醒斑银弄蝶Carterocephalus alcina Evans, 1939与其近似种(鳞翅目: 弄蝶科: 链弄蝶亚科)的关系
2021-01-09侯永翔李宇飞千葉秀幸范骁凌
侯永翔,李宇飞,王 敏,千葉秀幸,范骁凌
(1 华南农业大学 植物保护学院,广东 广州 510642;2 西安交通大学 公共卫生学院,陕西 西安 710061;3 檀香山毕肖普博物馆,美国 夏威夷 96817 - 0916)
弄蝶科Hesperiidae昆虫是一类色泽比较单一,翅面斑纹变化较大的蝴蝶,其种间、属间,甚至亚科间有模拟现象,表现出翅面斑纹的高度相似性,而有的种类雌、雄或种下存在明显的差异。越来越多的研究表明,仅依据成虫的形态特征,尤其过分依赖翅面斑纹的传统分类导致的误判与分类地位的混乱普遍存在[1-10]。
银弄蝶属CarterocephalusLederer, 1852 隶属于弄蝶科链弄蝶亚科Heteropterinae,目前已知约19种,分布于亚洲、欧洲和北美洲,在中国均有分布,其中12种为我国特有。成虫活跃于高山草甸及林窗地带,喜访花和在潮湿地面吸水;幼虫以雀麦属Bromus、洋狗尾草属Cynosurus等禾本科植物为食[11]。
醒斑银弄蝶C.alcina是Evans于1939年以产自云南腾冲的标本为模式描述的[12]。李传隆[13]以产自云南昆明西山的标本为模式描述了黄斑银弄蝶C.alcinoides,之后的多数研究中仅有黄斑银弄蝶的记录[11, 14-21];另外,将其他种误定为该种的也不少见[11, 22]。近来我们在研究银弄蝶时发现,2头产自北京的标本与醒斑银弄蝶非常相似,但又存在明显差异,有研究将其误定为黄斑银弄蝶[18-19, 23-24]。
本研究的目的旨在系统研究银弄蝶形态学的基础上,以醒斑银弄蝶及相近物种为主要研究对象,综合其形态特征、COI序列和地理分布等信息,厘清它们之间的关系。
1 材料与方法
1.1 试验材料
根据Evans[25]、武春生等[11]的分类结果,本研究以醒斑银弄蝶、黄斑银弄蝶为主要研究对象,为了更好地解决疑难种的分类及近似种的关系问题,研究材料包括这2个种不同分布地,同一分布地斑纹变异的不同个体以及近似斑纹的其他种,共9种,32 个个体 (表 1),根据近年弄蝶科系统发育研究[10, 26-27],选取链弄蝶亚科Heteropterinae中4个不同属的物种:链弄蝶Heteropterusmorpheus(Genebank 序列号:KF881050)、小弄蝶Leptalinaunicolor(Genebank序列号:AB192500)、糜弄蝶Metisellametis(Genebank序列号:KY019796)、胆弄蝶Dardarinadardaris(Genebank 序列号:GU161460)为外群。除GeneBank下载的序列外,其余均为本研究测序结果,并已上传NCBI,具有标本编号的材料均保存于华南农业大学昆虫学系昆虫标本馆(表1)。
1.2 试验方法
1.2.1 形态学研究 选取代表性标本展翅后用SONY DSC - RX100 拍照,将其腹部完整取下,用100 g/L的NaOH溶液加热处理,检视其外生殖器,采用Keyence VHX-5000超景深三维显微系统拍照。稳定的斑纹、外生殖器特征是鉴定种的标准,采用Fan等[3]的形态术语。
1.2.2 COI序列的获取 总 DNA 的提取使用天根生化科技(北京)有限公司的血液/细胞/组织基因组DNA 提取试剂盒 (DP304)。COI序列 (658 bp)扩增与测序引物多为LCO1490和HCO2198[28],部分扩增困难的标本,根据黄翅银弄蝶C.silvicola[29]、醒斑银弄蝶C.alcina[27]全线粒体基因组及本研究已获得的序列,采用新设计的1对引物(CarF:5′-CGCGCTACTAATCATAAAGATATTG G-3′ 和CarR:5′- CTTCAGGGTGTCCAAAAAATCA-3′)。PCR 扩增采用 20 μL 体系,包括 1 μL DNA 模板,各 0.8 μL(10 μmol/L)的上、下游引物,10 μL 的 2 ×EasyTaqPCR superMix(北京,全式金生物技术有限公司)。PCR 扩增程序为 94 ℃ 预变性 4 min;94 ℃变性 30 s、48 或 49 ℃ 退火 45 s,72 ℃ 延伸 1.5 min,循环35或40次;72 ℃延伸7 min。测序由上海生工生物工程股份有限公司完成。
