽度较宽,分别为0.975、0.970和0.957,并且都大于生长季(分别为0.871、0.913和0.916);林麝和灰尾兔的时间生态位宽度在2个季节的变化不大;豹猫在生长季的时间生态位最低(0.500),并且小于非生长季(表2)。
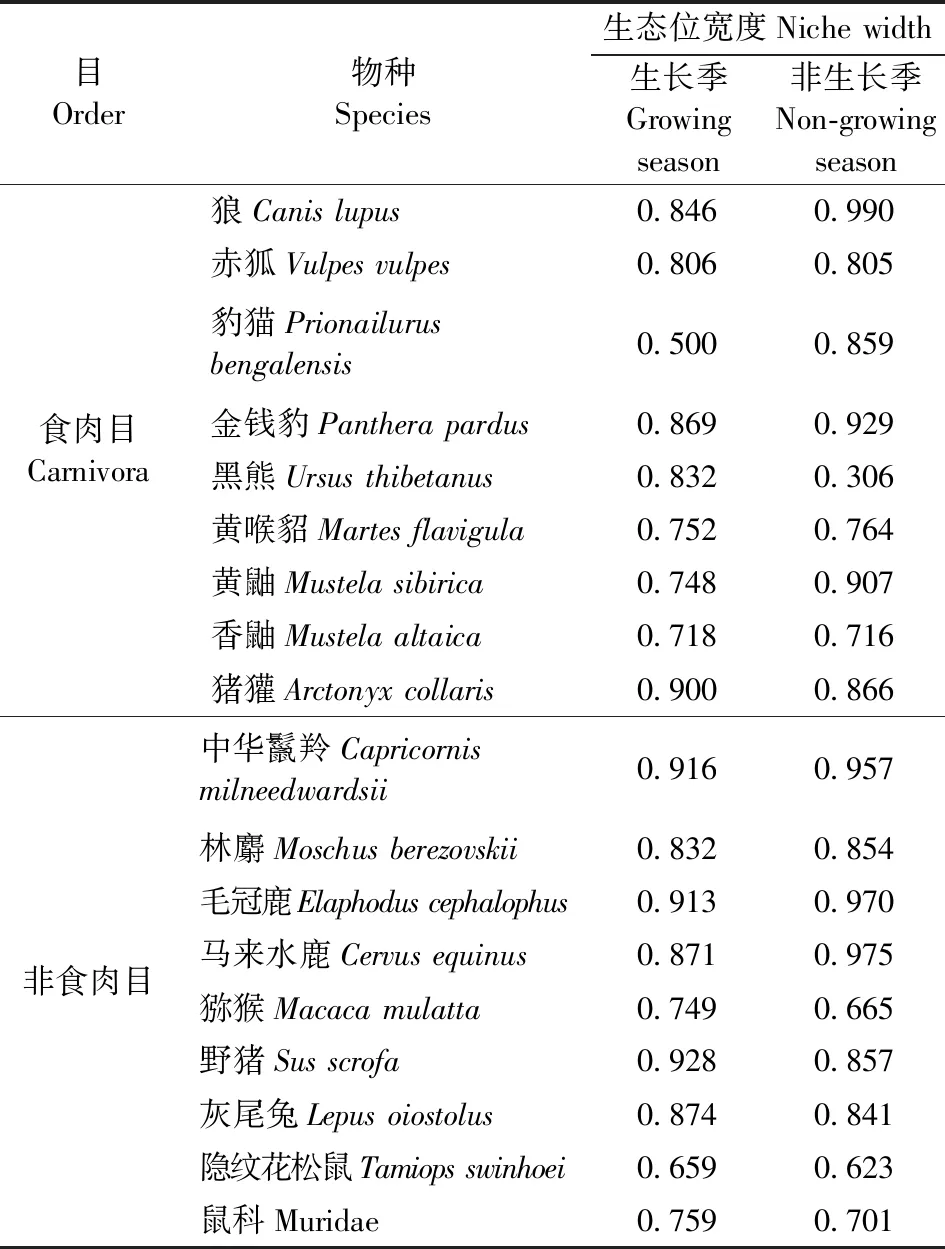
表2 18个物种时间生态位宽度Table 2 Temporal niche width of 18 species
3.3 时间生态位重叠
生长季的时间生态位重叠指数大于0.5的有93对,小于0.5的有60对。其中,灰尾兔与马来水鹿的时间生态位重叠指数最高(0.939),其次是黄鼬与香鼬(0.918)、香鼬与猕猴(0.873),而豹猫与其他动物的生态位重叠指数均较低(表3)。

表3 18个物种在生长季的时间生态位重叠Table 3 Temporal niche overlap of 18 species in growing season
非生长季的时间生态位重叠指数大于0.5的有100对,小于0.5的有53对。其中,毛冠鹿与马来水鹿的时间生态位重叠指数最高(0.933),其次是马来水鹿与狼(0.862)、毛冠鹿与狼(0.853),而黑熊在非生长季与其他动物的时间生态位重叠指数均小于0.5(表4)。
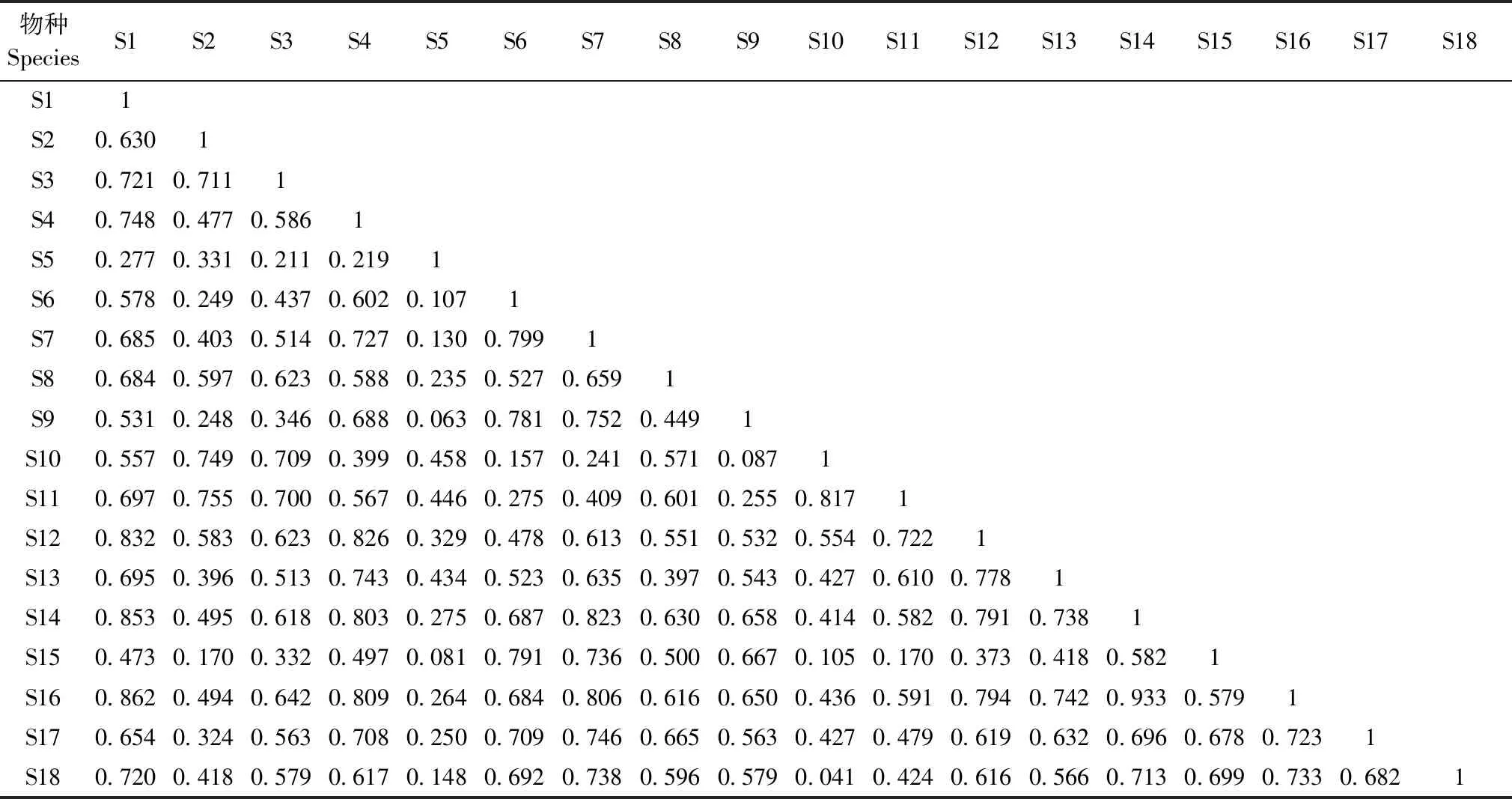
表4 18个物种在非生长季的时间生态位重叠Table 4 Temporal niche overlap of 18 species in non-growing season
4 讨论
多度是在一定区域内特定物种个体的多少,是种群和群落的重要参数,但相对多度不是估计种群个体的绝对数量,而是衡量单位空间内动物数量的相对指标(孙儒泳,2001)。区内豹猫、金钱豹、黑熊、香鼬的相对多度较低,毛冠鹿、马来水鹿、灰尾兔的相对多度较高,这可能与该地区分布的这些物种的相对种群数量有关。
