花生结瘤起始因子基因鉴定及其对氮肥的响应
2021-01-14田丽彬刘译阳张佳蕾李荣冲崔凤万书波李国卫
田丽彬刘译阳张佳蕾李荣冲崔 凤万书波李国卫*
(1.山东师范大学生命科学学院,山东 济南250014;2.山东省农业科学院生物技术研究中心/山东省作物遗传改良与生态生理重点实验室,山东 济南250100)
氮是空气中含量最丰富的元素,约占空气总体积的78.1%,但通常情况下空气中的氮气不能直接被植物吸收利用,现代农业依赖工业氮肥提高作物产量。 近年来发现大量使用工业氮肥存在种种弊端,除成本高昂外,还会污染环境。 于是,经济环保的生物固氮越来越引发关注[1]。
生物固氮是解决农田生态环境污染、减少氮肥投入的可行性方法之一。 豆科植物与根瘤菌的共生固氮能力居各固氮体系榜首[2]。 固氮遗传通路中的重要组成成分结瘤起始因子NIN 是最早发现的与结瘤相关的转录因子,在结瘤早期发挥重要调节作用,同时发现NIN 的同源蛋白NLP(NIN-like protein)也与氮源利用有关[3-4]。 Schauser等[5-6]最早在豆科植物百脉根突变体中鉴定出NIN,发现它能调控根瘤菌感染线形成和原基细胞启动,随后在豌豆(Pisum sativum)中鉴定发现了NIN同源基因Sym35,又在拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)中发现众多NIN-like基因[7-10]。
花生是我国重要的油料作物,种植范围广泛。花生与根瘤菌互作形成根瘤,根瘤固氮是花生吸收利用氮的重要方式。 然而,生产中氮肥的过量使用形成氮阻遏效应,抑制根瘤的形成,限制固氮作用的发挥[11]。 研究结瘤起始因子基因及其对氮肥的响应对深入研究花生根瘤形成具有重要意义。因此,深入研究花生NIN基因家族及其在不同种植模式下对氮肥的响应,有利于了解结瘤因子在根瘤发育中的调控作用,进而明确其在花生根瘤固氮体系中的功能,为提高花生固氮能力和减轻氮肥投入提供科技支撑。
1 材料与方法
1.1 基因检索
在phytozome(https://phytozome.jgi.doe.gov/pz/portal.html)数据库中检索大豆(Glycinemax)、水稻(Oryza sativa)、拟南芥(Arabidopsisthaliana)的NIN 蛋白序列。 利用同科植物大豆NIN蛋白序列,在PeanutBase(https://peanutbase.org/)中运行BLASTP程序筛选出花生NIN 蛋白序列,以满足E-value<e-20的标准筛选出目标蛋白序列,以FASTA 格式进行保存。
1.2 生物信息学分析
