SAM和GSH发酵联产过程中合成关键酶的作用机制
2021-01-18马瑞何鑫
马瑞 何鑫
(宁夏回族自治区森林病虫防治检疫总站,宁夏 银川 750001)
引言
SAM在生物体内是重要的中间代谢物质,参与多种生化反应,在临床上应用十分广泛,在药用方面有很大的应用前景,受到越来越多的生命科学领域和医药领域人士的关注。
GSH是细胞内非蛋白巯基的主要组成部分,在生物体内具有维持适宜氧化还原环境、作为抗氧化剂保护酶和蛋白的硫氢基、防止自由基的氧化损伤等重要功能,使其在临床、食品、体育运动等多个领域有广泛的应用前景[1]。目前生物合成法即酶法和微生物发酵法较受研究者关注,而微生物发酵法因其发酵微生物(细菌或酵母)培养简单,生产工艺和方法不断改善,成为了目前工业化生产SAM和GSH的普遍方法[2]。
SAM和GSH均是含硫化合物代谢网络中的重要节点物质,具有共同的代谢途径[3]。因此研究不同可控条件下的SAM和GSH联合高产,具有重要的理论和生产价值。ATP的存在为SAM和GSH合成提供了能量,而ATP主要是由细胞呼吸作用产生的。因此可以通过改变发酵条件影响细胞的呼吸,进而影响细胞的能量代谢途径,最终实现GSH和SAM的联合高产。本文通过设置不同溶氧控制条件(15%、30%、45%、60%以及对照组恒转速)探究不同溶氧条件对发酵联产产量影响,对比得出最佳溶氧控制条件。
1 材料与方法
菌种为C.utilis CCTCC M 209298,由苏州大学基础医学与生物科学学院微生物工程实验室筛选并保存。
1.1 培养基
种子培养基(g·L-1):葡萄糖20,蛋白胨10,酵母粉10,pH6.0。
发酵培养基(g·L-1):葡萄糖35,NH4SO410,KH2PO412.3,MgSO40.05,CaCl20.05,L-Met 4.6。
1.2 培养方法
1.2.1 一级种子培养
-70℃冷冻保藏的种子活化后接入准备好的一级种子培养基中,摇床转速200r·min-1,恒温30℃培养24h。
1.2.2 二级种子培养
以10%(v/v)接种量将新鲜种子液接种入二级种子培养基中,恒温30℃,200r·min-1摇床培养20h。
1.2.3 分批发酵培养
将新鲜二级种子液按10%接种量接入装液量为3L的5L在位发酵罐中,控制温度30℃,初始通气量1vvm,pH5.0(在pH电极的检测下,通过2mol·L-1的H2SO4和2mol·L-1的NaOH自动流加控制),初始搅拌转速350r·min-1。发酵过程中,当溶氧降低到一定值后,通过手动调节转速和通气量来控制恒定发酵溶氧水平,培养时间30h。
1.3 取样方法
