青藏高原高寒灌丛2003—2016年生长季的不同月份CO2通量对气温日较差的响应
2021-01-16祝景彬贺慧丹李红琴张法伟李英年杨永胜张光茹王春雨罗方林
祝景彬, 贺慧丹, 李红琴, 张法伟, 李英年,5,*, 杨永胜,5, 张光茹,4, 王春雨,4, 罗方林,4
1 枣庄学院旅游与资源环境学院,枣庄 277160
2 中国科学院西北高原生物研究所,西宁 810001
3 洛阳师范学院生命科学学院,洛阳 471934
4 中国科学院大学,北京 100049
5 中国科学院高原生物适应与进化重点实验室,西宁 810001
由于化石燃料的燃烧和土地利用方式的改变,导致大气中CO2浓度显著增加,根据IPCC第五次评估报告,到2100年全球平均气温将升高0.3—4.8℃[1]。草地生态系统约占全球陆地总面积的三分之一,是地球碳循环的重要组成部分。与低海拔或低纬度地区相比,高海拔和高纬度地区的气温增长率更高[2]。高寒生态系统的碳平衡动态对气候变化极其敏感,并且气温是影响高寒生态系统碳循环的最关键因素[3]。因此,充分了解高寒生态系统碳循环的主要过程,对于有效预测全球碳循环对环境变化的响应具有重要意义[4]。过去大量的研究集中在环境因素(如温度、辐射、水和土壤营养)、生物因素等对高寒生态系统碳通量的影响[5-6]。然而,对于高寒生态系统碳通量与日较差(ADT)之间关系的研究相对较少。
过去的研究认为,ADT主要是通过较高的白天温度促进植被光合速率,以及较低的夜间温度降低生态系统呼吸来实现促进碳水化合物的积累[7]。但是,由于地理位置的差异,以及气候环境因素的不同,CO2通量对ADT的变化可能有不同的响应[8-9]。大量的研究表明,大气温度的不对称增温会对生态系统生产力产生不同的影响[10-11]。例如,在北美草原的研究表明,植被生物量与日最低气温之间存在显著的正相关关系,而与日最高气温和日平均气温没有显著的相关性[12]。但是,在菲律宾的研究发现,植被生物量与日最低气温显著负相关[13]。有研究表明,如果植物呼吸作用在夜间消耗更多的碳水化合物,将会刺激次日植物光合作用,使植被光合生产能力更强[14-15],这说明ADT的降低可能会导致光合作用的过度补偿,可能会减弱甚至抵消温室效应的影响,最终维持生态系统碳的稳定[16]。这些差异主要是由不同研究地点的环境因素和水热条件的差异造成的[8]。然而,由于对中国草地碳通量的研究起步较晚,这些研究主要集中在中国的低海拔地区[3, 6],ADT可能对低温高海拔的青藏高原地区碳通量产生不同的影响。
青藏高原对气候变化极其敏感,被称为全球的“预警区域”,在全球气候行为中发挥着至关重要的作用[17]。近50年来所观测到的地表气温的增温并非均匀,在不同的地理位置和不同的时间框架内,气温变化存在很大的差异[18]。这种变暖趋势中存在昼夜增温不对称,夜间温度的上升比白天温度的上升更快,即ADT将会减小[19]。然而,ADT的减小对青藏高原高寒生态系统碳通量的影响尚不清楚。所以,了解青藏高原生态系统的CO2动态及其对气候变化的响应,对于有效预测全球碳循环对环境变化的响应极其重要[6]。涡度协方差法(EC)是测量大气和各种生态系统之间碳收支的最有用的方法之一,采用EC法测定了青藏高原大气与高寒灌丛生态系统的CO2交换。本研究的总体目标是:1)量化高寒灌丛生态系统在生长季CO2通量与ADT的变异特征。2)明确在生长季的不同月份CO2通量对ADT的响应。
