西沙永兴岛抗风桐与海岸桐群落凋落叶分解及中型土壤动物的贡献
2021-01-16王雅雅童升洪韩晓鸽简曙光申卫军
王雅雅,饶 鑫,童升洪,韩晓鸽,王 俊,简曙光,任 海,申卫军,*
1 中国科学院华南植物园, 广州 510560
2 中国科学院大学, 北京 100049
植物凋落物分解是生态系统中养分循环和能量流动的关键过程。全球尺度上净初级生产力50%以上通过凋落物分解归还土壤[1]。在生物和非生物因素的作用下,凋落物分解后参与土壤有机质形成,并释放养分元素到土壤中为植物生长提供营养[2-3]。前人研究表明,凋落物分解主要受气候、凋落物质量和土壤生物群落的影响[4-5]。在全球尺度上,凋落物分解过程主要受气候因素影响[6];而在局域尺度上,则依赖于凋落物质量和土壤生物[7]。植物凋落物质量通常用初始C∶N、木质素﹕N和木质素﹕P等化学计量比来衡量,初始C∶N和木质素﹕N越低,凋落物质量越高,越有利于分解[8-9]。土壤动物是凋落物分解过程中被广泛研究的生物因子[10],其对凋落物分解的作用主要通过以下三种方式:(1)取食或破碎凋落物增加微生物的接触表面积;(2)通过粪便改变凋落物化学组分;(3)通过改变微生物群落结构、刺激微生物活性等而加快凋落物分解[11-13]。土壤动物根据体径可分为大型(>2 mm)、中型(0.2—2 mm)和小型(<0.2 mm)土壤动物[14];中型土壤动物因其分布广泛、类群丰富及食性广等特征而备受关注[15-16]。土壤中型土壤动物主要由弹尾目和螨类组成,通过取食真菌、细菌以及植物凋落物碎屑而参与凋落物分解过程[17-18]。
从全球尺度看,土壤动物对凋落物分解的作用主要表现为正效应,平均可以解释凋落物损失的27%[5]。土壤动物对凋落物分解的贡献也受气候因素和凋落物类型的影响[6,19],在不同生态系统中土壤动物对凋落物分解的影响有差异。在草原生态系统中,中型土壤动物对低质量凋落物分解的促进效果明显[20];而在旱地生态系统中,凋落物分解并未受到土壤动物去除的影响,主要是由非生物的光解发挥主导作用[21];Powers等[22]在热带雨林的研究结果表明中型土壤动物的排除使凋落物的分解速率降低了50%;另有研究者在巴西中部的原始热带稀树雨林中的研究也得出了类似的结论:土壤动物对凋落物分解的贡献率达13%—57%[23],雨季的贡献更大。
我国南海诸岛基本上都是珊瑚岛,其中西沙群岛自然岛屿最多,植被最具代表性,自然乔木植被主要是抗风桐(Pisoniagrandis)纯林和海岸桐(Guettardaspeciosa)纯林。近年来,由于受到人类活动和全球变化影响而出现了退化现象,亟需进行植被恢复[24]。在植被养护管理方面以及重新造林过程中,维持或恢复当地原生植物群落组成至关重要[25]。由于土壤动物尤其是中型土壤动物能促进凋落物分解、加速碳氮周转和调节植被组成[26],它们在植被恢复过程中土壤肥力改善方面能发挥重要作用。恢复或新建的珊瑚岛礁植被生态系统中,土壤动物发挥作用的大小,也是评价恢复成效的一个重要指标。但是,目前关于我国南海珊瑚岛生态系统凋落物分解及土壤动物在分解过程中的作用还没有开展研究。
另外,在南海珊瑚岛一些新建的先锋群落中,观察到很多地表的凋落叶虽然叶肉组织已经剥落,但叶脉网络仍然完整保存,这可能是凋落叶分解速率慢的一种表现;猜想可能是缺少了土壤动物的碎化或微生物的作用,但缺少相关的实验验证。本研究采用凋落物袋法,探究南海西沙永兴岛优势树种抗风桐和海岸桐凋落叶分解过程中中型土壤动物的作用,为深入理解热带珊瑚岛生态系统的养分循环过程,土壤肥力形成过程和研发有效的土壤改良措施提供科学依据。
