烟草抗青枯病突变体153-K的抗性遗传及与农艺性状的关系
2021-01-18牛文利巫升鑫余文程崖芝杨兴有余祥文陈志华刘勇丁安明孙玉合王卫锋
牛文利 巫升鑫 余文 程崖芝 杨兴有 余祥文 陈志华 刘勇 丁安明 孙玉合 王卫锋
摘要:烟草青枯病是一种细菌性病害,严重影响烟叶生产,筛选抗青枯病的烟草种质并解析其抗性遗传效应对指导抗病育种具有重要意义。本研究选用感病品种翠碧一号(CB-1)和抗青枯病突变体153-K为亲本,构建了F2群体,利用“主基因+多基因”混合遗传模型分析方法,研究其在安徽、福建两个病圃环境下的遗传效应,并对153-K青枯病抗性与农艺性状进行相关分析。结果表明,153-K在安徽病圃中的最优遗传模型为MX2-EEAD-AD,即2对等显性主基因+加性-显性多基因模型;153-K在福建病圃中最优遗传模型为MX2-AD-AD,即2对加性-显性-上位性主基因+加性-显性多基因模型。相关分析结果表明,在安徽、福建两个病圃环境下,青枯病抗性与株高呈显著负相关;而与叶片数、节距、茎围相关性均不显著。
关键词:烟草;青枯病;抗性突变体;遗传分析;相关分析
烟草青枯病(Tobacco wilt disease)是由青枯雷尔氏菌(Ralstonia solanacearum)引起的土传细菌性维管朿病害,该病害易感染、传播快、毁灭性强。烟草青枯病一般在烟草生长后期发生,发病后烟叶产量显著降低,烤后烟叶青杂烟、光滑烟较多,化学成分不协调,严重影响烟草的品质,可用性明显下降。截至目前,青枯病已成为热带、亚热带地区烟田的主要病害。在我国,以云南、福建、广东四川、贵州等南方烟区发病较为严重,某些年份甚至造成毁灭性损失,如:云南文山、临沧和保山等地。近年来,青枯病正在逐渐向北方烟区蔓延在山东、河南、辽宁等烟区均已报道。
青枯病的防治主要有农业、化学和生物防治等措施,但均不能有效预防青枯病的发生与蔓延,且各种防治措施都存在不足及局限性,比如在生产中大量使用农药对环境造成严重破坏等。因此明确烟草青枯病抗性的遗传规律,选育抗青枯病品种,是目前青枯病防治最基本、最有效、最经济的防治措施。
选育抗病品种,首先要有良好的抗源并分析其遗传规律。目前青枯病抗源来源十分狭窄,国内烟草青枯病的抗源主要是从普通烟草品种T448A选育而来,由其育成的DB101及其衍生的Coker139NC95和Coker319是抗青枯病育种的主体亲本。