取样自发酵起始0h、3h、6h、9h、12h、15h、18h、21h、24h、27h、30h从发酵罐中吸取发酵液50mL至量筒中,吹打混匀后分装至离心管中,10mL 3管,5mL 4管。低速离心机3500rpm离心10min后,收集2mL上清进行发酵液残糖测定,酵母泥放入-70℃冰箱保存。
1.4 分析方法
1.4.1 细胞干重(DCW)[4]
取10mL发酵罐培养液,3500rmp离心10min,蒸馏水洗涤3次后进行离心,收集菌体沉淀,70℃烘干至恒重后称量。
1.4.2 残糖浓度测定[5]
取转化后的菌液离心,取10mL上清液,适当稀释后取2mL,加入1.5mL DNS,沸水浴5~7min,再冷水浴冷却至室温,定容至25mL,520nm处测OD值,通过标准曲线对比得出残糖浓度。
1.4.3 GSH测定[6]
离心所得酵母泥在-70℃冷冻过夜后,第2天用40%酒精25mL摇床30℃振荡萃取2h后,3500r·min-1离心10min,取上清进行胞内GSH测定。获得的样品胞内稀释25倍后采用DTNB-谷胱甘肽还原酶循环法进行定量测定。
在2mL体积的比色皿中按顺序依次加入100μL DTNB,700μL NADPH和经适当稀释的样品200μL(胞内稀释25倍,胞外稀释100倍)。室温下加入10μL谷胱甘肽还原酶启动反应,在412nm处测定反应体系的初始OD值和反应3min后的OD值。
1.4.4 SAM的提取和测定
10mL发酵液离心取细胞沉淀,向沉淀中加10mL 0.35M稀硫酸,4℃静置萃取2h,3500r·min-1离心10min,上清液经0.22μm膜过滤即得样品(约需1mL)。得到的样品采用HPLC法进行分析。色谱柱为反相SunFireC18柱(4.6nm×250nm),流动相为0.5mol·L-1的甲酸铵(化学纯,用甲酸溶液调pH至4.0)溶液,流速0.8mL·min-1,检测波长254nm,柱温25℃,检测池温度30℃,进样量20μL[7]。
1.4.5 辅因子定量分析
离心得到的酵母泥,液氮处理1min后放入-70℃超低温冰箱中保存。将湿细胞重新悬浮在5mL 0.2mol·L-1磷酸缓冲液(pH7.0)中,超声波破碎10min(功率22%,破碎10s,间隔10s),离心后得到的上清液作为待测样品,采用HPLC法检测辅因子含量[8]。
1.5 酶活测定
1.5.1 γ-谷氨酰半胱氨酸合成酶(γ-GCS)酶活测定
微量定磷法,使用南京建成γ-GCS检测试剂盒(A091-2)测定。
1.5.2 SAM合成酶酶活测定
发酵过程中SAM合成酶的酶活水平是通过测定体外酶促反应液的SAM含量来监控[9]。
2 结果与讨论
2.1 不同溶氧控制条件对细胞生长和SAM、GSH产量影响