生态位宽度是一个物种所能利用的各种资源总和,时间生态位宽度反映物种的活动强度,生态位宽度越宽,该物种的活动强度越大,也就是说物种在采食等日常行为上花费更多的时间、消耗更多的能量(孙儒泳,2001)。在研究区域中,除黑熊在非生长季的时间生态位宽度小于0.5,其他动物全年均表现出较宽的时间生态位宽度;并且在食物丰富的生长季,狼、赤狐、豹猫、金钱豹的时间生态位宽度小于非生长季,可能是因为其在采食上的时间消耗较少。根据最优采食理论,在食物供应不足时,较宽的生态位可以使消费每个食物单位获得最大回报,因此,动物趋于泛化取食;而在食物丰富的环境中,可能导致生物选择性采食和狭窄的生态位宽度(MacArthur &Pianka,1966;Schoener,1971;MacArthur,1972)。因此,捕食者较宽的生态位宽度从侧面反映出该研究区域内资源的可利用性低,特别是本研究区域内非食肉目动物的生态位宽度较宽,捕食者需花费大量的时间获得资源,且资源可利用性低,其承受的环境压力大。捕食者中个体大的物种往往会选择活动性较小、觅食成功率较大、个体较大的猎物,以减少能耗(孙儒泳,2001)。本研究中,狼、豹猫、金钱豹和黑熊在生长季和非生长季的相对多度均低,但其时间生态位宽度较宽,这说明其在觅食后可能长时间都不会大量活动,以减少能耗,与该结论一致。
其次,生态位重叠显示了种间潜在的相互作用水平(朱曦等,1998)。生态位完全重叠的物种具有较相似的生物学特性和生态适应性,资源丰富时共同利用资源,资源短缺时则发生竞争;生态位完全不重叠的种类,说明其对自然资源的利用存在明显的差异,生态位分化显著(马英等,2011)。因此,物种间生态位重叠小,表明竞争小或者捕食与被捕食关系弱;反之,则表明竞争大或者捕食与被捕食关系强(杨春文等,2008)。