通过Clustalx软件比对拟南芥、大豆、水稻和花生NIN蛋白序列后,使用Mega5.2软件,采用邻接法(neighbor-joining)构建进化树[12];利用GSDS(http://gsds.cbi.pku.edu.cn/)在线软件分析基因结构[13];利用MEME(http://meme-suite.org/)在线软件分析蛋白保守结构域。 利用花生转录组数据(NCBI:PRJNA354652),使用Cuffdiff软件[14]计算花生NIN基因家族在不同组织的表达量(FPKM 值),利用HemI软件进行表达模式分析[15]。
1.3 花生NIN 基因在不同施氮条件下的表达分析
供试品种为花育22。 试验设4个处理,花生单作不施氮肥(N0)、花生单作施90 kg/hm2氮肥(N90)、花生//玉米间作不施氮肥(IN0)和间作施90 kg/hm2氮肥(IN90)。 在始花期花生取新鲜根,清洗后液氮速冻,不同地块分别取样3次。 提取RNA 进行转录组测序,用cv.Tifrunner基因组作为参考基因组,使用Cuffdiff软件计算RPKM 值,分析NIN基因家族的表达丰度,利用HemI软件进行表达模式分析。
2 结果与分析
2.1 花生NIN 基因鉴定与系统进化分析
使用大豆NIN转录因子的蛋白序列在花生数据库PeanutBase中进行基因检索,得到栽培种花生中33个NIN转录因子的蛋白序列(表1)。 分析结果发现,33个花生NIN基因不均匀地分布于14条染色体上。 其中,13号染色体上分布最多,存在5个NIN基因;其次为01 号染色体,存在4 个NIN基因;03、08、17和20号染色体上各有3个;05、10、11和15号染色体上各有2个;其余4条染色体上各有1个。 其中,Arahy.F3ZCPW和Arahy.ZR07MJ为一对等位基因。
为进一步了解花生NIN 与其他物种NIN 的系统进化关系,利用拟南芥(14个)、水稻(13个)、大豆(28个)和花生(33个)的NIN 转录因子蛋白构建进化树(图1)。 根据亲缘关系远近将NIN分为A、B、C和D 4组,其中A 组又进一步分为A1~A4 4个亚组。 A组中花生NIN基因最多,共14个,A1和A3亚组各含5个花生NIN基因,A2和A4亚组各含2个;B组中花生NIN基因次之,为11个;C组和D组各含4个NIN基因。 不同物种NIN基因分散于4组中,并未因物种差异而单独聚集,说明花生与其他物种NIN基因之间亲缘关系较近。
2.2 花生NIN 基因结构分析