对于发酵,溶解氧既属于营养因素,又属于环境因素,特别是对于本次实验SAM和GSH的生产代谢过程中,DO的改变会影响产朊假丝酵母呼吸链中的氧化还原,同时也会对酵母细胞的生长和SAM、GSH形成产生影响。由于在发酵的不同阶段,细胞对溶氧的需求水平并不相同,恒定转速的发酵很容易造成发酵过程中溶氧的不足或过剩,使得菌体发生厌氧代谢或氧中毒,对发酵产生不利的影响[10]。为此本实验将设定恒定溶氧,研究恒溶氧对发酵过程的影响,探究菌体生长的最佳溶氧水平。
为了探究不同溶氧条件下对SAM和GSH产量代谢影响,本实验将溶氧条件分别设定为15%恒溶氧、30%恒溶氧、45%恒溶氧、60%恒溶氧以及对照组恒转速(不控溶氧)5个批次来分别对比细胞生长情况(见图1)。由图1可见,各溶氧控制条件细胞量均在15h达到最大值,此时葡萄糖基本耗尽;各溶氧条件下,细胞量水平相似,在发酵前15h内,30%恒溶氧发酵的细胞量水平较高;此时细胞合成已经停止,但因为一些中间代谢物质的存在,目标产物SAM和GSH仍在继续合成[2]。由图2可见,对GSH和SAM合成有利的溶氧控制条件分别为30%恒溶氧和60%恒溶氧;在30%恒溶氧的控制条件下,GSH的积累量在18h达到了最大值376.6mg·L-1,与实验对照组恒转速相比提高了46%;在60%恒溶氧的控制条件下,SAM的积累量在30h时达到最大值243.8mg·L-1,与实验对照组恒转速条件相比提高了26%。
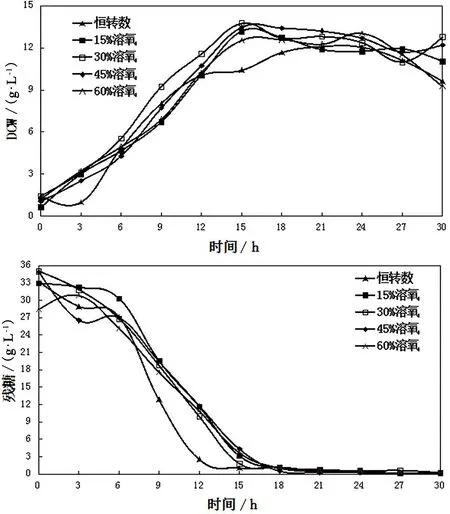
图1 不同溶氧下细胞量和残糖量

图2 不同溶氧下SAM、GSH以及两者联合产量
进一步将SAM和GSH联产积累量作图分析可以看出,综合2种目标产物的联产量,在30%恒溶氧控制的条件下产量最高,在27h达到最大值594mg·L-1,与实验对照恒转速组相比提高了38%。为了更加直观地关注GSH、SAM发酵过程中的生产效率,本实验将发酵过程中的各指标参数列于表1。从表中各指标参数综合对比来看,30%恒溶氧的参数与各实验组参数指标相对比,其各项发酵指标均较为理想。对于此实验结果,以下将从GSH、SAM的发酵过程中的酶活角度来阐述和分析。
2.2 不同溶氧控制条件下GSH和SAM发酵过程分析
在上一节中,通过图表直观地了解到30%恒溶氧发酵条件下GSH和SAM的联产量达到最高,与对照组相比提高了38%。在发酵过程中,与GSH、SAM合成紧密联系的是γ-谷氨酰半胱氨酸合成酶和SAM合成酶,其酶活程度直接决定了GSH和SAM的产量高低。从图2可见18h后GSH几乎不再合成,GSH胞内含量逐渐下降,分析原因有2点:GSH合成速度逐渐慢于GSH降解速度,使得GSH胞内含量逐渐趋于平缓甚至下降;培养基内谷氨酸供给缺乏,谷氨酸是GSH合成必须的前体物质。在对不同溶氧条件下γ-GCS酶活数据分析(图3),在15h时酶活达到最高。此时细胞的葡萄糖基本耗尽,酶活逐渐降低,GSH的积累量逐渐持平甚至降低。从图3中可以看出,30%的恒溶氧条件下,γ-GCS的酶活明显高于其它实验组,由此可以得出30%恒溶氧条件的GSH产量高是由于γ-GCS的酶活最高。
