粤北石灰岩山地天然次生林优势乔木种土壤-根系-叶片生态化学计量特征
2021-01-22刘强林伟通詹红星温伟良吴永彬
刘强 林伟通 詹红星 温伟良 吴永彬



摘要:以粤北石灰岩山地次生天然林中6种常见优势乔木树种为研究对象,测定了林木根区土壤、根系和叶片的C、N、P等6种营养元素存在水平,运用生态化学计量学方法综合分析了土壤、根系以及叶片的营养元素含量,同时计算分析了计量比指标,并探讨了石灰岩植被生长过程中土壤、根系与叶片等方面的内部影响机制。结果表明:林地土壤C、N、P、K元素含量种间离散程度不大,Ca、Mg元素含量种间离散程度较大;根系营养元素含量由高到低依次为C、Ca、N、K、Mg、P, 根系Ca、Mg元素含量种间离散程度均较大,其他元素含量种间离散程度相对较小;叶片营养元素含量由高到低依次为C、Ca、N、K、Mg、P,营养元素存在水平的大小关系是Ca>K>Mg型,其中叶片C、N、P、K、Mg元素含量种间离散程度不大,而叶片Ca含量种间离散程度较大。经过对叶片成分含量及相应计量比的研究分析,认识到石灰岩植物呈现出P、K较低以及Ca、Mg较高的特點,圆叶乌桕和任豆生长受P元素限制,樟树、朴树、枫香和香叶树生长受N元素限制,6种树种均未受到K元素限制。研究区林地土壤、根系和叶片营养元素水平相互之间的回归分析显示,土壤中的元素的增加或减少对叶片C、Ca和Mg的影响较为显著,叶片C的同化和储存受根系C、N、P、K元素的影响,叶片Ca的吸收利用也与根系C、N、K元素含量息息相关,石灰岩植物通过这样一种特殊的营养调节机制,促使土壤、叶片与根系间的营养转化达到动态平衡。
[关键词] 石灰岩山地;天然次生林;优势乔木树种;生态化学计量学
Abstract: Six dominant tree species in the secondary natural forest of limestone mountain in northern Guangdong Province were studied,the contents of C, N, P, K, Ca and Mg in the soil, root system and leaf of the forest root area were measured. The contents of the nutrient elements in the soil, root system and leaf as well as their metrological characteristics were comprehensively analyzed by the ecological chemometrics method. The synergistic relationship among the soil, root system and leaf in the growth process of limestone vegetation was discussed. The results show that: the content of C, N, P, K elements in forest soil was not dispersed among species, but the content of Ca, Mg elements was dispersed among species; The order of the contents nutrient elements in root system was: C > Ca > N > K > Mg> P, he contents of Ca and Mg in root system were more dispersed among species, while the contents of other elements were less dispersed among species; The order of the contents nutrient elements in leaves was C > Ca > N > K > Mg> P, the characteristics of the contents of nutrient elements were Ca >K > Mg. The contents of C, N, P, K and Mg in leaves were not dispersed among species, but the contents of Ca in leaves were dispersed among species. The analysis of leaf element content and its stoichiometric ratio shows that limestone plants have the characteristics of low P, K and high Ca, Mg; the growth of Sapium rotundifolium and Zenia insignis is limited by P element, the growth of Cinnamomum bodinieri , Celtis sinensis , Liquidambar formosana and Lindera communis is limited by N element, and six tree species are not limited by K element. The regression analysis among soil, root system and leaf nutrient elements shows that the increase or decrease of elements in soil has a significant effect on leaf C, Ca and Mg. The assimilation and storage of leaf C are affected by root C, N, P and K elements. The absorption and utilization of leaf Ca is also closely related to the content of root C, N and K elements. Through such a special mechanism of nutrient regulation, limestone plants promote the dynamic balance of nutrient transformation among soil, leaves and roots.