1 材料与方法
1.1 试验地概况
试验在青海海北高寒草甸生态系统国家野外科学观测研究站(海北站) 附近的高寒灌丛中进行。海北站地处青藏高原东北隅祁连山北支冷龙岭东段南麓坡地(37°29′—37°45′ N,101°12′—101°23′ E,海拔3200—3600 m)。该地区属高原大陆性气候,有明显的干冷(11月至次年4月)和暖湿(5月至10月)季节。根据1980年至2010年的气象资料,平均年气温约为-1.0℃,最温暖的月份在7月约为10℃,最冷的月份在1月约为-15℃。年平均降水量约580 mm,其中80%的降水集中在暖湿季节[20]。高寒灌丛生态系统的植被冠层由灌木和禾本科两层组成。植被以金露梅(Potentillafruticosa)为主要建群种,冠层高度约为30—60cm,灌木覆盖度约为60%—80%。草的次生层由矮嵩草(KobReiahumilis)、垂穗披碱草(Elymusnutans)、异针茅(Stipaaliena)、麻花艽(Gentianastraminea)等组成,在生长高峰期(7—8月),植被冠层高度约8—16 cm,植物覆盖度70%—80%。土壤是一种粘性壤土,草地在4月底或5月初开始变绿,地上生物量从5月到8月呈上升趋势,在7月下旬或8月达到最大值[21]。
1.2 研究材料及分析方法
1.2.1通量和相关监测
利用涡度相关技术对研究区CO2通量进行长期连续的观测(2003—2016年)。海北站通量塔安装在相当平坦的区域中心(坡度< 1%),植被相同(灌木覆盖率70%),覆盖面积约12km2。通量塔高为2.2 m,通量观测的主要仪器为开路式涡度相关观测系统和常规气象观测系统, 其中涡度相关系统包括三维超声风速仪(CSAT3, Campbell Scientific, Logan, USA) 、红外气体分析仪(LI- 7500, LI-COR Lincoln, USA) 和数据采集器(CR5000, Campbell Scientific, Logan, USA),主要用于观测植被与大气界面的CO2通量,数据采集频率为10 Hz,每30 min输出平均值。