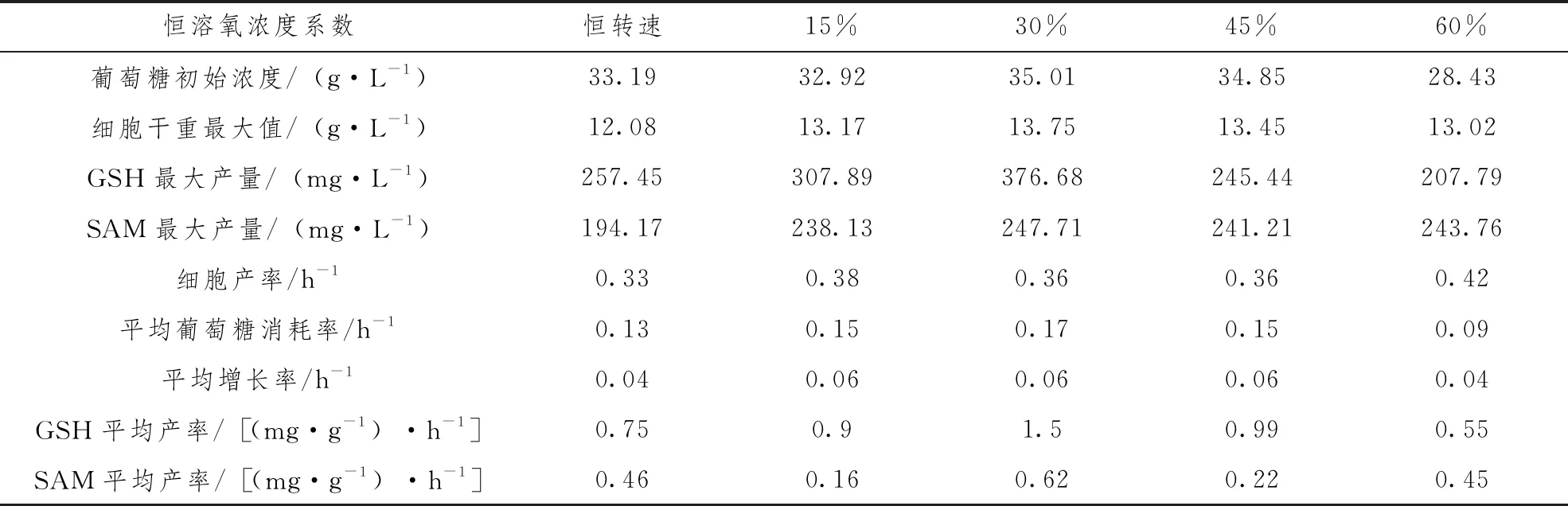
表1 不同溶氧条件下GSH和SAM联产发酵过程参数
与SAM合成密切相关的是SAM合成酶,在整个发酵过程当中60%恒溶氧发酵的胞内含量高于其它溶氧控制条件的实验组,与起始胞内含量相比,SAM的胞内提高了104%。综合其它组的SAM合成规律发现细胞在15~18h之后合成速度明显加快,SAM胞内含量急速上升,这与SAM合成酶的联系十分紧密。从图3中可知15h后SAM合成酶的酶活由之前的下降趋势逐渐开始上升,随着其上升的趋势,SAM在胞含量逐渐提高。这是由于15h之后,GSH基本不再合成,而SAM是GSH合成中的前体物质,GSH不再合成,SAM的消耗量降低,由此得到了一定量的积累。在30%恒溶氧的发酵过程当中,γ-GCS酶活始终保持着明显高于其它组的活性,其GSH合成十分旺盛,使得其相应的SAM消耗也高于其它实验组,因此在SAM合成产量方面并没有优于其它组。从图3可见,30%恒溶氧18h后SAM合成十分旺盛,与起始SAM胞内含量相比提高了300%。
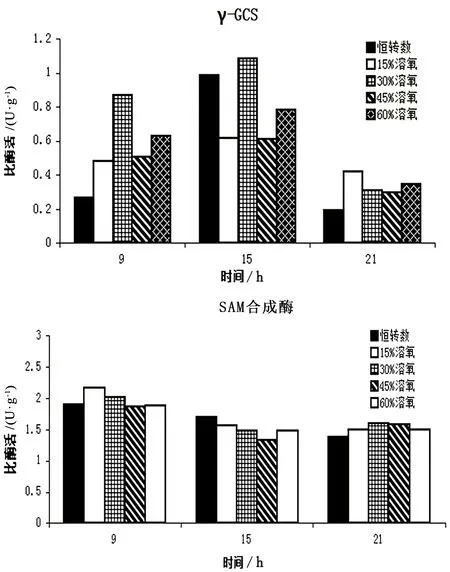
图3 不同溶氧条件下的γ-GCS、SAM合成酶活性
从γ-GCS和SAM合成酶角度可分析得出30%恒溶氧发酵的SAM和GSH联产量之所以高于其它实验组,是由于其γ-GCS酶活明显高于其它组别,并且SAM胞内含量处于上升阶段时SAM合成酶亦处于活跃阶段,酶活程度的高低直接影响到目标产物产量高低。
3 结论
本实验过程中,通过对实验数据的分析得出由于15h葡萄糖基本耗尽,细胞在这时停止合成,最高生物量达到13.75g·L-1。
不同溶氧条件下,SAM和GSH联产发酵产量不同,30%恒溶氧最利于GSH合成,较对照组提高了46%,在18h达到376.4mg·L-1;60%恒溶氧最利于SAM合成,较对照组提高了26%,在30h达到243.8mg·L-1。综合2者合成量进行对比,30%恒溶氧最利于SAM、GSH发酵联产,较对照组产量提高了38%,在27h达到594mg·L-1。
SAM、GSH合成关键酶酶活直接影响SAM、GSH产量,酶活高有利于SAM和GSH合成。对γ-GCS酶活数据分析,在15h时酶活达到最高,30%恒溶氧的γ-GCS酶活明显高于其它实验组,使得其GSH含量最高。SAM合成随SAM合成酶的酶活升高而加速,并且由于18h后GSH合成逐渐减慢,SAM消耗减少,因此积累量增多。