Key words: Limestone mountains; Secondary natural forest; Dominant tree species; Ecological stoichiometry
在广东省北部,分布着广泛的石灰岩地貌。而该区域自然环境的恶化将使资源持有量不断下降,威胁到当地群众的资源使用需求,自然不利于社会经济的稳定发展。因此,加强对该区域的自然生态监管,提升资源的管理水平,对于维持该区域经济的稳定发展具有重大意义。毫无疑问,森林植被的覆盖情况以及恢复问题,是优化石灰岩区域生态环保工作的关键所在 [1-2]。提升养分的利用水平以及再吸收效率等,属于这种特定地理环境中增强植物适应性,改善所在区域生态环境的关键[3]。而在该领域的研究分析中,生态计量学手段是用来分析生态体系各组分问题,一种分析主要组成元素动态平衡和耦合研究方法[4],在石灰岩区域开展生态计量学的分析,对全面掌握石灰岩植被生态环境下的植物生长发育、营养成分供给等,做好植被的恢复与重建,对改善石漠化的自然环境工作具备十分关键的指导价值[5-6]。 等元素对于植物的生长发育过程非常重要,属于其中的代表性制约因素, 植物机体的生理效应发挥代谢过程的完成,都和这些元素充分相关。生态体系中C和N、P等不同元素的循环变化表现为一个耦合的模式,养分水平的变化直接作用于生态体系的整个运作过程。目前,有关石灰岩地区植物元素以及养分循环的研究处于起步阶段[7-14],且主要集中在西南喀斯特地区,此次课题分析以广东省代表性的石漠化生态系统为研究目标,拟通过研究6个粤北石灰岩天然次生林优势乔木树种的根区土壤、根系和叶片C、N、P、K、Ca、Mg计量学指标,同时研究“土-根-叶”体系C、N、P、K、Ca、Mg之间的关系,揭示粤北石灰岩山地天然次生林植物“土-根-叶”营养元素之间的变化规律,从生态化学计量学角度以期为石灰岩地区森林演替与衰退、生态系统养分供应平衡、自然修复过程等带来更多的借鉴以及实践指导。
1 材料与方法
1.1 研究区概况
研究地位于广东省清远市清新白湾省级自然保护区(24°16′N,112°48′E)。清远处在广东的中部偏北位置,北江中下游,属于珠三角到山区丘的过渡区域,在偏北部位置中分布着代表性的石灰岩地带,中间区域多为低山丘陵,西南方位地形比较低,分布着小面积的平原。是带明显的热带气候,冬天气温比较高,夏季受海洋和地形影响比较温和,年均温度数值在 范围内,无霜时间超过 ,年均降雨量为2139 mm,降水具有明显的波动性,集中在夏季阶段,旱季有时长达数月。白湾为代表性的石灰岩地带,97%属于石山和洼地,仅仅 的土地上有植被、土地覆盖。在植被构成上,重点为人工林、灌木草丛等。
1.2 研究方法
1.2.1 树种选择与样品采集
植物材料根據徐瑞晶[15]对粤北石灰岩植物群落植物多样性与土壤因子研究,选取广东清新白湾省级自然保护区内早期次生林中重要值较大的6种石灰岩森林优势乔木树种,分别为朴树(Celtis sinensis)、樟树
(Cinnamomum bodinieri)、香叶树(Lindera communis)、枫香(Liquidambar formosana)、圆叶乌桕(Sapium rotundifolium)、任豆(Zenia insignis),其中圆叶乌桕、任豆、枫香和朴树为落叶树种,樟树和香叶树则为常绿阔叶树种。
1.2.2 植物和土壤样品采集