常规气象观测系统主要用于连续气象要素采集,数据采集(Model CR23X, Campbell Scientific, Logan, USA)时段与通量数据相同,观测项目主要包括温度、湿度、辐射、风向、风速等。光合有效辐射(PPFD)传感器(L1190SB, LI-COR, Lincoln, USA)安装在高1.2 m的支架上。空气温度/湿度传感器(Model HMP45C, Vaisala, Helsinki, Finland)安装在防辐射罩内(Model 41002, RM Young Company, Michigan, USA)。土壤温度(Ts)主要是利用热电偶温度传感器(107-L, Campbell Scientific, Logan, USA)分别测定5、10、20、40和80 cm处的Ts。5、20和50 cm处的土壤含水量(SWC)利用时域反射计(Model CS615-L, Campbell Scientific, Logan, USA) 测定。
依据长期地面群落物候的观测,将返青期(5月1日—5月31日)和枯黄期(10月1日—10月31日)定义为植被生长季(6月1日—9月30日)与非生长季(11月1日—4月30日)的过渡期[6]。本研究中分析的是生长季(6至9月)的数据。日最高温度(MaxTa)与日最低温度(MinTa)之差为当天的日较差(ADT)。月GPP、Re、NEE分别为当月的日GPP、Re、NEE的累加值,生长季GPP、Re、NEE分别为当年整个生长季的日GPP、Re、NEE的累加值。月MaxTa、MinTa、ADT分别为当月的日MaxTa、MinTa、ADT的平均值,生长季MaxTa、MinTa、ADT分别为当年整个生长季的日MaxTa、MinTa、ADT的平均值。
1.2.2数据处理和插补
由于供电间断、探头结露或仪器异常等情况,涡度相关观测系统采集到的原始数据缺失或异常现象十分普遍[22]。通量数据的处理按照ChinaFLUX推荐的通量数据处理标准流程进行[23],在经过WPL校正后[24],以|NEE| < 1.0 mg CO2m-2s-1为阈值标准,再以10d数据为窗口,3δ (标准差)为标准进行数据的异常剔除。针对夜间 (PPFD < 10 μmol m-2s-1)湍流发展不充分,舍去摩擦速度u*<0.15 m/s所对应的通量数据。缺失的通量数据,一般通过建立环境因子与有效通量数据的非线性经验方程进行插补。本研究夜间通量数据(Reco,n)采用其与5 cm深处土壤温度(Ts)的Van′t Hoff函数(方程(1))进行插补,生长季白天的碳通量数据(NEE)则利用与PPFD的Michaelis-Menten函数(方程(2))进行插补[25-26]。
Reco,n=Reco,refeln(Q10)(Ts-10)/10
(1)
(2)
式中,Reco,ref为生态系统在10℃时的参考呼吸速率,Q10是生态系统呼吸熵(呼吸随温度升高10 ℃所增加的倍数),反映了生态系统呼吸对温度的敏感性。(2)式中,Reco,d是生态系统白天的呼吸速率,a和Pmax分别为生态系统表观光量子效率和潜在最大CO2同化速率(即PPFD趋向极大时生态系统净光合速率)。以上参数均为方程拟合值。