本研究前期通过EMS技术诱变翠碧一号(CB-1)获得抗青枯病突变体材料153-K,将该突变体在温室和田间病圃进行青枯病鉴定,153-K均表现为高抗。因此,本研究利用153-K构建CB-1x153-K组合,并通过P1、P2、FI和F2多世代联合分析,探究153-K抗性基因的遗传规律,以期为培育优质的抗青枯病新品种提供理论基础,提高育种效率。
1材料与方法
1.1试验材料
试验材料:抗青枯病突变体153-K由中国农业科学院烟草研究所生物技术研究中心利用EMS诱变CB-1获得。以CB-1为父本,突变体153-K为母本,杂交获得F代,F1自交获得F2群体。
1.2试验设计
试验于2020年在安徽省宣城市寒亭镇和福建省福州市宦溪镇两个病圃进行。田间种植行距1.2m,株距0.5m。P,P2,F随机区组设计,重复3次,每重复10株。安徽F2群体调查205株,福建F2群体调查165株。
1.3农艺性状调查
按照烟草病虫害分级及行业标准规定的调查方法(YCT39-1996),以株为单位调查发病情况,并计算病情指数。
参照标准YC/T142-2010《烟草农艺性状调查测量方法》和文献[11]中所列方法对F2群体进行主要农艺性状调査,调查性状为株高、茎围、节距、叶片数。
1.4数据分析
采用Microsoft Excel和SPSS20.0软件进行数据处理、统计分析与相关分析。采用卡方检验和植物数量性状“主基因+多基因”混合遗传模型分析方法进行遗传分析。
2结果
2.1153-K抗病性遗传规律分析
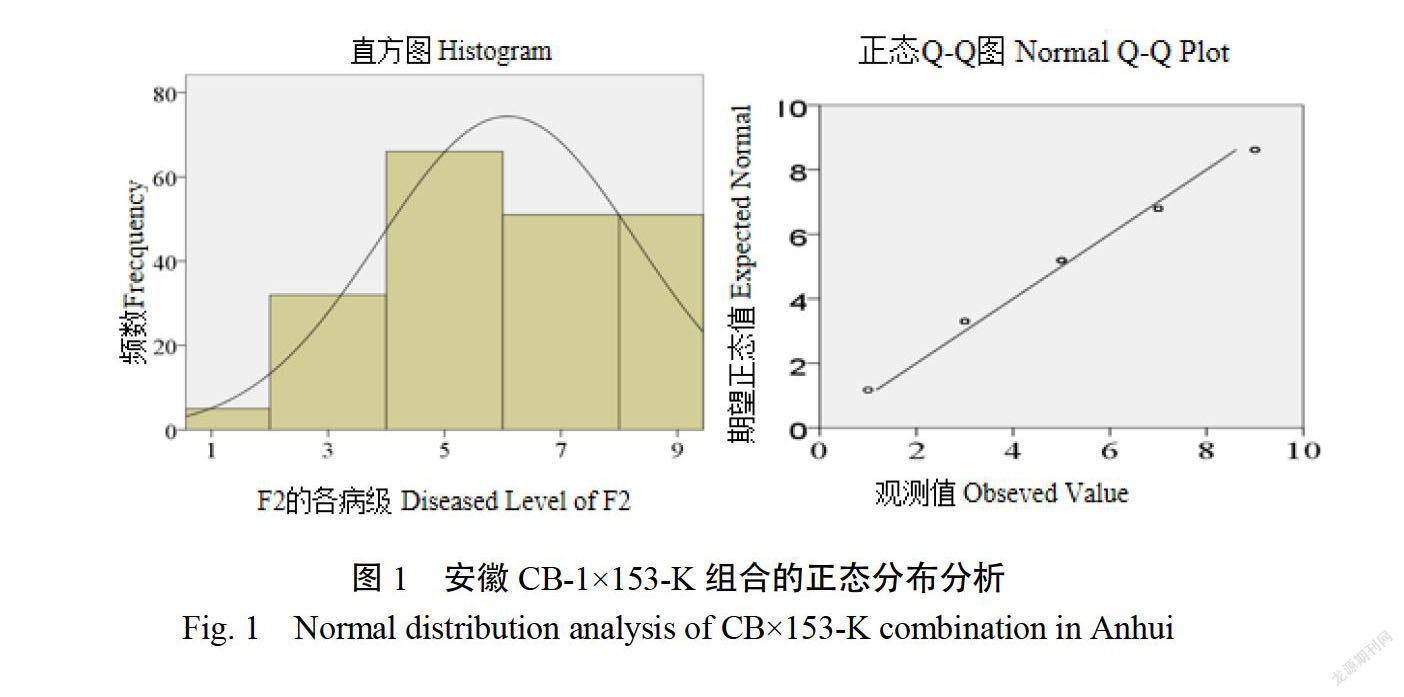
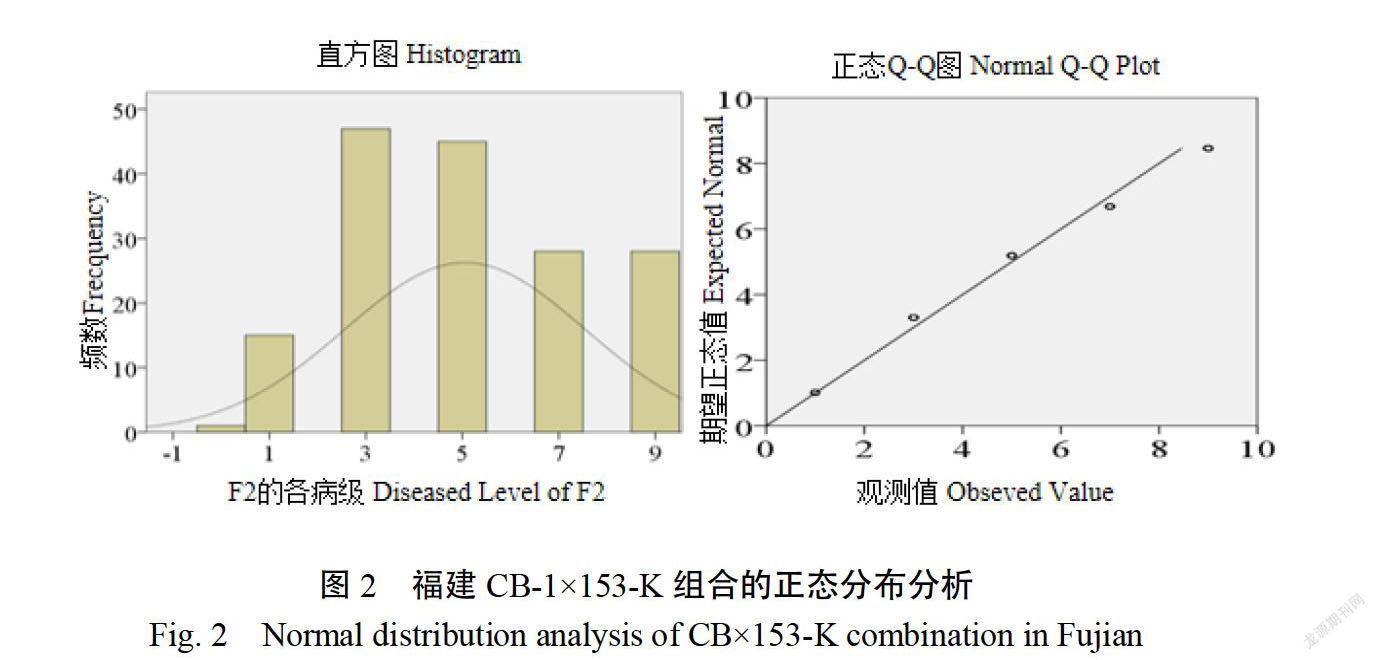
2.1.1病圃青枯病发病情况安徽P(CB-1)的病情指数为8333,P2(153-K)的病情指数为222;福建P1(CB-1)的病情指数为84.79,P2(153-K)的病情指数为31.58。在安徽、福建两个病圃环境下P(CB-1)对青枯病均表现为高感,突变体P2(153-K)对青枯病均表现为高抗。安徽F(CB-1x153-K)病情指数为90.65。福建F(CB-1×153-K)的病情指数为90.64,且没有完全抗青枯病植株的存在,这表明青枯病抗性为隐性遗传。通过SPSS20.0软件对F2代各病级株数进行正态分布分析(卡方检验),发现直方图中频率分布有明显的波峰,且偏度约等于0,说明F2代各病级株数呈正态分布;Q-Q图检验看出各病级的频数基本分布在直线附近,进一步说明F2代各病级株数服从正态分布(图1、2)。该结果表明两个环境下的F2代均存在一定性状分離,可以进行遗传规律的分析。