对33个花生NIN基因结构分析,发现多数花生NIN基因的外显子数目为4~6个,其中14个基因含有4个外显子,10个基因含有5个外显子,6个基因含6个外显子。 花生NIN基因的外显子长度和内含子数量相似,说明花生NIN基因在进化上保守。 其中,Arahy.1E9R5B、Arahy.L1HKP、Arahy.ILQ5AU和Arahy.P2WFEB变异较大。Arahy.1E9R5B和Arahy.L1HKP发生了外显子增多的变异,其中Arahy.1E9R5B含有10个外显子,并且内含子长度明显增长,其编码的氨基酸数目也较其他的多;Arahy.L1HKP含8个外显子,内含子长度增长,但其外显子长度减少,导致编码氨基酸数目并未增多。Arahy.ILQ5AU和Arahy.P2WFEB发生了外显子减少的变异,二者均只含有1个外显子,导致其编码氨基酸数目减少 (图2)。
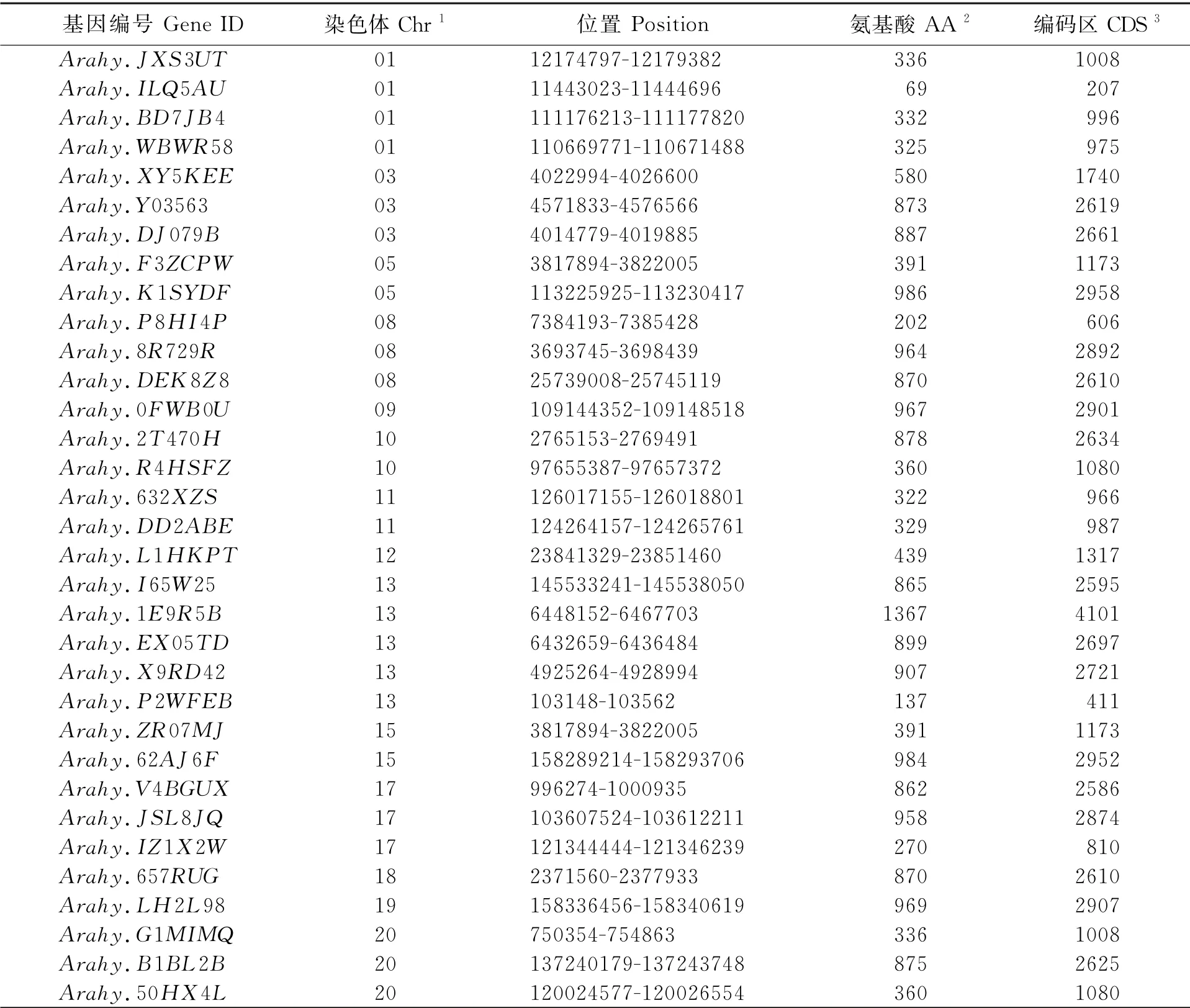
表1 花生NIN 基因家族生物信息学分析Table 1 Bioinformatics analysis of NIN family in peanut
2.3 花生NIN 转录因子保守结构域分析
利用MEME在线软件搜索花生NIN 转录因子的保守结构域,结果发现花生NIN 转录因子具有10个保守结构域,分别命名为motif 1~motif 10(图3)。 同一组的花生NIN 家族成员具有相似的保守结构域,不同组的花生NIN 家族成员的保守结构域相差较大。 所有的花生NIN 转录因子中均含有motif 7。 经鉴定,motif 7 为RWP-RK(PF02042,一段保守的60个氨基酸长度的蛋白质序列,可与DNA 结合),该序列是NIN 转录因子中最典型的结构,广泛存在于其他物种NIN 转录因子中, 是识别NIN 家族成员的统一标准[16-17]。 A 组中只含 有motif 1 和motif 7 两 个保守结构域。 B组中保守结构域最多,其中motif 9和motif 10是其所特有的,除Arahy.XY5KEE和Arahy.L1 HKPT 外,B 组花生NIN 转录因子序列含有特殊的motif 10。 B 组 和C 组 都 存 在motif 3,鉴定为PB1(Protein binding 1),推测其可能为蛋白质识别结合结构域。
2.4 花生NIN 基因组织表达分析