涡度相关系统无法直接测定生态系统总初级生产力(GPP)和生态系统呼吸(Re),需利用公式外推得到。涡度相关系统观测的CO2通量为NEE,将夜间呼吸函数外延至白天(Reco,d),即可得生态系统的总呼吸量(Re), 进而可得生态系统总初级生产力(GPP,方程(3)),日GPP由Re减去NEE得到,如式(3)所示,日Re为白天呼吸(Reco,d)和夜间呼吸(Reco,n)之和。负NEE和正NEE分别代表生态系统对CO2的吸收和释放[27]。MaxTa、MinTa、ADT和CO2通量的关系采用回归分析,对所测定的数据利用Excel和SPSS20.0软件对数据进行统计分析。
GPP=Re-NEE =Reco,d+Reco,n-NEE
(3)
2 结果与分析
2.1 高寒灌丛在生长季日较差的变化特征
图1可见,在2003—2016年的生长季中每日MaxTa和MinTa的均值呈现先升高后降低的单峰变化趋势。6、7、8、9月的日MaxTa和MinTa的平均值分别为(14.0±0.7)、(16.6±0.8)、(16.2±0.9)、(12.1±0.8)℃和(0.2±0.6)、(3.5±1.3)、(2.9±1.3)、(-0.3±1.2)℃。但是,日ADT的均值在生长季的波动较小,没有表现出明显的变化趋势(图1),说明在青藏高原ADT的变化与温度的变化并不同步。6、7、8、9月的日ADT平均值分别为(13.6±0.4)、(13.1±1.3)、(13.2±0.9)、(12.1±1.1)℃,说明9月的日ADT相对偏低。同时,分析表明2003—2016年整个生长季的日MaxTa、MinTa和ADT的均值并没有表现出明显的年际变化趋势(P>0.05)。
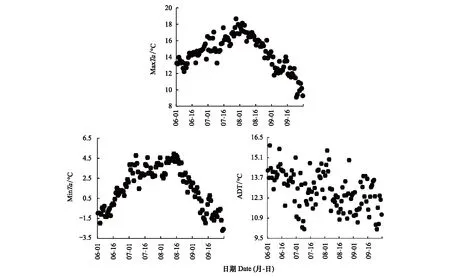
图1 高寒灌丛在2003—2016年生长季(6—9月)平均日最高气温(MaxTa,℃)、日最低气温(MinTa,℃)和日较差(ADT,℃)Fig.1 The average of diurnal maximum temperature (MaxTa,℃), the diurnal minimum temperature (MinTa,℃), and the amplitude of diurnal temperature (ADT,℃) of alpine shrubland in the growing season (June—September) from 2003 to 2016
表1可见,在生长季各月的日ADT与MaxTa呈极显著正相关(P<0.001),日ADT与MinTa呈极显著负相关(P<0.001),通过比较“r2”的值发现只在6月MaxTa对ADT的主导作用相对较强,在其他月份MinTa对ADT的变化有相对更强的控制。此外,线性回归也表明,整个生长季的日MinTa(r2=0.39,P<0.001)比MaxTa(r2=0.33,P<0.001)对日ADT变化起相对更强的控制作用。
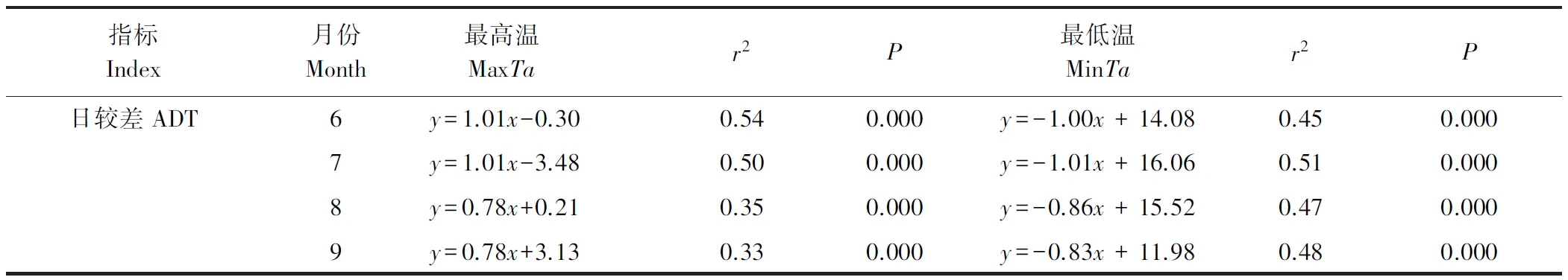