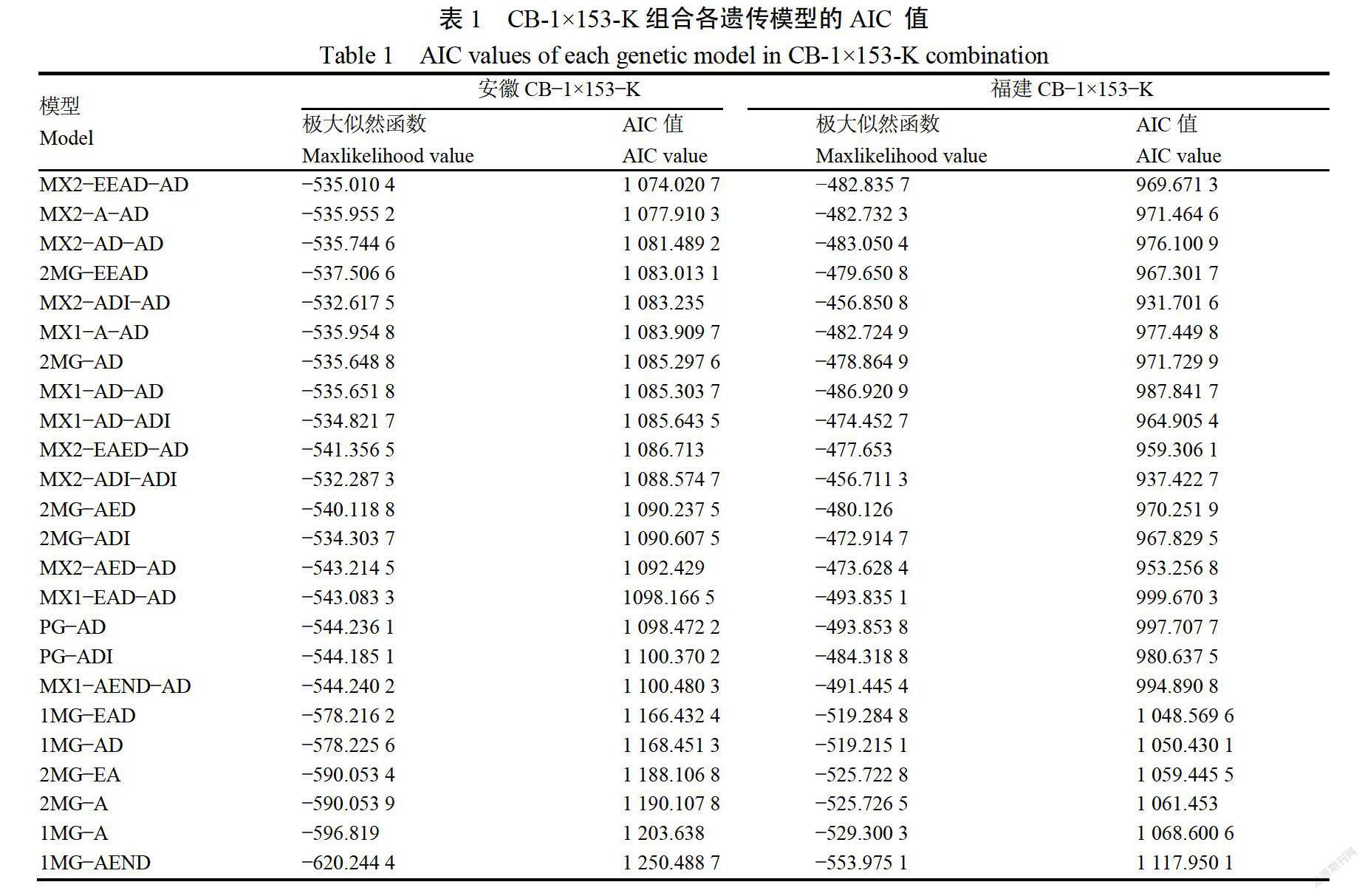
2.1.2主基因+多基因混合模型遗传规律分析利用P1、P2、F1和F2四个世代,对安徽、福建两个环境下的烟草抗青枯病突变体153-K的抗青枯病数量性状进行“主基因+多基因”混合遗传分析,得到两个环境下的24种模型的AIC值、极大似然值(表1),根据AIC值最小原则选择最优模型。在安徽CB-1153-K组合中,AIC值较低的有MX2-EEAD-AD、MX2-A-AD、MX2-AD-AD和2MG-EEAD;在福建CB-1x153-K组合中,AIC值较小的模型有MX2-ADI-AD和MX2-AD-ADI,将以上模型作为两个环境下的备选模型。
利用、2、る、n、Dm对安徽、福建两个环境下的备选模型进行适合性检验,统计量达到显著水平个数最少及AIC值最小的模型为最优模型。统计结果显示(表2),安徽CB-1×153-K组合中,MX2-EEAD-AD、MX2-A-AD、MX2-AD-AD和2MG-EAD分别有5、7、7、7个统计量与该模型的差异达到显著水平(p《0.05),根据AIC值最小且统计量差异达到显著水平最少原则,确定MX2-EEAD-AD模型为安徽CB-1×153-K组合的最优遗传模型,即2对等显性主基因模型;同理,在福建病圃CB-1×153-K组合中(表3),MX2-ADI-AD和MX2-ADI-ADI分别有8、8个统计量与该模型的差异达到显著水平(p《0.05),根据AIC值最小原则确定MX2-ADI-AD为福建CB-1×153-K组合的最优遗传模型,即2对加性-显性-上位性主基因模型。
2.13遗传参数估计对安徽、福建两个环境下最适模型的遗传参数进行估计。结果表明(表4),在安徽,群体平均数(m)为3.9687,第1对基因的加性效应(da)为1.6121,显性效应为0。第2对基因的加性效应和显性效应均为0,说明两对基因只存在加性效应。此外,ja=ja==0,说明两对主基因不存在相互作用。多基因的加性效应(の)为0.409,显性效应()为1.1795,以显性效应为主主基因的遗传率为71.81%。