利用19个花生组织的转录组数据绘制基因表达热图,分析NIN基因在花生不同组织中的表达情况,推测该基因在花生中的功能(图4)。 分析发现,Arahy.I65W25和Arahy.V4BGUX两个基因在花生根和根瘤表达量最高,说明这两个基因与早期根瘤形成有关,与之前报道过的其他物种NIN基因在结瘤早期发挥作用结果一致[18]。 除上述2个基因在根与根瘤表达量高外,Arahy.2T470H、Arahy.1E9R5B、Arahy.LH2L98和Arahy.0FWB0U在根和根瘤也有较高表达,推测上述4个基因与早期根瘤形成也有一定关系。Arahy.62AJ6F、Arahy.K1SYDF、Arahy.LH2L98和Arahy.0FWB0U在 花生各个组织中均有表达,推测其表达具有时空特异性,参与花生各个部位氮素利用的调控。 其余基因在检测的组织中表达量较小,推测这些基因可能在其他未检测组织表达。
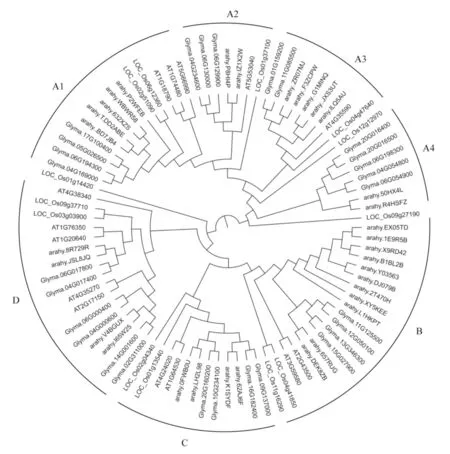
图1 花生、大豆、水稻和拟南芥NIN 家族进化树Fig.1 The phylogenetic analysis of the NIN family in peanut,soybean,rice and Arabidopsis

图2 花生NIN 家族基因结构Fig.2 Gene structure of NINs in peanut
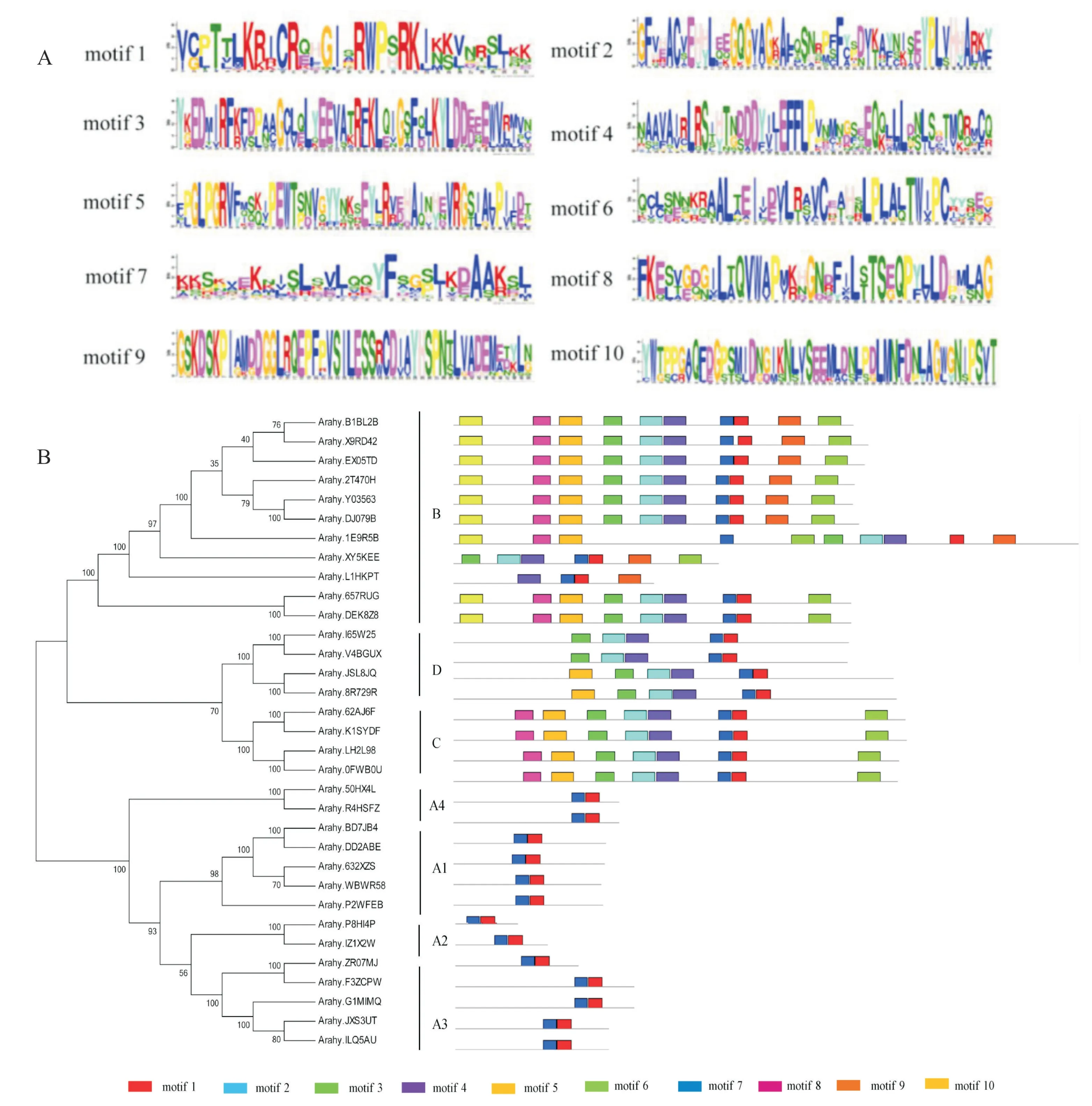
图3 花生NIN 家族保守结构域Fig.3 Conserved motifs of NIN family members in peanut
2.5 花生NIN 基因在不同施氮条件下的表达分析
为进一步分析花生NIN基因对不同氮素的响应,利用转录测序分析了花生单作和花生//玉米间作两种模式下始花期花生根系中NIN基因对氮肥的响应。 与图4结果一致,该基因家族在根中的表达丰度都比较低,检测到有10个基因在4种生长条件下均没有表达,但Arahy.K1SYDF、Arahy.DEK8Z8、Arahy.62AJ6F和Arahy.657RUG在4种处理下表达量高。 另外,检测到12个基因在两种条件下表达丰度发生变化。 单作条件下,Arahy.0FWB0U和Arahy.2T470H的表达受氮素诱导,表达量升高,Arahy.DEK8Z8、Arahy.657RUG、Arahy.8R729R和Arahy.JSL8JQ的表达受氮素抑制,表达量降低;间作条件下,Arahy.K1SY0F、Arahy.8R729R、Arahy.2T470H和Arahy.62AJ6F的表达受氮素诱导,表达量升高,Arahy.0FWB0U和Arahy.657RUG的表达受氮素抑制,表达量降低(图5)。
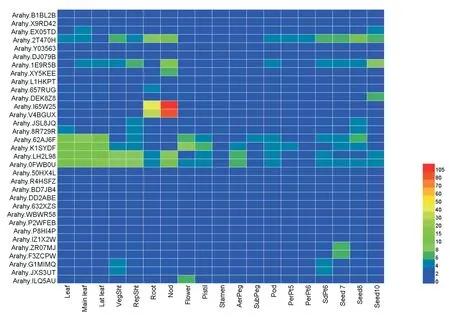
图4 花生NIN 基因组织特异性表达分析Fig.4 Tissue-specific expression of NIN gene family in peanut