在坡向、坡度、海拔等其他因素大体保持一致的地段划定调查范围,在调查区域中,每个树种建立一个半径为10m的圆形样地,共6个样地,并开展常规调查,在选取的6种优势种中,每种分别选取分布相对集中的5株标准木,共计选择标准木30株。对每株标准木完成土壤、根系以及叶片的收集工作,共采集90份样品。
通过专业的剪刀工具获取4个方位以及不同高度位置的枝条,将采集的枝条混匀,摘下叶片,取大约500g枝条装袋,置于80℃烘箱中烘至恒重,随后进行粉碎处理,妥善放置后对叶片 水平进行测定;通过专业设备挖掘从而得到了根部样品,在标准木附近掘取所需的实验对象——树根,同时筛选出尺寸超过 的粗根样以及尺寸小于 的细根,二者比例大概一致,随后装好后转移到 环境中烘干到固定质量,在试试粉碎处理后用来测试 水平。
土壤取标准木根系1m范围内的表层土(0-20cm),混匀后取1kg,每株标准木取1个土样,共采土样30个。土样在室温条件下自然风干,风干后去除根、石头等杂物,研磨,过0.25mm筛,保存好用于测定 含量水平。
1.2.3 分析方法
用高温外热重铬酸钾氧化-容量法测定土壤有机C(因为土壤所含有的无机碳十分微量,可以忽略不计,所以本文通过有机碳来代表全碳含量)含量,通过开氏-蒸馏滴定法测定全N的含量,氢氧化钠熔融-钼锑抗比色法测定全P含量,氢氧化钠熔融-火焰原子吸收分光光度法测定全K含量,三酸消解-火焰原子吸收分光光度法测定全Ca、Mg含量;在植物叶片与根系元素含量测定上,高温外热重铬酸钾氧化-容量法测定全C含量,硫酸-双氧水消煮-蒸馏滴定法测定全N含量,硫酸-双氧水消煮-钒钼黄比色法测定全P含量,全Ca、Mg含量用干灰化-稀盐酸溶解-火焰原子吸收分光光度法测定[16]。而火焰分光光度计型号:Z-2300(来源于日立企业),相关光度计款式: (南方精密仪器设备企业)。
1.2.4 数据分析
不同树种叶片、根系以及根区土壤的 、Mg元素含量均采用 软件分析并作图;采用SPSS 19.0软件对实验过程中掌握的资料信息展开方差研究以及多重对比( 法),后续完成关联度研究,回归处理。
3 结果与分析
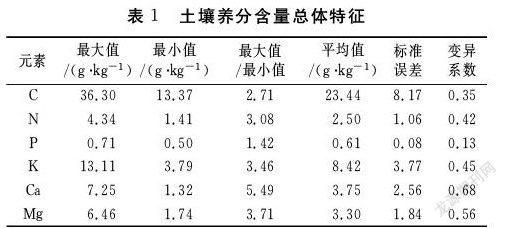
3.1林地土壤养分含量特征
由表可以看出,6种树种土壤全C含量均值为23.44g/kg,标准差为8.17,变异系数为0.35,属于中等变异;土壤全N含量的平均值为2.50g/kg,根据国家部门第二次土壤普查划分和设定的标准[17],属于一级(大于2 g/kg)水平,标准差为1.06,变异系数为0.42,属于中等变异;土壤中全P含量平均值为0.61g/kg,属于三级(0.6-0.8g/kg)水平,标准差为0.08,变异系数为0.13,属于弱变异;土壤中全K含量平均值为8.42g/kg,属于五级(5-10g/kg)水平,含量较低,标准差为3.77,变异系数为0.45,属于中等变异;土壤中全Ca和Mg含量平均值分别为3.75g/kg和3.30g/kg,一般土壤中Ca 的含量为13.7 g/kg ,Mg 为6.0 g/kg,可见林地土壤Ca、Mg含量均偏低,二者的变异系数分别为0.68和0.56,均大于0.5,属于强变异。
3.2 根系养分含量特征
由表2可知,根系C含量最高,平均值为420.87g/kg,标准差为25.43,最大值/最小值和变异系数均较小,属于弱变异;根系P含量最低,变异系数为0.38,属于中等变异;根系Ca、Mg含量的变异系数均较大,属于强变异,说明根系Ca、Mg含量种间差异较大。根系元素含量平均值从大到小排序为:C>Ca>N>K>Mg>P。