本研究结果显示,生长季赤狐与金钱豹、黑熊的重叠指数高,但赤狐与非食肉目的灰尾兔、隐纹花松鼠、鼠科重叠指数高,黑熊与野猪、马来水鹿重叠指数高,金钱豹与马来水鹿、灰尾兔、隐纹花松鼠、鼠科重叠指数高,所以,即使赤狐和黑熊的时间生态位重叠指数高,但其在食性上出现的分化、以及潜在捕食对象时间生态位的分化决定了它们不可能成为竞争对手。而赤狐和金钱豹则在生长季存在着一定的竞争。其次,豹猫与金钱豹、猪獾重叠指数高,豹猫与非食肉目的灰尾兔、隐纹花松鼠、鼠科重叠指数高,而金钱豹与灰尾兔、隐纹花松鼠、鼠科重叠指数高,所以豹猫和金钱豹在生长季存在一定的竞争关系,综上,金钱豹在生长季采食难度较大,因此需要花费较多的时间来觅食,这可能是其时间生态位宽度较宽的原因。
在非生长季中,狼和食肉目的豹猫、金钱豹的重叠指数较高,与非食肉目的马来水鹿、毛冠鹿、中华鬣羚重叠指数高,而豹猫与灰尾兔重叠指数高,金钱豹与中华鬣羚、毛冠鹿、马来水鹿重叠指数高,可看出在非生长季,狼和金钱豹在时间生态位和潜在捕食对象上存在较大的竞争,但是马来水鹿、毛冠鹿、中华鬣羚相互之间重叠指数高且生态位宽度宽,说明它们在数量和活动时间上占优势,同时,亦可作为初级营养级为食肉目动物提供充足的食物。综上所述,狼和金钱豹在非生长季存在竞争,但充足的食物资源和活动时间使它们能够顺利生存。
生态位重叠度低的物种间竞争小,生态位分化大(孙儒泳,2001)。豹猫属于夜行性,而猕猴属于昼行性,两者在活动节律上的差异较大,生态位分化大,故重叠指数低。相反,有蹄类如毛冠鹿、马来水鹿、中华鬣羚的生态位重叠指数较高,反映出种间具有较大的竞争、生态位分化小。本研究对新龙县食肉目兽类及其潜在猎物在时间生态位特征与重叠进行了分析和探讨,可为该区域的生物多样性研究提供本底资料,也为该区域动物保护和管理提供依据。
致谢:感谢给予本研究帮助的各位野外工作人员,以及龙溪虹口国家级自然保护区赵衡提供的帮助。