图5 花生NIN 基因在不同氮肥情况下的表达分析Fig .5 Expression analysis of peanut NIN gene under different nitrogen fertilizers
3 讨 论
生物固氮是解决氮肥造成的环境污染问题的重要方法之一。 NIN转录因子是根瘤固氮遗传通路上的重要组成成分,在结瘤早期发挥重要作用,进一步研究发现,尽管其功能不同,但都与氮素利用有关。在没有根瘤菌侵染的情况下,NIN 转录因子能够诱导根瘤皮层细胞分裂[19];在不同氮素条件下,NLP7能提高植物光合速率和氮同化能力,增加植物的生物量,而nlp7突变体则表现出明显的氮缺乏表型和抗旱能力[9,20];NLP8通过响应硝酸盐信号促进种子萌发[21],这说明NIN及其同源蛋白在植物根瘤形成与根系氮素利用方面发挥重要作用。 因此,研究花生NIN基因的功能,能够明确其在花生根瘤固氮体系中的作用,探索提高花生根瘤固氮能力,减少氮肥施用量,解决氮肥过度使用造成的环境污染问题。
本研究利用生物信息学技术对花生NIN基因的理化性质、进化关系、基因结构、表达模式等进行了初步分析。 花生NIN基因按亲缘关系远近分为4组,其他物种NIN基因分散于各组中。 A3亚组中Arahy.ILQ5AU仅编码69个氨基酸,形成RWPRK保守结构域,且该结构域为NIN基因家族所共有的,由此推测花生NIN基因通过RWP-RK 保守结构域起始根瘤形成,调节氮素利用[16,19]。 基因结构和保守结构域分析表明,花生NIN 家族外显子数目及长度相似,并且同一组的花生NIN 转录因子具有相似的保守结构域,说明同组的花生NIN基因可能具有相似的功能。 花生NIN基因家族成员较多,说明该家族可能在花生不同部位发挥不同功能,Arahy.I65W25和Arahy.V4BGUX两个基因在花生根和根瘤处表达量相对较高,推测它们可能是花生中主要参与根瘤形成的基因,进一步验证了NIN转录因子在根瘤形成早期中的作用[18]。Arahy.F3ZCPW和Arahy.ZR07MJ在花生籽仁中相对表达量较高,推测其参与硝酸盐促进种子萌发过程,与先前报道过的拟南芥At NLP8 的表达模式一致[21-22]。Arahy.657RUG、Arahy.DEK8Z8、Arahy.K1SYDF和Arahy.62AJ6F等4个基因在不同种植模式以及不同施氮条件下表达量变化较明显,说明这4个基因可能参与了不同种植模式及氮肥条件下根瘤发育的调控。 上述研究为挖掘花生根瘤固氮潜力,提高花生对氮肥的利用率,从而解决氮肥过量使用造成的环境污染问题奠定了理论基础。