1 材料和方法
1.1 研究区概况
本研究的研究样地位于西沙永兴岛,是海南三沙市政府所在地。永兴岛位于北纬16°50′,东经112°20′,属于中国西沙群岛东部的宜居群岛,是南海诸岛中面积最大的自然岛屿(约2.10 km2)。该区域属热带海洋性季风气候,年平均降水量1500 mm,多集中在5—9月,干湿季差异明显。年平均气温为26.3℃,年太阳辐射量大,日照时数长,终年高温,蒸发量大。土壤基质为珊瑚和贝壳类残体在鸟粪的长期作用下形成的磷质石灰土和滨海盐土。岛上主要植物群落为抗风桐、海岸桐和草海桐(ScaevolasericeaVahl)等形成的单优群落[27-29]。
1.2 样地布设
本研究选择抗风桐纯林和海岸桐纯林作为研究样地。在每个样地中随机布置3个5 m×5 m的重复样方,同时分别以样地中的抗风桐和海岸桐凋落叶为研究对象,选用孔径分别为0.1 mm和2 mm,规格为15 cm ×20 cm的凋落物袋。0.1 mm孔径的凋落物袋基本排除中型土壤动物,但允许小型土壤动物进入(主要为线虫、原生动物等湿生类动物),视为无中型土壤动物对照;2 mm孔径凋落物袋内允许中型和小型土壤动物进入,但排除大型土壤动物,视为有中型土壤动物处理。两种孔径凋落物袋质量损失之差即为中型土壤动物的贡献[18]。在2018年7月开始布置凋落物分解实验,每个凋落物袋内装入风干后的凋落叶10 g。在每个样方的土壤表面放置两种孔径的凋落物网袋各20袋,共240袋,分别于2018年10月、2019年1月、4月、8月和10月进行凋落物袋回收,每次每个样方回收2—3袋。其中,一袋用于失重率测定,45℃烘箱中烘48 h至恒重,用毛刷轻轻去除凋落叶表面的杂质和砂砾后称重,烘干的凋落叶样品用研磨仪(JXFSTPRP- 48, 中国)粉碎后过60目筛,留待进行元素测定。其中,凋落叶TC、TN和TP含量分别采用重铬酸钾容量法-外加热法、奈氏比色法和钼锑抗比色法进行测定;凋落叶木质素、纤维素和半纤维素含量采用中性和酸性纤维洗涤法(Van Soest法)测定。一袋用于中型土壤动物分离,利用Tullgren法提取中型土壤动物,在Olympus体视显微镜(SMZ140-N2LED)下镜检,并鉴定、分类[14]。
在进行凋落物袋布设的同时,在样地中安装土壤温湿度自动记录仪(MC301N, 美国),实时监测表层土壤的温度和湿度。此外,分别在布置凋落物袋时,在每个样方内用直径35 cm的土钻各取5钻0—5 cm和5—10 cm层土壤,充分混合,风干后过筛,进行基本理化性质分析,包括pH、含水量、土壤总有机碳(Total organic carbon, TOC)、土壤TN及土壤TP等的分析。土壤样品pH测定用电位法(水土比1∶2.5);含水量测定用烘干法(105℃, 24 h);土壤TOC、TN和TP的测定方法与上述植物凋落叶的分析方法相同。
1.3 数据处理与分析
凋落叶分解快慢用失重率、残留率和分解常数来度量,计算公式如下:
凋落叶失重率:
Lt=(W0-Wt)/W0×100%
(1)
凋落叶元素残留率:
Rt=(Ct×Wt)/(C0×W0)×100%
(2)
凋落物分解常数(Olson衰减指数模型[30]):y=Wt/W0=a·e-kt
(3)
式中,W0为初始凋落叶干重(g),Wt为某次取样时的凋落叶干重(g);C0为初始凋落叶养分元素的含量(%),Ct为取样时养分元素的含量(%),Wt/W0表示质量残留率(%),a为分解系数,k为分解常数,t为分解时间(月)。