表1 生长季每个月的日MaxTa、MinTa与ADT的线性回归Table 1 Linear regression between daily MaxTa, MinTa and ADT of each month in growing season
2.2 高寒灌丛在生长季CO2通量的变化特征
在2003—2016年的生长季的逐日GPP和Re的均值呈先升高后降低的单峰变化趋势,逐日NEE的均值呈现先下降后上升的“V”型变化趋势(图2)。十四年的月GPP在生长季6、7、8、9月的均值分别为(102.6±16.9)、(172.2±23.2)、(150.5±20.9)、(76.6±9.9) gCm-2month-1,整体上看在7月高寒灌丛的光合生产能力最强;6、7、8、9月Re的均值为(69.9±11.1)、(100.3±21.6)、(98.3±20.4)、(72.5±12.6) gCm-2month-1,说明在7月高寒灌丛的生态系统呼吸最强,但是7月和8月的Re差别不大;月NEE在生长季6、7、8、9月的均值分别为(-32.8±9.8)、(-71.9±14.5)、(-52.4±10.0)、(-4.2±8.6) gCm-2month-1,由此说明在7月高寒灌丛生态系统的碳汇能力最强。整个生长季总GPP、Re和NEE平均值分别为(501.9±60.2)、(340.7±54.4)、(-161.2±30.1) gCm-2,高寒灌丛在生长季为碳汇。
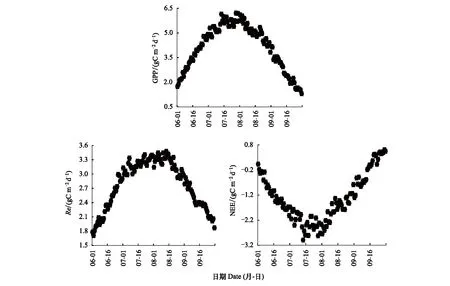
图2 高寒灌丛在2003—2016年生长季(6—9月)的平均(GPP, gCm-2d-1)、(Re, gCm-2d-1)和(NEE, gCm-2d-1)Fig.2 The average of gross primary production (GPP, gCm-2d-1), ecosystem respiration (Re, gCm-2d-1), and net ecosystem CO2 exchange (NEE, gCm-2d-1) of alpine shrubland in the growing season (June—September) from 2003 to 2016
由表2可见,在生长季6、7、8、9月,都表现为GPP对NEE的变化影响更强,并且在6月GPP对NEE的控制相对最强(r2=0.78)。同时,线性回归分析也表明,高寒灌丛生态系统NEE的变化主要受到GPP(GPP= -1.13NEE+2.64,r2=0.74,P<0.001)的控制,受Re(Re= -0.15NEE+2.61,r2=0.05,P<0.001)的影响相对较弱(图3)。
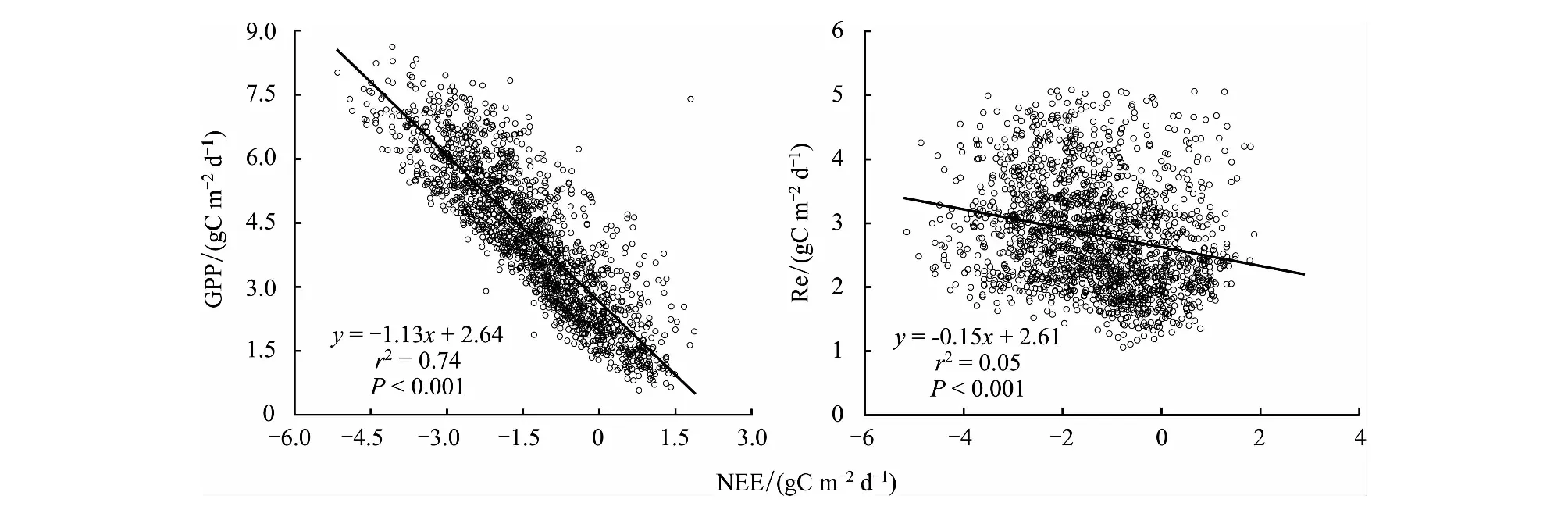
图3 高寒灌丛在生长季的逐日NEE与GPP、Re的关系Fig.3 The Relationship between daily NEE and GPP, Re of alpine shrubland in the growing season