3.3 叶片养分含量总体特征
3.3.1 叶片营养元素含量
由表3可以看出,叶片平均含量大于20 g·kg-1的元素有C、N、Ca,以叶片C含量最高,平均值为426.84g/kg;其余叶片P、K、Mg三种元素平均含量均小于大于10g/kg,以叶片P含量最低,平均值仅为1.56g/kg;叶片元素含量平均值从大到小排序为C>Ca>N>K>Mg>P。结果显示,6种优势乔木树种叶片营养元素含量特点对应为 型。离散程度越大则变异系数也越大,离散程度的顺序可总结为Ca>Mg>N>K>P>C,其中Ca的变异系数最大,为0.62,其余5种营养元素的变异系数均小于0.5。基于最大、最小数值的对比分析,Ca的值最大,为7.63,可见Ca元素的物种变异程度相对较大,因此树种叶片存在差异时,其表现出来的对Ca元素的吸收情况具有不同。C元素的变异系数以及最大值/最小值均是各元素中最小的,树种间变异程度相对较小,不同树种叶片对C元素的储蓄具共性。
3.3.2 叶片营养元素含量间的比值
由表4可知,叶片营养元素比值不同树种间也存在差异,其中叶片 N/K比值均以圆叶乌桕最高,C/N比值以枫香最高,K/P比值以樟树最高, 和 比值以朴树最高,K/Mg比值以香叶树最高。
结合表4 能够发现,粤北石灰岩优势乔木树种和全球陆地植物平均水平进行对比分析,前者叶片N/P、 Ca/K、Ca/Mg比值都超過后者,而C/N、K/Mg比值前者明显不及后者,K/P比值两者大体一致。此外,结合表5还能观察到,虽然同一比值不同种间具有相应的差别,不过变异系数值均较小,意味着叶片营养元素的分布是稳定和协调的状态。粤北石灰岩优势乔木树种叶片元素之间的计量关系,表明了P、K 元素成分较少而 元素含量太高的自然环境表现,另外也体现了该范围内的植物为了在一定自然环境下适应下来,而对自身的元素含量进行调控与修正。
3.4 土壤、根系和叶片之间C、N、P、K、Ca、Mg含量的关系分析
3.4.1土壤和根系C、N、P、K、Ca、Mg含量的关系分析
选择x1、x2、x3、x4、x5、x6分别代表林地土壤C、N、P、K、Ca、Mg含量(g/kg),y1、y2、y3、y4、y5、y6分别代表植物根系C、N、P、K、Ca、Mg含量(g/kg),通过 软件对植物根系养分含量和土壤养分含量展开回归分析研究。随后围绕所创建的方程展开显著性分析,观察到植物根系N、K、Mg和土壤元素含量的回归模型均未达到显著状态,表明土壤养分含量未对植物根系N、K、Mg含量产生显著影响,而植物根系C、P、Ca与土壤养分含量回归模型均达到了显著水平(P<0.05):
y1=1.737+1.955x1-13.767x2+14.326x3-0.239x4+0.064x5+1.032x6
y3=0.035-0.004x1+0.026x2+0.828x3+0.002x4-0.001x5+0.028x6
y5=0.606+0.330x1-3.903x2+3.227x3-0.002x4+0.949x5-0.109x6
对以上3个方程不同的回归指标展开显著性分析,剔除不显著项后,得到回归模拟方程,见表5。由表5可知,植物根系C 水平和土壤C水平满足明显的正相关,和N水平满足高度的负相关;植物根系P水平和土壤P水平满足高度的正相关;植物根系Ca水平和土壤N水平满足高度的负相关,和土壤Ca水平满足高度的正相关(P<0.05)。
3.4.2林地土壤和叶片C、N、P、K、Ca、Mg含量的关系分析