在福建,CB-1153-K中(表4),群体平均数m)为3.4263,第1对基因的加性效应(da)为2.4959,显性效应(ha)为-15203,則d》ba且.显性度(d)小于1,说明第1对基因主要表现为加性作用。在第2对基因中,加性效应值(b)为0.4969,显性效应值(h)为-1.6512,则第2对基因主要表现为显性效应。对两对基因相互作用的参数值进行分析,加性×加性互作效应()为0.5268,加性显性互作效应(ia)为1.6213,显性x加性互作效应(ia)为2.4962,显性显性互作效应()为1.6746,上位性效应均为正向,其中ia》トia》i,表明上位性效应中以显性ⅹ加性互作和显性ⅹ显性互作效应较大,说明其促进了抗病性提高。主基因的遗传率为94.74%,表明抗性主要受主基因作用,可在后代中稳定遗传。
2.2153-K抗病性与农艺性状的相关性分析
2.2.1农艺性状对2020年安徽和福建田间农艺性状进行调查分析。从图3可以看出,CB-1株高显著高于153-K(p0.05)(图3A),叶片数与153-K接近,不存在显著性差异(图3B)(p》0.05),茎围显著粗于153-K(图3C)(《0.05);CB-1节距在安徽显著高于153-K,而两个品种在福建没有显著性差异(图3D)(p》0.05)。因此,突变体153-K与CB-1相比,153-K的单株表型为整体较矮小,而叶片数无显著性变化。
2.2.2F2群体的病情级别与农艺性状的相关分析对F2的病情级别(病级)与田间主要农艺性状进行相关分析,结果表明(表5),在安徽,病级与株高的相关系数是-0.475(p《0.01),为极显著负相关,与叶片数、节距、茎围为不显著负相关。在福建,病级与株高的相关系数是-0.221(p《0.05),为显著负相关,病级与叶片数、节距为不显著负相关,与茎围为不显著正相关。
3讨论
抗源材料是烟草抗青病育种的基础,T448A是国内外少数抗青枯病的种质资源之一。1945年美国首个育成抗青枯病品种Oxford26,后来相继培育出了DB101、Coker139。用Coker139育成的NC95、Coker319和G-28是20世纪70年代美国主要抗青枯病品种和主体亲本。而我国青病抗病育种研究起步较晚,开始于20世纪90年代。我国新审定的抗青枯病新品种有云烟、云烟2031、云烟87、岩烟97等。目前国内大部分青枯病抗性来源难以追踪,抗性来源明确的亲本也不属于常见的抗源。