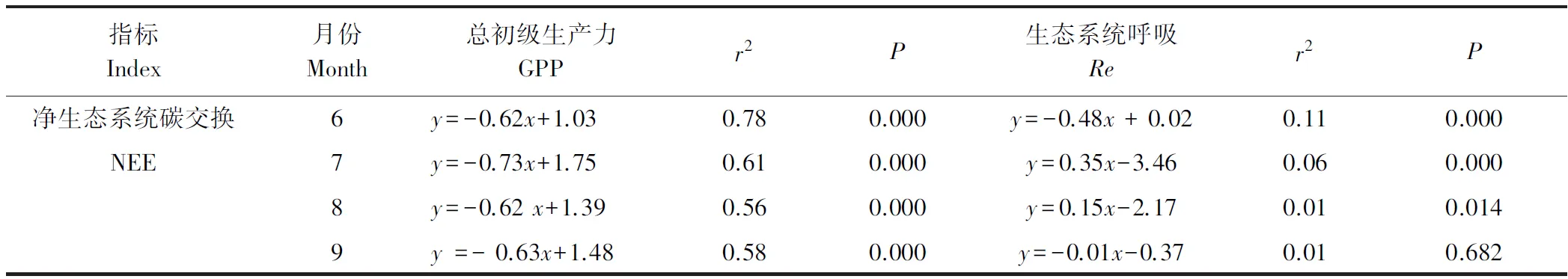
表2 生长季每个月的日GPP、Re与NEE的线性回归Table 2 Linear regression between daily GPP、Re and NEE of each month in growing season
2.3 高寒灌丛在生长季的不同月份日较差对CO2通量的影响
表3可见,GPP与MaxTa在生长季的各月都呈极显著正相关(P<0.001),与MinTa在6、7、9月显著正相关(P<0.05),与ADT只在6月没有显著相关性。通过比较“r2”值,说明GPP最主要的控制因素为MaxTa,并且在植被生长巅峰期(7、8月)相关性更显著。此外,线性回归分析表明,月GPP与月MaxTa显著正相关(r2=0.76,P<0.001),生长季GPP与生长季MaxTa显著正相关(r2=0.41,P=0.01)。Re与MaxTa、MinTa在生长季的各月都呈极显著正相关(P<0.001),与ADT在7月和9月极显著相关(P<0.01),也可看出Re最主要的控制因素是MinTa,在7、8月相关性较弱,在6、9月相关性较强。此外,线性回归分析表明,月Re与月MinTa显著正相关(r2=0.41,P<0.001),生长季Re与生长季MinTa没有显著相关性(r2=0.20,P=0.11)。
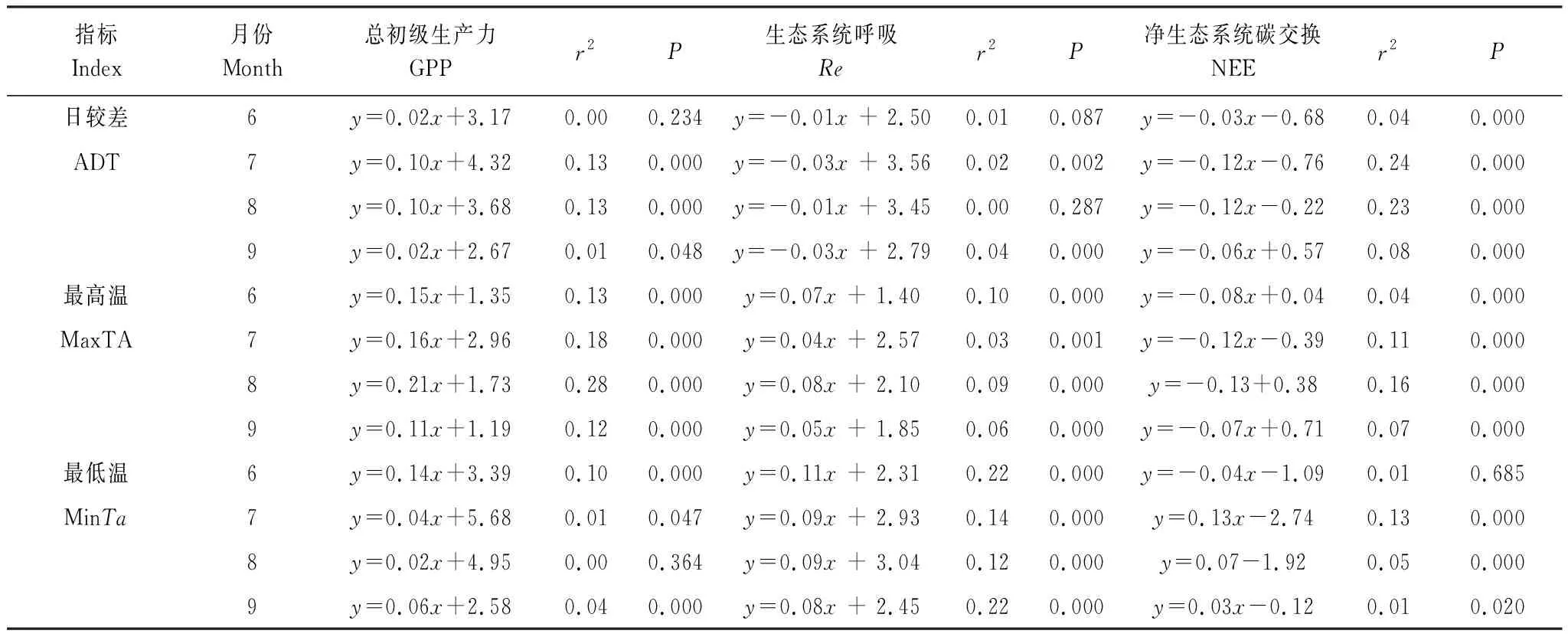
表3 逐日MaxTa、MinTa、ADT与GPP、Re、NEE的线性回归Table 3 Linear Regressions between daily GPP, Re, NEE and ADT, MaxTa, MinTa
NEE与MaxTa、ADT在生长季的各月都呈极显著负相关(P<0.001),NEE与MinTa在7月、8月、9月显著正相关(P<0.05)。通过比较“r2”值,可看出NEE最主要的控制因素是ADT,在7、8月相关性较强,在6、9月相关性相对较弱。由此说明,高寒灌丛生态系统ADT的增大有利于植被碳水化合物的积累,并且在植被生长巅峰期(7、8月)主要是通过光合作用的增强来实现碳水化合物的积累,而在生长初期(6月)和生长末期(9月)主要是通过呼吸作用的减弱来实现碳水化合物的积累。
此外,对整个生长季的逐日数据进行回归分析发现,GPP与MaxTa呈极显著正相关(P<0.001),Re与MinTa呈极显著正相关(P<0.001)(图4)。但是NEE与ADT呈极显著的二次曲线关系(P<0.001)(图4),阈值为16.4℃,当日较差大于16.4℃时,ADT越大越不利于植被有机质的积累;此外,线性回归分析表明,月NEE与月ADT,以及生长季NEE与生长季ADT都没有显著相关性(P>0.05),这种结果可能是由于生长季不同时期的气候环境因素之间的差异造成的。但是,整体上看,仍能说明高寒灌丛生态系统ADT的增大有利于植被光合生产能力的提高。