选择x1、x2、x3、x4、x5、x6分别代表林地土壤C、N、P、K、Ca、Mg含量(g/kg),y1、y2、y3、y4、y5、y6分别代表植物叶片C、N、P、K、Ca、Mg含量(g/kg),通过 软件对叶片养分含量和土壤养分含量的关系展开回归分析研究。随后围绕所创建的方程展开显著性分析,观察到叶片 和土壤元素含量回归模型均未达到显著层次,表明土壤元素没有对叶片N、P、K含量产生显著影响,而叶片C、Ca、Mg含量和土壤元素含量回归模型,则实现了显著水平(P<0.05):
y1=543.403-2.478x1+24.786x2-71.088x3-10.608x4-0.346x5+4.350x6
y5=-14.530+2.297x1-20.055x2+8.982x3+5.909x4+0.673x5-5.967x6
y6=3.448+0.120x1-0.814x2-1.850x3+0.124x4+0.008x5-0.556x6
对上面3个方程的不同系数展开显著性测验分析,剔除不显著项后,得到回归模拟,模拟方程见表6。由表6可知,叶片C含量和土壤环境K含量满足高度的负相关,Ca含量和土壤C、K含量都满足高度的正相关,Mg含量和土壤Mg水平满足高度的负相关(P<0.05)。
3.4.3根系和叶片C、N、P、K、Ca、Mg含量的关性分析
选择x1、x2、x3、x4、x5、x6分别代表植物根系C、N、P、K、Ca、Mg含量(g/kg),y1、y2、y3、y4、y5、y6分别代表植物叶片C、N、P、K、Ca、Mg含量(g/kg),通过 软件对根系养分含量和叶片养分含量的关系展开回归分析研究。随后在方程中完成显著性测试分析,观察到叶片N、P、K、Mg和根系元素含量回归模型均未达到显著层次,表明根系养分含量没有对叶片N、P、K、Mg含量产生显著影响,而叶片C、Ca含量和根系元素含量回归模型,均实现了显著水平(P<0.05):
y1=544.005-5.482x1+56.390x2-120.628x3-10.750x4-0.202x5+11.002x6
y5=-12.283+3.291x1-31.001x2+25.024x3+5.703x4+0.486x5-7.665x6
对上面2个方程的不同系数展开显著性测验分析,剔除不显著项后,得到回归模拟,模拟方程见表7。由表7可知,叶片C含量和根系C、P、K含量均呈显著负相关,同时和根系N含量满足高度的正相关;叶片Ca含量和根系C、K含量都满足高度的正相关,而和根系N含量满足高度的负相关(P<0.05)。
4 结论与讨论
4.1 “土-根-叶”系统养分特征比较及营养元素间关系探讨
根系营养的维持,和土壤环境之间高度相关。根系所出现的生理变化以及内在机制都是土壤环境下实现的,二者关系密切,土壤养分水平对根系的生长具有关键作用。本研究区植物根系C含量要明显高于土壤C含量,但和土壤C含量具有高度的正比例关联( ),表明植物根系虽然不从土壤中直接吸收C元素,但土壤中C元素的储量将直接作用于植物的吸收过程,进而影响植物同化C的能力。此外,本研究区植物根系P与土壤P满足高度的正相关关系( ),根系Ca和土壤Ca同样满足高度的正相关关系,根系P、Ca含量也明显高于土壤P、Ca的含量,土壤是植物根系营养的主要来源,而相比之下,土壤中的营养成分要“贫瘠”一些,这意味着根系具备集中和涵养元素的作用。整体而言,土壤和根系不同元素方面更多地是正相关状态,较少地表现为负相关性,这与徐露燕[19]的研究认为土壤营养成分的提升或者下降,都会影响根系环境中部分元素的增加的结论相一致。