改进传统的育种手段和方法,对快速培育烟草抗青枯病优良品种非常重要。EMS诱变技术是人为获得某些可遗传新材料的常用方法。中国农业科学院烟草研究所采用EMS处理“翠碧一号”种子,经过M2、M3代抗病性鉴定,获得“翠碧一号”抗青枯病突变体22个。突变体153-K就是通过EMS诱变技术获得的高抗青枯病的新抗源,为烟草抗青枯病育种及抗病机理的研究提供了材料。
烟草青枯病抗性是受遗传和环境两个相互作用因素的影响。前人研究表明,不同烟草种质资源对青枯病的抗性遗传存在差异。杨友才等对烟草抗性资源448A的抗性遗传机理进行研究,结果表明其抗性受显性单基因控制。QIAN等叫研究认为,岩烟97的青枯病抗性在田间表现为主基因控制。耿锐梅等对岩烟97的青枯病抗性进行分析,结果表明其抗性受2对加性、显性、上位性主基因以及加性、显性、上位性多基因控制。邹文莉等对烤烟品种翠碧一号通过EMS诱变而来的突变体117-K、486-K的抗性遗传规律进行分析,发现其抗性遗传效应存在一些差异,突变体117-K的抗性受2对主基因控制,具有加性效应。突变体486-K的抗性受2对主基因控制,具有加性-显性-上位性效应。
本研究对安徽、福建环境中突变体153-KxCB-1组合P1、P2、F1和F2四个世代采用“主基因+多基因”混合遗传模型进行联合分析,在安徽,153-K表现为2对主基因控制,具有等显性效应。在福建,受2对加性-显性上位主基因控制,与邹文莉等24对突变体486-K进行的青枯病抗性遗传规律研究结果一致。本研究在安徽、福建两个环境下,其抗性遗传效应存在一些差异,可能是因为:(1)不同环境下病情的分化差异很大。青枯病多发生在高温高湿条件下,土壤、气候等生态条件也会影响青枯病的发生。因此,同一品种在同一年份不同试验点的发病情况可能也有差异。(2)烟草青枯菌菌系分化比较复杂,因寄主范围、地理分布、致病性以及生理特性的多样性和复杂性被公认为多变的复合种青枯菌还具有种下遗传多样性。福建、安徽两个试验点的青枯菌都是青枯菌生理小种1号,以演化型进行划分时都是演化型1,但其演化型下具有序列变种差异性。福建省拥有13、14、15、17、34和44六个序列变种,安徽青枯菌为序列变种15,安徽、福建两个试验点青枯菌在序列变种上具有差异性。不同试验点的青枯菌在致病性和某些性状上可能也存在差异。这可能是153-K遗传模型不同的原因。总之,虽然两个试验点遗传模型略有不同,但153-K青枯病抗性都是受两对主基因控制,且均具有显性效应。