土壤动物群落组成特征以及对凋落叶分解的贡献率(Cf)用以下公式计算:
土壤动物对凋落叶分解的贡献率:
Cf=(L2-L0.1)/L0.1×100%
(4)
(5)
Pieluo均匀性指数:
Jwx=H′/lnS
(6)
Margalef丰富度指数:
D=(S-1)/lnN
(7)
公式(4)—(7)中L2和L0.1分别表示2 mm和0.1 mm孔径凋落物袋内凋落叶失重率。S为中型土壤动物总类群数,Pi为第i个类群的个体数占总个体数的比例,N为类群总个体数。
采用独立样本t检验分析不同孔径凋落物袋处理间的差异,单因素方差分析(one-way ANOVA)检测分解时间对凋落叶分解的影响,Pearson相关性分析探究土壤动物对凋落叶分解的贡献率与土壤动物群落结构变化的相关性。统计分析采用SPSS21.0软件,作图用Origin9.0和R.3.4.4软件。
2 结果与分析
2.1 土壤理化性质和凋落物质量
永兴岛抗风桐和海岸桐群落土壤含水量、土壤温度与湿度,以及pH、TOC、TN和TP含量均没有显著差异。这些土壤理化性质在两个取样土层(0—5 cm和5—10 cm)之间也没有显著差异(P>0.05; 表1)。这些结果表明两个植物群落凋落叶分解的小气候环境相似。

表1 抗风桐和海岸桐纯林土壤理化性质(均值±标准误)Table 1 Soil physical and chemical properties in the P. grandis and G. speciosa forests (mean±SE)
除半纤维素含量外,两个树种凋落叶的初始化学质量存在一定差异。抗风桐凋落叶的TC、TP、木质素和纤维素含量显著低于海岸桐,而其TN含量显著高于海岸桐凋落叶(P<0.05; 表2)。抗风桐凋落叶的C∶N和木质素﹕N均显著低于海岸桐凋落叶(P<0.05; 表2)。因此,抗风桐凋落叶初始质量显著高于海岸桐。
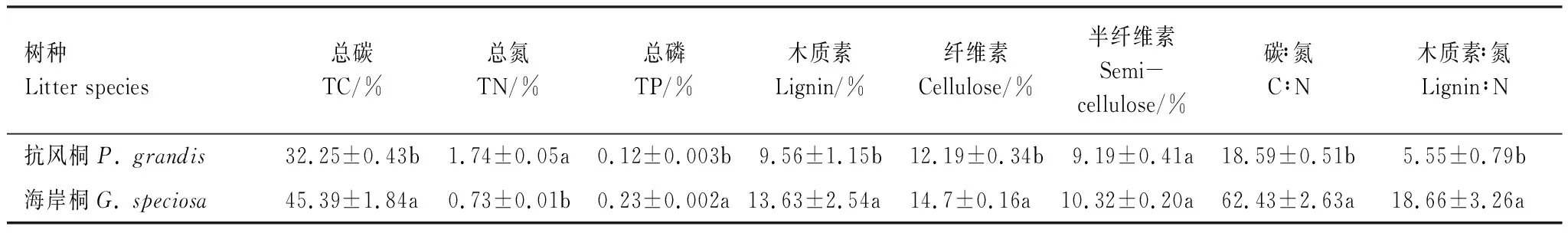
表2 抗风桐和海岸桐凋落叶初始化学质量(均值±标准误)Table 2 Initial chemical properties of P. grandis and G. speciosa leaf litter (mean±SE)
2.2 质量损失和养分释放
在两种凋落叶分解15个月期间,凋落叶失重率受凋落物袋孔径和分解时间的影响。本研究中,两种凋落叶分解过程大体可分为两个阶段,分解前6个月失重较快,6个月后分解速率变慢。分解前6个月,抗风桐0.1 mm凋落物袋内凋落物失重率(43.5%)显著低于2 mm凋落物袋内凋落叶失重率(60.3%)(P<0.05; 表3);海岸桐0.1 mm和2 mm孔径凋落物袋内凋落叶失重率分别为33.3%和34.3%,两者之间差异不显著(P>0.05; 表3)。分解15个月后,抗风桐0.1 mm凋落物袋内凋落叶失重率(56.8%)显著低于2 mm凋落物袋内凋落叶失重率(77.6%)(P<0.05; 表3);海岸桐两种孔径凋落物袋内凋落叶失重率分别为51.0%和51.7%,两者之间无显著差异(P>0.05; 表3)。
中型土壤动物对抗风桐和海岸桐凋落叶分解的贡献率随分解时间变化。在分解0—6个月内,对抗风桐和海岸桐的平均贡献率分别为12.3%和4.8%(P<0.05; 表3);分解6—15个月内,平均贡献率分别为33.0%和12.3%(P<0.05; 表3)。这些结果显示,中型土壤动物对抗风桐凋落叶失重率在分解早期和晚期均有显著影响,而对海岸桐凋落叶失重率仅在分解后期有显著影响。