土壤以及叶片等在养分变化中具备十分关键的功能,已有的研究[20-22]也表明了两者之间存在一定的联系。本研究中,叶片N、P、K含量和土壤元素含量的关联性并不突出,这与大部分研究认为叶片N、P、K 含量与土壤养分含量相关性总体较好的结论[20-22]有所不同,说明元素吸收机理的复杂性,盲目施肥并不一定带来促进植物生长的效果。另一方面,土壤中的元素的增加或减少对叶片C、Ca和Mg的影响较为显著,这可能是由于石灰岩区植物的营养吸收特性造成的。叶片是光合效应与呼吸过程的关键场所,同样是保存与固定成分的典型场合,养分重点是来自于空气,同样有很大一部分经过根系吸收,因此叶片与根系成分满足相应的联系。本分析表明,叶片 Ca、Mg的含量均要高于根系,这是因为叶片是起同化作用的器官,有着旺盛的新陈代谢功能,所以养分含量也相应较高。相关性方面,叶片C含量和根系C、P、K含量满足高度的负相关状态,不过和根系N含量满足高度的正相关状态;叶片Ca含量和根系C、K含量满足高度的正相关状态,和根系N含量满足高度的负相关状态(P<0.05)。总体看来,叶片C的同化和储存受根系C、N、P、K元素的影响,叶片Ca的吸收利用也与根系C、N、K元素含量息息相关,石灰岩植物通过这样一种特殊的营养调节机制,促使叶片与根系间的营养转化达到动态平衡,以此来维持植物在石灰岩地区的正常生长。
4.2 粤北石灰岩山地优势树种与其他地区植物叶片养分特征比较
本研究中,树种叶片营养元素含量特点是Ca>K>Mg型特征,这和前人在喀斯特地区所做的研究[7,23-25]基本一致。本研究叶片N的质量分数高于珠三角地区[26]、黔中地区[9]和全国陆地植物[27],但要低于同为喀斯特地区的茂兰[7],大致处于全球尺度植物的平均水平[28];叶片P的质量分数与珠三角地区[26]、茂兰[7]、黔中地区[9]和全国陆地植物[27]水平大致相同,略低于全球尺度植物[28]的平均水平,具有低P的特点。K元素在植物体内的生理机制以及循环代谢中发挥着关键性的功能,是植物体内含量最为丰富的阳离子,也是生物必需的营养元素[29]。本研究区植物叶片K的质量分数低于茂兰地区[7]、黔中地区[9]以及全球尺度植物的平均水平[18],K含量較低,这和阎凯等[30]认为植物在生境恶劣地区会选择性地吸收富集更多K元素来应对贫瘠环境的结论并不一致。Ca属于植物生长的关键元素,它对膜结构的稳定性有重要影响,Mg作为叶绿素的必要成分,缺镁时,叶绿素的形成受到阻碍,光合作用的功能也会受到阻碍[31]。本研究区植物Ca的质量分数与茂兰喀斯特地区[7]水平大致相同,但要显著高于全球尺度植物的平均水平[18],叶片Ca元素非常丰富;Mg的质量分数虽然要低于茂兰地区[7],但也要高于全球尺度植物的平均水平[18],叶片Mg元素也较为丰富。总体来说,本研究区的树种叶片具有低P、K,高Ca、Mg的特点,这和罗绪强等[7]在茂兰喀斯特地区的研究结果一致,反映了低P、K,高Ca、Mg是石灰岩植物在养分吸收特性上的共性,另外也体现了石灰岩区植物为适应外部环境而做的努力,在元素上的协调机制。
4.3各树种叶片化学计量特征以及对植物生长的限制性因子判断
植物叶片的C/N与C/P说明植物吸收过程中所具备的同化C的水平,在一定程度上可以体现出植物的营养利用水准,也就是说C积累效率和保存能力,与N和P元素的供应高度相关[32]。本研究6种树种中,枫香、香叶树和樟树的C/N比值分别为30.32、24.97和22.79,C/P比值分别为209.41、263.79和306.66,都超过了世界范围内的均值[18]22.5与 ,也表明这三种树种具有较全球水平更高的营养利用效率。