4结论
本研究采用卡方检验和“主基因+多基因”混合遗传模型联合分析了烟草抗青枯病突变体153-K在安徽、福建两地试验点的遗传规律,分别为MX2-EEAD-AD和MX2-ADI-AD模型。在安徽试验点,突变体153-K的抗性受2对主基因控制,具有等显性效应。在福建试验点中,突变体153-K的抗性受2对主基因控制,具有加性-显性-上位性效应。对安徽和福建两个试验点CB-1×153-KF2群体的病级与农艺性状进行相关性分析,结果表明,F2病级与株高显著负相关。叶片数、节距、茎围对青枯病发病程度影响不显著。
参考文献
[1]陈勇,金晨钟烟草青枯病的发生与防治现代农业科技,2009(10):92-93
[2]朱贤朝,王彦亭,王智发.中国烟草病害M北京:中国农业出版社,2002:152-162
[3]陈瑞泰,朱贤朝,王智发,等.全国16个主产烟省(区)烟草侵染性病害调查报告.中国烟草科学,1997(4):1-7
[4]刘勇,秦西云,王敏,等.云南省烟草青枯病危害调査与病原菌分离[J].中国农学通报,2007,23(4):311-314
[5]刘勇,范江,李永平.烟草抗青枯病育种研究进展.中国烟草学报,2012,18(6):93-99
[6]何明兴,沈亮,邱恒良,等,烟草青枯病的发生及防治.现代农业科技,2019(11):11115
[7] LIY, FENG J, LIU H L, et al. Genetic diversity and pathogenicity of Ralstonia solanacearum causing tobacco bacterial wilt in China[J]Plant Disease,2016,100(7):1288-1296
[8] MA L, ZHANG H Y, ZHOU X K, et al. Biological control tobacco bacterial wilt and black shank and root colonization by bio-organi fertilizer containing bacterium Pseudomonas aeruginosa NXHG29 Applied Soil Ecology, 2018, 129(5): 136-134