图4 高寒灌丛在生长季的逐日GPP与MaxTa、Re与MinTa、NEE与ADT的关系Fig.4 The relationship between GPP and MaxTa, the relationship between Re and MinTa, the relationship between NEE and ADT of alpine shrubland in the growing season
3 讨论
研究结果表明,与MinTa和ADT相比,MaxTa对灌丛生长季GPP变化起着相对更强的主导作用。潜在的原因可能是生长季热量条件对高寒生态系统植物的光合性能有很强的直接影响[28- 30],使高寒生态系统植被的代谢对温度波动具有充分的表型可塑性[3,6,31]。此外,温度能够影响土壤中微生物的生物量、酶活性,由于青藏高原高寒生态系统的土壤中含有大量土壤有机质[22],并且微生物分解土壤有机质的过程对温度极其敏感,较高的温度能够促进土壤有机质的分解,进而为植被的生长提供更多的养分,因此间接刺激了植被提高其光合生产能力,所以在一定冠层叶面积指数的水平上,温度对高寒生态系统的光合作用具有关键控制[7]。此外,在本研究附近的高寒草甸生态系统,研究了连续7 d极端高温造成的夏季热浪促进了正午GPP的提高,这个研究结果也能说明GPP与MaxTa的正相关关系[32]。总之,在未来气候变暖的背景下,将会提高高寒灌丛植被的光合生产能力。
有研究表明,夜间温度的升高会促使植物夜间消耗更多的碳水化合物,这将刺激植物在接下来的几天的光合作用得到加强,也就是“光合补偿”[14,16]。这个研究结果暗示ADT的减小可以间接促进GPP的提高。然而,我们的研究发现相反的结论,即在高寒灌丛生态系统中,ADT的增加可以促进高寒灌丛中的GPP的提高(表3)。这可能由于青藏高原的海拔高、温度低,夜间呼吸消耗较少的碳水化合物,不足以导致光合作用的过度补偿。此外,ADT与MaxTa呈显著正相关(P<0.001),这可能间接导致GPP与ADT之间存在相关性。同时,也可看到在6月ADT与GPP相关性不显著,这可能是由于生长季初期气温较低,植被生长缓慢,MaxTa与MinTa对GPP影响的差异较小造成的,由6月份MaxTa、MinTa与GPP线性方程的斜率(0.15 vs 0.14)相近(表3)可以证实这个结果。
生态系统呼吸(Re)是陆地上最大的碳通量之一,因此Re的微小波动会导致大气中CO2浓度的显著变化[33]。研究发现,日MinTa较MaxTa对Re的主导作用更强 (表3)。然而,过去的研究表明Re对气候变化的响应是高度可变的[33-34]。许多研究表明,增温显著增强了自养呼吸,但抑制了异养呼吸,导致温度变化对Re没有显著影响,表明Re通过调节其内源成分的响应具有抵御气候变暖的潜力[35]。Re是由自养呼吸和异养呼吸共同决定的,由于在不同生态系统自养呼吸和异养呼吸对气候变化的响应存在差异,因此导致Re对气候变暖响应的不确定性[35]。