1.2.3 数据分析 序列检查、拼接及遗传距离的计算使用MAGE 7.0软件[30]。系统发育树构建采用最大似然法 (Maximum likelihood,ML)和贝叶斯分析法 (Bayesian inference,BI)。其中 ML 使用 RAxMLHPC2 on XSEDE 8.2.12 软件[31],选择 GTR+G 模型,重复抽样 1 000 次,以自展值 (Bootstrap support values, BS)评估其节点的稳定性;BI使用MrBayes on XSEDE 3.2.7a 软件[32],采用可逆跳跃马尔可夫链蒙特卡罗 (Reversible-jump markov chain monte carlo,MCMC)策略,运行 5×106代,每 1 000 代抽样 1 次,用后验概率 (Posterior probabilities, PP)评估其分支的稳定性。2种分析均在CIPRES 线上服务平台上完成(https://www.phylo.org/)[33]。

表 1 本研究银弄蝶属Carterocephalus的标本信息1)Table 1 The specimen information of Carterocephalus in this study
采用 Fig Tree 1.4.4 和 Photoshop CS5 软件显示与编辑系统发育树。
2 结果与分析
2.1 分子系统发育分析
基于ML、BI构建的系统发育树基本一致,仅部分节点的支持值存在差异(图1)。系统树显示银弄蝶属Carterocephalus单系并得到较强支持(BS=91, PP=1);研究材料聚为明显的 9 支,分别对应形态上的基点银弄蝶C.argyrostigma、射线银弄蝶C.abax、银线银弄蝶C.patra、前进银弄蝶C.avanti、白斑银弄蝶C.dieckmanni、愈斑银弄蝶C.houangty、黄翅银弄蝶C.silvicola与银弄蝶C.palaemon、Clade A 和醒斑银弄蝶C.alcina。其中射线银弄蝶与银线银弄蝶,前进银弄蝶与白斑银弄蝶的姐妹关系得到强烈支持;黄翅银弄蝶、银弄蝶、Clade A和醒斑银弄蝶聚为高度支持的单系支(BS=98, PP=1)。在这一支中,银弄蝶C.palaemon非单系,并与黄翅银弄蝶C.silvicola聚为有强烈支持的一支(BS=98, PP=1);昆明西山的2个样本,即黄斑银弄蝶C.alcinoides没有独立成支,而是嵌套于醒斑银弄蝶C.alcina中,结合形态特征与地理分布,提出黄斑银弄蝶C.alcinoidesLee, 1962是醒斑银弄蝶C.alcinaEvans, 1939 的异名;北京产的 2 个个体独立聚成一支(Clade A),综合形态特征及遗传距离,Clade A应该是1个新种,即长斑银弄蝶CarterocephaluslongimaculatusHou, Fan & Li sp.nov.。
2.2 长斑银弄蝶Carterocephalus longimaculatus Hou, Fan & Li sp.nov.
http://zoobank.org/A68ADACD-5893-41F1-AED8-00F72BE5995B

图 1 基于COI条形码以最大似然法构建的银弄蝶属的系统发育树Fig.1 Phylogenetic tree of Carterocephalus based on COI barcodes using Maximum likelihood analysis
Carterocephalussp.Yangetal., 1994: 54[34].
Carterocephalusalcinus: Seto, 2013: Plate37[24].
Carterocephalusalcinoides: Wangetal., 1998:188[18]; Wang, 1999: 277[19].