[9] SMITH T E, CLAYTON E E Inheritance of resistance to bacterial wilt in tobacco [J]. Journal of Agricultural Research, 1948, 760)
[10]王元英,周健中美主要烟草品种亲源分析与烟草育种.中国烟草学报,1995,2(3):11-22
[11]刘国顺.烟草栽培学D[M].北京:中国农业出版社,2003
[12]王建康,盖镒.利用杂种F2世代鉴定数量性状主基因-多基因混合遗传模型并估计其遗传效应[J].遗传学报,1997(5):43240.
[13]章元明,盖镒.数量性状分离分析中分布参数估计的ECM算法作物学报,2000(6):6970
[14]章元明,盖镒,张孟臣.利用PIFIP2和F2或F(2:3)世代联合的数量性状分离分析.西南农业大学学报,200022(1)
[15]李永平,卢秀萍,王颖宽烤烟新品种云烟202的选育及特征特性.中国烟草科学,2005(4):16-18.
[16]焦芳婵,肖炳光,李永平,等烤烟新品种“云烟203”的选育及特征特性.西南农业学报,2010,23(3):625-628
[17]李永平,王颖宽,马文广,等.烤烟新品种云烟87的选育及特征特性中国烟草科学,2001(4):38-42
[18]刘贯山,孙玉合.烟草突变体[M]上海:上海科学技术出版社,2016:345-353
[19]刘贯山.烟草突变体创制、筛选与鉴.中国烟草科学,201334(2):13-14.
[20]王新.烟草抗青枯病突变体的鉴定与转录组分析[D].北京:中国农业科学院,2018.
[21]杨友才,周清明,朱列书.烟草青枯病抗性基因的遗传分析及RAPD标记.中国烟草学报,2006(2):38-42.
[22] QIAN Y L, WANG X S, WANG D Z. The detection of QTLS controlling bacterial wilt resistance in tobacco(Ntabacum L)[J]Euphytica,2013(192):259-266
[23]耿锐梅,程立锐,刘旦,等烟草青枯病抗性遗傳效应分析.中国烟草科学,2019,40(4):7-13
[24]邹文莉,牛文利,杨华应,等.烟草青枯病抗性突变体486-K和117-K的遗传分析中国烟草科学,2020,40(2):1-7