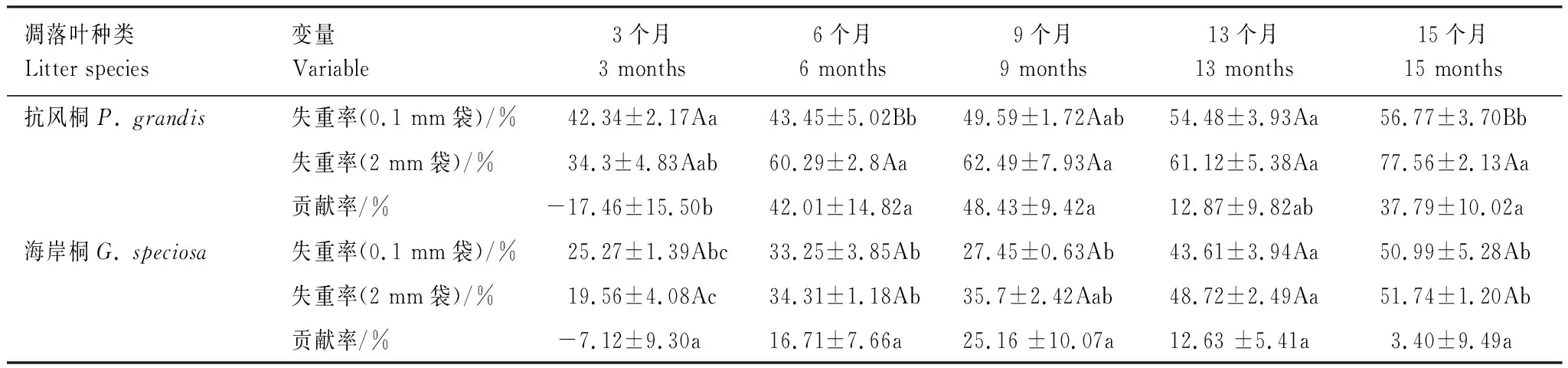
表3 抗风桐和海岸桐凋落叶分解过程中失重率及中型土壤动物贡献率的变化(均值±标准误)Table 3 Mass loss and mesofauna contribution ratios (mean±SE) to the decomposition of P. grandis and G. speciosa litters
根据经典Olson分解模型拟合凋落物质量残留率随分解时间动态变化及计算分解常数。抗风桐凋落叶在0.1 mm和2 mm孔径凋落物袋内的分解常数分别为0.05和0.08,也即分解周期分别为20个月和12.5个月,两种凋落袋处理间差异显著(P<0.01; 图1)。海岸桐分解常数在两种孔径凋落物袋处理下分别为0.04和0.05,也即分解周期分别为25个月和20个月,凋落物袋处理间差异不显著(P>0.01; 图1)。因此,两种孔径凋落物袋处理显著影响抗风桐分解常数和分解周期,而对海岸桐没有显著影响。

图1 两种孔径凋落物袋处理下抗风桐和海岸桐凋落叶分解过程中质量残留率变化Fig.1 Mass remaining ratios of the two litter species (P. grandis and G. speciosa) in the two mesh-size litterbags实线是0.1 mm孔径凋落物袋内质量残留率随分解时间的拟合曲线,虚线是2 mm孔径凋落物袋内质量残留率随分解时间的拟合曲线,R2为决定系数,表示方程拟合度;t50%和t95%分别表示凋落物分解50%和95%时所需要的时间(月)
抗风桐和海岸桐凋落叶在分解过程中表现出相似的养分释放变化趋势。随着分解过程的持续,两种凋落叶的TC和纤维素残留率呈现出降低趋势;TN残留率则呈现出升高趋势;TP、木质素、半纤维素、C∶N及木质素﹕N的残留率先上升后下降,其中TP残留率变化在时间上差异不显著(P>0.05; 图2)。
独立样本t检验表明,抗风桐和海岸桐的养分元素残留率在不同分解时期受凋落物袋孔径影响(P<0.05; 图2)。与2 mm凋落物袋相比,0.1 mm凋落物袋内抗风桐凋落叶TC、纤维素、木质素和半纤维素的释放在分解后期显著减少(P<0.05; 图2),残留率分别下降了57.1%、53.8%、55.6%和22.5%,促进了抗风桐TN的富集,残留率提高了64.5%;而对于海岸桐,0.1 mm凋落物袋处理抑制了其凋落叶分解过程中纤维素和半纤维素的释放,但分解15个月后养分残留率在两种孔径凋落物袋处理间差异不显著(P>0.05; 图2)。因此,抗风桐和海岸桐凋落叶分解过程中元素释放变化趋势相似,但在不同分解时期养分元素残留率受凋落物孔径影响,且其对抗风桐养分释放影响更大。

图2 两种孔径凋落物袋处理下抗风桐和海岸桐的养分元素残留率随分解时间的变化Fig.2 Dynamics in nutrient mass remaining for P. grandis and G. speciosa in two mesh-size litterbags不同小写字母表示不同分解时间凋落叶失重率在0.05水平上差异显著,*表示同一分解时间不同孔径凋落物袋处理间差异显著(*P<0.05, **P<0.01)
2.3 中型土壤动物群落组成随分解时间的变化
在凋落叶分解过程中,抗风桐和海岸桐凋落物袋内共计捕获中型土壤动物462只,其中优势类群为螨类,常见种为腹足纲、虫齿目和弹尾目等。按食性分为杂食、食菌、食植、食腐和食肉五大功能类群,其中杂食性占主要优势。