鉴别特征:外观与醒斑银弄蝶C.alcina相似,其主要不同在于前翅中室基部具有1个三角形小斑;后翅Sc+R1室具有1个不规则方斑,端部M1-M2室具有1个长斑,延伸至亚缘。

图 2 2种银弄蝶成虫Fig.2 Adults of two Carterocephalus species
雄性 (图 2A、2B):触角长约 7 mm;下唇须第2节粗、腹面被黄色长毛,第3节尖细、前伸。前翅长13~14 mm,前缘中部没有明显凹入;翅正面深褐色,斑纹橙黄色;中室端斑大,近方形,前缘具有1个较宽条状斑纹,与中室斑相连,终止于R2室基部,中室基部有1个三角形斑,与前缘斑相连或不相连,翅端部R3-R5室、M1-M2室2个方形大斑相连,R4、R5外缘常有 2 个小斑,M3、CuA1、CuA2室斑相连呈1条宽短带,CuA2室近基部有1个水滴状黄斑,向基部渐窄;反面除外缘具有1列浅黄色小斑、中室端脉淡黄外,与正面基本相同。后翅正面底色、斑纹颜色同前翅,端部分别于M1、M2,CuA1和CuA2室有3个黄斑,其中M1、M2室的最大并延伸近外缘,并在外1/3处有缢缩状,M3室有时可见1个极小斑,Sc+R1室具有1个不规则的方斑,中室基部具有1个水滴形斑,向基部渐窄;反面底色浅褐色,与正面斑对应位置的斑浅黄色,其余奶白色,外缘及亚缘具有2列小斑,Sc+R1室、CuA2室基部均具有1个长斑,中室斑与M1、M2斑之间具不规则的条形斑。前翅缘毛深褐色,后翅外缘毛橙黄色。
雄性外生殖器(图3A、3B):钩形突狭长渐细,端部两裂;颚形突左右愈合,渐窄,末端稍短于钩形突端;囊形突很长,占基腹弧的一半以上;阳茎很长,长于整个基腹弧的骨环结构,阳茎端膜被小刺;阳基轭片环状且沿阳茎背腹面突出;抱器端两裂,其中背端突钝圆,腹端突末端突出,被小刺。
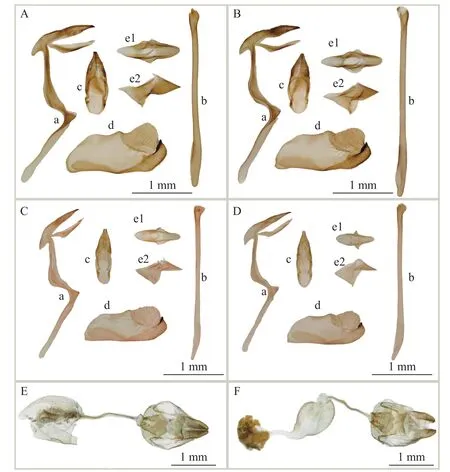
图 3 2种银弄蝶的外生殖器Fig.3 Genitalia of two Carterocephalus species
雌性:未见标本。根据北京蝶类原色图鉴118页图24a[34],前翅前缘橙黄长条及R4、R5室外缘的小斑缺如,中室基部斑小;后翅M1、M2室大斑不向外延伸、Sc+R1斑小,其余似雄性。
模式标本:正模♂,北京灵山,2017-Ⅵ-3,李鹏采;副模♂,北京小龙门,2014-Ⅴ-25。模式标本保存于华南农业大学昆虫系昆虫标本室。
词源:学名longimaculatus源于 longi+maculatus,意为后翅正面M1、M2室的长斑。
主要分布于中国辽宁(建平)、北京(灵山)、天津(八仙山)、河南(济源)等地。该种在北京地区的生境为林间空地及草地,成虫常被目击访花,发生期为5月初至7月中下旬。
2.3 黄斑银弄蝶是醒斑银弄蝶的异名
Evans[12]描述醒斑银弄蝶Carterocephalus alcina时除检视了腾冲的标本外,同时还检视了产自丽江与昆明的雌、雄标本共19头。李传隆[13]依据产自昆明西山的2雄1雌标本及河口的1雄标本描述了黄斑银弄蝶C.alcinoides。根据原描述后者与前者极为相似,但又不同,表现在“前翅正面前缘橙黄色长条斑粗且中部膨大;前翅反面4、5(M1、M2)室的斑相连,且内缘略成一条直线;雄性外生殖器钩形突左右分叉,互相紧贴,形似钢笔尖;颚形突左右连成一体,尖长略等于钩形突;雌蝶前缘长条斑缺如”[13]。事实上,与二者的模式种(图2C、2D)相比较,前翅前缘橙黄长条斑仅端部宽窄稍有差异,前翅反面M1、M2斑基本一致,而雌性根据Evans的描述[25]和黄斑银弄蝶一样,前翅前缘条缺如。