自然环境下,N与P元素的分布减少,使得N与P演变为陆地环境下作用于植物生长的制约性元素[27]。叶片的N/P比值可以体现植物生长的制约性因子状态,满足 时,则认为是受到N的制约,满足 时,则表明是受到P含量制约,当N/P处在 范围内,意味着N与P独立或者共同作用于植物的生长发育[33]。结合本研究6种树种叶片N/P比值,圆叶乌桕叶片和任豆N/P比值分别为18.04和16.13,均大于16,同样超过全国均值[27](14.1)与世界平均水准[34](13.89),表明在物种水平上分析,圆叶乌桕和任豆的生长主要受P元素限制;樟树、朴树、枫香和香叶树叶片N/P<14,表明生长受N元素限制。有研究指出,根据叶片N/P比值分析相关制约因素运用在群落水平上的状况[35],此次课题分析掌握到的资料信息仅仅在物种水平,小尺度水平上来研究具有明显的不足,后续应该选取更大样本来印证本研究的结果。根据K元素限制阈值的划分标准[36]:当 , 时,植物的生长受K元素限制,结合本研究6种树种叶片N/K和K/P比值,6种树种均未受到K元素限制。
[参 考 文 献]
[1] 喻理飞,朱守谦,叶镜中,等.退化喀斯特森林自然恢复过程中群落动态研究[J].林业科学,2002, 38(1):1-7.
[2] 魏兴琥,李森,罗红波,等.粤北石漠化过程土壤与植被变化及其相关性研究[J].地理科学,2008,28(5):662-666.
[3] 俞月凤,何铁光,彭晚霞,等.喀斯特峰丛洼地不同类型森林养分循环特征[J].生态学报,2015,35(22):7531-7542.
[4] ZHANG J H, ZHAO N, LIU C C, et al. C:N:P stoichiometry in China’s forests: From organs to ecosystems [J]. Functional Ecology,2017
[5] 皮发剑,舒利贤,喻理飞,等.黔中喀斯特10种优势树种根茎叶化学计量特征及其关联性[J].生态环境学报,2017,26(4):628-634.
[6] 杜家颖,王霖娇,盛茂银,等.喀斯特高原峡谷石漠化生态系统土壤C、N、P生态化学计量学特征[J].四川农业大学学报,2017,35(1):45-51.
[7] 罗绪强,张桂玲,杜雪莲,等.茂兰喀斯特森林常见钙生植物叶片元素含量及其化学计量学特征[J]. 生态环境学报, 2014(7):1121-1129.
[8] 汪舒雅,喻理飞,黄宗胜.黔中喀斯特区不同光合功能群树种凋落叶分解特性研究[J].西部林业科学, 2014(6):139-142.
[9] 皮发剑,袁丛军,喻理飞,等.黔中天然次生林主要优势树种叶片生态化学计量特征[J]. 生态环境学报, 2016, 25(5):801-80
[10] 贺静雯,刘颖,余杭,等.干热河谷优势灌木养分重吸收率及其C:N:P化学计量特征[J].北京林业大学学报,2020,42(01):18-26.
[11] 刘立斌,钟巧连,倪健.贵州高原型喀斯特次生林C、N、P生态化学计量特征与储量[J].生态学报,2019,39(22):8606-8614.
[12] 吴丽芳,王紫泉,王妍,等.喀斯特高原不同石漠化程度土壤C、N、P化学计量特征和酶活性的关系[J].生态环境学报,2019,28(12):2332-2340.