中型土壤动物群落结构存在季节差异,在分解13个月后中型土壤动物类群数和总个体数达到峰值,主要是真螨目和寄螨目动物(表4);多样性和丰富度也达到最高。但不同分解阶段抗风桐和海岸桐凋落物袋内中型土壤动物的多样性指数、均匀度指数和丰富度指数变化趋势相似,差异不显著(图3)。

表4 凋落叶分解过程中中型土壤动物种类和数量Table 4 Taxa and number of soil mesofauna individuals observed at different sampling times

图3 抗风桐和海岸桐凋落叶分解过程中中型土壤动物群落组成的变化Fig.3 Changes in soil mesofauna community composition with decomposition time for P. grandis and G. speciosa litters 不同小写字母表示不同分解阶段各指标在P<0.05水平上差异显著;D为Margalef丰富度指数; H′为Shannon-Wiener多样性指数; Jwx为Pieluo均匀性指数
用两种凋落叶5次采样时间的数据进行Pearson相关分析,结果显示中型土壤动物个体数和类群数与凋落物质量、中型土壤动物贡献率以及土壤湿度的相关性均不显著。中型土壤动物个体数和类群数仅与土壤温度呈显著正相关性(P<0.05; 图5)。

图4 中型土壤动物个体数、类群数与凋落叶化学性质、土壤动物贡献率及土壤温湿度的相关性Fig.4 Pearson correlations of mesofauna abundance and richness with litter chemistry, mesofauna contribution and soil climate蓝色小方框代表正相关系数,橙色小方框代表负相关系数,颜色越深相关性越强
3 讨论
本研究结果表明,中型土壤动物的排除减缓了两种凋落叶分解过程中的质量损失,影响了养分释放过程,中型土壤动物的贡献因凋落叶类型和分解时间而异,这与绝大多数前人研究结果一致[5,19,31]。龙健等[32]在茂兰喀斯特森林的研究结果认为土壤动物对初始凋落物质量高的凋落物分解贡献更大;Fujii等[33]对不同海拔梯度上森林凋落物分解的研究结果也显示中型土壤动物对高质量凋落物分解的贡献率更大。研究结果也显示中型土壤动物的排除对抗风桐凋落物叶分解的影响更大;抗风桐的C∶N和木质素﹕N显著低于海岸桐,初始凋落物质量较高,中型土壤动物可能更加偏好取食这种质量较高的凋落叶,从而更有利于其分解。然而,在青藏高寒草甸凋落物分解的研究显示,土壤动物对C∶N高的凋落物分解促进作用更大[34],这可能是低温干旱的环境下,土壤动物对C的利用需求更高导致的。有一些学者在温带森林和热带雨林的研究同样支持中型土壤动物对质量低的凋落物分解作用更显著[25,35],可能的原因是中型土壤动物通过捕食作用调节真菌分解者群落结构,使真菌的多样性更高,更有利于低质量凋落物分解;这说明中型土壤动物对凋落物分解的作用受凋落物质量的影响,但更依赖于气候因素调控。综上,本研究中中型土壤动物对两种植物凋落叶分解的作用差异,一方面可用初始凋落物质量来解释,另一方面可能还受到环境因子的限制。