另外通过检视15雄6雌标本,其中包括腾冲标本2头、昆明标本2头,发现不同分布地,甚至同一分布地的不同个体,雄性前翅前缘橙黄长条斑端部的宽窄有变异,M1、M2室斑的内缘成直线是不稳定的 (图 2E、2F、2G、2I、2J、2L、2M);雌性前缘条斑缺如 (图 2H、2K、2N);前翅端部 R3-R5室和 M1-M2室2个大斑通常雌性不相连(图2H、2K、2N),雄性有变异,有相连的、有稍微分离的,甚至有左右翅连接不一致的(图2E)。通过对斑纹有变异的外生殖器解剖发现,雄性外生殖器除分布于四川格西沟的抱器瓣腹端突稍钝外,腾冲、昆明及其他分布地的基本一致(图3C、3D)。雌性外生殖器基本一致,前后阴片对称,前阴片上缘浅的小V型,后阴片稍弧形凹入(图3E、3F),与袁峰等[35]所描述存在一定差异。系统发育树表明,昆明、腾冲及其他分布地的21个个体交织成高度支持的一支,且遗传距离为0~0.60%,其中,昆明与腾冲样本间遗传距离仅为0.15%;格西构分支与其余个体间遗传距离为0.60%,形态特征上,除了上述变异外中室基部有的个体可见小黄色斑纹(图2J、2L、2M),外生殖器无明显不同(图3C、3D)。综合形态特征、分子系统发育分析、遗传距离及地理分布信息,我们认为黄斑银弄蝶C.alcinoidesLee, 1962是醒斑银弄蝶C.alcinaEvans, 1939 的新异名 (syn.nov.)。
3 讨论与结论
银弄蝶属已知19种20余亚种,本文仅包括醒斑银弄蝶C.alcina及其近似种且无涉及亚种的材料,因此该属的单系性仍需增加物种数和分子数据进一步确认。
根据翅面斑纹,以前的研究认为醒斑银弄蝶C.alcina与银弄蝶C.palaemon、黄翅银弄蝶C.silvicola、愈斑银弄蝶C.houangty、射线银弄蝶C.abax(2亚种:指名亚种与银线亚种C.abaxpatra)、美丽银弄蝶C.pulchra相似,其中与射线银弄蝶C.abax的关系最近[12, 25]。Sugiyama[36]将银线亚种提升为种级分类单元,即银线银弄蝶C.patra。本研究结果支持该亚种的提升,同时显示银弄蝶C.palaemon、黄翅银弄蝶C.silvicola、长斑银弄蝶C.longimaculatussp.nov.与醒斑银弄蝶C.alcina为高度支持的单系群,且后翅具大而醒目的淡黄斑,M1、M2室斑与Sc+R1中部斑远离是其共同特征。基于基因组的研究也发现银弄蝶C.palaemon、黄翅银弄蝶C.silvicola和醒斑银弄蝶C.alcina成单系[10]。遗传距离分析显示文中银弄蝶种间的平均遗传距离为1.2%~10%,长斑银弄蝶C.longimaculatussp.nov.与其他种的平均遗传距离为3.9%~10%,其中与醒斑银弄蝶的遗传距离为3.9%。系统发育树中长斑银弄蝶C.longimaculatussp.nov.独立成支,尽管支持值不高,但2种方法均显示其在已知种中与醒斑银弄蝶的关系更近。黄翅银弄蝶C.silvicola与银弄蝶C.palaemon在系统发育树上聚为一支,其平均遗传距离为1.2%,但产自日本的银弄蝶C.palaemon并没有与欧洲的聚在一起,其是亚种还是种?银弄蝶C.palaemon已知有近10余亚种。Cong等[10]基于基因组数据认为C.palaemon mandan为独立种。因此这2个种及种下问题的澄清需要更广泛的取材进行进一步深入研究。
致谢:衷心感谢标本采集及提供者:李鹏、张珑,云南省农业科学院何成兴研究员,华南农业大学黄珍富、纪树钦、汤静、魏福宏、韩雨珂、缪志鹏、莫世芳、曹成龙和谭舜云。感谢华南农业大学洞穴生物实验室提供Keyence VHX-5 000超景深三维显微系统以完成外生殖器拍照。感谢伦敦英国自然历史博物馆Blanca Huertas博士,中国科学院动物研究所的武春生研究员与姜楠博士在模式标本检视过程中给予的热情帮助与大力支持。