[13] 喻阳华,钟欣平,王颖.喀斯特高原峡谷区土壤大/中/微量元素的生态化学计量特征[J].西南农业学报,2019,32(9):2068-2072.
[14] 喻阳华,钟欣平,李红.黔中石漠化区不同海拔顶坛花椒人工林生态化学计量特征[J].生态学报,2019,39(15):5536-5545.
[15] 徐瑞晶.粤北石灰岩植物群落多样性与土壤因子研究[D].广州:华南农业大学,2014.
[16] 鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社,2000.
[17] 李志洪,赵兰坡,窦森.土壤学[M].北京:化学工业出版社,2005.
[18] 泰兹,奇格尔.植物生理学[M]. 4版.宋纯鹏,王學路,等译.北京:科学出版社, 2009:60-75.
[19] 徐露燕.湘潭锰矿区不同年龄栾树林土壤、根系和叶片C、N、P化学计量特征[D]. 长沙:中南林业科技大学, 2014.
[20] 涂淑萍,叶长娣,王蕾,等.黄竹叶片营养与土壤肥力及产量的相关研究[J].江西农业大学学报, 2011, 33(5):918-923.
[21] 李广会,郭素娟,邹锋,等.板栗叶片营养与土壤养分的动态变化及回归分析[J].中南林业科技大学学报, 2012, 32(9): 41-46.
[22] 邱权,李吉跃,王军辉,等.柴达木盆地不同居群白刺叶片营养与土壤养分的回归分析[J].西北农林科技大学学报自然科学版, 2014(11):121-128.
[23] 周運超.贵州喀斯特植被主要营养元素含量分析[J].山地农业生物学报, 1997(1):11-16.
[24] 杨成,刘丛强,宋照亮,等.贵州喀斯特山区植物营养元素含量特征[J].生态环境学报,2007, 16(2):503-508.
[25] 旷远文,温达志,闫俊华,等.贵州普定喀斯特森林3种优势树种叶片元素含量特征[J].应用与环境生物学报, 2010, 16(2):158-163.
[26] 吴统贵,陈步峰,肖以华,等.珠江三角洲3种典型森林类型乔木叶片生态化学计量学[J].植物生态学报, 2010, 34(1):58-63.
[27] Han W, Fang J, Guo D, et al. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J]. New Phytologist, 2005, 168(2):1007-1019.
[28] Elser J J, Fagan W F, Denno R F, et al. Nutritional constraints in terrestrial and freshwater food webs[J]. Nature, 2000, 408(6812):578-580.
[29] 郑炳松,程晓建.钾元素对植物光合速率、Rubisco和RCA的影响[J].浙江农林大学学报,2002,19(1):104-108.
[30] 阎凯,付登高,何峰,等.滇池流域富磷区不同土壤磷水平下植物叶片的养分化学计量特征[J].植物生态学报,2011,35(4):353-361.
[31] 潘瑞炽,王小菁,李娘辉.植物生理学[M].北京:高等教育出版社, 2012.
[32] Huang J, Wang X. Leaf nutrient and structural characteristics of 32 evergreen broad-leaved species[J]. Journal of Eastchina Normal University, 2003(1):92-97.
[33] Koerselman W. The Vegetation N:P Ratio: a New Tool to Detect the Nature of Nutrient Limitation[J]. Journal of Applied Ecology, 1996, 33(6):1441-1450.
[34] Reich P B, Oleksyn J, Tilman G D. Global Patterns of Plant Leaf N and P in Relation to Temperature and Latitude[J]. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(30):11001-11006.
[35] Güsewell S, Koerselman W. Variation in nitrogen and phosphorus concentrations of wetland plants[J]. Perspectives in Plant Ecology Evolution & Systematics, 2002, 5(1):37-61.
[36] Venterink H O, Wassen M J, Verkroost A W M, et al. Species Richness-Productivity Patterns Differ between N-, P-, and K-Limited Wetlands[J]. Ecology, 2003, 84(8):2191-21.