根据当前的研究结果,中型土壤动物显著影响抗风桐和海岸桐养分释放过程,且主要在分解后期较明显;这与王振海[36]对长白山针叶林凋落物分解的研究结果类似。土壤动物对凋落物分解初期养分释放的促进作用不显著,可能与初期侵入凋落物袋内的土壤动物数量和种类较少有关。本研究结果显示,中型土壤动物显著促进了分解过程中抗风桐TC、TN、TP、纤维素、木质素和半纤维素的释放,显著促进了海岸桐TN、纤维素、半纤维素的释放并抑制了其TP的释放,且显著提高了抗风桐和海岸桐分解前期的C∶N和木质素﹕N。Lin等[30]在亚热带森林的研究结果也表明土壤动物促进凋落物TC和TN的释放;然而,在亚高山湿润森林的研究表明,分解前期土壤动物抑制凋落物C和N的释放,分解后期促进高质量凋落物C和N的释放[37]。这与本研究结果不一致,中型土壤动物在分解前期促进了抗风桐和海岸桐TN的释放及抗风桐TC的释放,可能是由于土壤动物刺激了微生物的活性,加速了微生物对C、N的降解。Qiu等[38]对长白山阔叶林的研究结果显示,中小型土壤动物抑制了P的释放;刘瑞龙等[39]在川西亚高山和高山森林中的研究中发现土壤动物促进高质量凋落物分解过程中P的释放,但抑制低质量凋落物P的释放,这说明土壤动物对凋落物P的释放的影响受初始凋落物质量影响。我们的研究结果与大多数研究一致[33,39-40],土壤动物促进凋落物分解过程中纤维素、木质素和半纤维素的降解。综上,中型土壤动物影响凋落物分解过程中养分元素的释放,对两种凋落物影响差异显著,这主要可能归因于初始凋落物质量的差异。
中型土壤动物群落结构的季节性变化可能主要受土壤温湿度变化的影响[6],反映在分解后期(2019年8月)土壤温湿度较高时,优势种螨类和弹尾目等数量增多,土壤动物多样性达到顶峰。一般认为,土壤动物群落结构变化是凋落物分解的重要调控因子,土壤动物丰富度和多样性高有利于凋落物分解[41]。而本研究结果显示中型土壤动物个体数和类群数的变化与分解贡献率在统计上相关性不显著。罗媛媛等[42]在亚热带常绿阔叶林的研究表明不同分解阶段各种中小型土壤动物类群优势度因其功能不同而异,如腐食性的弹尾目在分解后期比例较大;我们的研究结果也类似,分解后期真螨目和寄螨目数量较多。因此,中型土壤动物对凋落物分解的作用可能不是由多样性决定的,而是一些优势类群决定的。杨志敏等[43]在内蒙古典型草原的研究结果表明在光辐射强的地区土壤动物对凋落物作用较弱,而光解起主要作用。南海珊瑚岛光照也比较强烈,光解作用在凋落物分解中的贡献也值得进一步研究。也有研究者认为排除中型土壤动物对凋落物分解的影响可能是由凋落物孔径大小不同引起的凋落物袋内微气候的差异导致的[44-45],我们的研究结果中也可能有不同孔径凋落物袋的影响效应。土壤生物在南海珊瑚岛凋落物分解中的作用还需要进一步的实验来证实,如利用13C同位素标记的凋落叶进行分解实验,再检测土壤动物或微生物生物量的13C,可证明特定土壤生物对凋落叶分解的直接贡献。
4 结论
抗风桐凋落叶具有更快的分解速率与更短的周转期。从加速元素周转的角度来看,选用抗风桐作为先锋树种进行退化珊瑚岛植被恢复更为有效。中型土壤动物显著影响了西沙群岛抗风桐和海岸桐凋落叶分解过程中的质量损失和养分释放;影响的程度因凋落物类型和分解时间而异。中型土壤动物的存在能促进凋落叶分解,对质量较高的抗风桐凋落叶分解贡献更大。中型土壤动物对抗风桐凋落叶分解在整个分解期间都有显著贡献,但对海岸桐凋落叶分解的贡献仅在后期显著。中型土壤动物群落总个体数与类群数主要与土壤温度显著相关,它们与凋落叶分解贡献率之间的统计相关性不显著。中型土壤动物群落对凋落叶分解的影响可能主要是一些优势类群贡献的,可能光解作用和不同孔径凋落物袋或它们的交互作用也有一定的影响效应。不同因素或机理在南海珊瑚岛生态系统凋落叶分解和元素循环中的作用仍需进一步的研究。