但以往的研究往往忽略了气候变化引起的MaxTa和MinTa的变化对生态系统呼吸的影响。研究发现MinTa与Re呈极显著正相关,说明MinTa的增加将促进未来气候变化下Re的提高。由于低温限制了青藏高原生态系统土壤有机质的分解,导致土壤中含有大量的土壤有机碳[6,21]。因此,夜间土壤呼吸可能对温度极其敏感,MinTa与生态系统呼吸存在显著的正相关关系,说明高寒灌丛生态系统的夜间呼吸对生态系统呼吸具有至关重要的作用。在6月和9月MinTa和Re的相关性更显著,这可能是由于这段时间的温度较低,生态系统呼吸相对较弱,所以夜间温度的变化引起夜间呼吸的变化对整个生态系统呼吸的影响更加显著。
研究发现,ADT在调节逐日NEE变化中起主导作用(表3)。本研究认为,青藏高原高寒灌丛在一定程度上ADT的增大有利于有机物质的积累,这与古松[7]和赵亮[21]的研究结果一致。本研究中,由表3可见,在6、7、8和9月的ADT与NEE都呈显著的负相关,但是由于不同月份降水、温度等因素之间的差异,导致不同月份的ADT与NEE差异较大,导致整个生长季的ADT与NEE的关系相对较为复杂,所以整个生长季的日ADT与日NEE呈二次曲线关系(图4)。但是,整体上看,研究结果仍能说明青藏高原高寒灌丛在生长季ADT的增大有利于生态系统碳的积累。此外,在7月和8月,雨热同期现象最为明显,最高温度和最高降水量导致植被光合作用在这个时期最强,因此植被的固碳能力主要是通过增强光合作用来实现的[6]。而在生长季初期6月和生长季末期9月,温度较低,光合作用较弱,因此碳汇能力更依赖于对呼吸作用的抑制[6,21]。这表明,在生长季的不同月份,在不同的水热条件下,植被可能以不同的方式来实现积累碳水化合物。
当前的大多数地球系统模型表明,陆地生态系统会因气候变暖而出现净碳损失,从而可能加剧气温上升[36-37]。大量研究表明,全球气候变暖背景下的MinTa的增长速度快于MaxTa,即未来ADT将会下降[18-19],暗示青藏高原高寒灌丛生态系统在未来气候变化的背景下,ADT可能会间接促进气候变化造成的碳损失。此外,在不同的水热条件下,ADT可能对不同生态系统的碳通量产生不同的影响,因此需要在更多的生态系统中评估ADT对CO2通量的影响,以更加准确的预测未来气候变化对碳收支的影响。
4 结论
(1)2003—2016年的生长季中,MaxTa和MinTa均呈现先升高后降低的单峰变化趋势,ADT在生长季的波动较小,没有呈现出明显的变化趋势,说明在青藏高原高寒灌丛ADT的变化与温度的变化并不同步。
(2)高寒灌丛在生长季为碳汇,ADT的增大有利于高寒灌丛碳水化合物的积累,并且在生长季的不同时期实现碳水化合物积累的方式不同。即,在植被生长巅峰期(7、8月)主要通过增强光合作用来实现碳水化合物的积累,而在植被生长初期(6月)和生长末期(9月)碳水化合物的积累更依赖于降低呼吸作用。
(3)未来气候变暖的背景下,MinTa的增长速度快于MaxTa,即ADT的减小将会间接削弱高寒灌丛生态系统的碳汇能力。
