马尾松无性系木材基本密度和纤维形态的变异及选择
2021-01-08尹焕焕刘青华周志春万雪琴余启新丰忠平
尹焕焕,刘青华,周志春,万雪琴,余启新,丰忠平
(1.四川农业大学 林学院,四川 成都 610000;2.中国林业科学研究院 亚热带林业研究所,国家林业和草原局马尾松工程技术研究中心/浙江省林木育种技术研究重点实验室,浙江 杭州 311400;3.浙江省淳安县姥山林场,浙江 淳安 311700)
木材品质是通过木材密度、木材强度、纤维长度、纤维宽度、纤维长宽比、微纤丝角、纤维素含量等材性性状来定量评价的,其中,木材基本密度和纤维形态是纸浆材材性研究最基础的2个性状。木材基本密度影响力学强度,决定木材及纤维制品的产量和品质;纤维形态与纸浆质量和纸张强度密切相关[1]。马尾松Pinusmassoniana具有速生丰产、适应性强和材性优良等优点,是中国南方山地主要的针叶用材树种。马尾松木材主要用作造纸、建筑和板材等。作为用材树种,最重要的是生长和材性性状,其中材性性状是衡量木材品质的最重要指标[2]。ZOBEL等[3]也曾指出:在初步的树木选择中要特别强调木材质量,因为木材性状遗传相当稳定,因而能够取得良好的效果。因此,对木材性状进行遗传改良是人工用材林经营中提高木材品质的最根本措施。鉴于马尾松是重要的纸浆材树种,国内外学者研究了马尾松木材基本密度和纤维形态在株内、株间[4]、种源[5−6]、家系[7−8]及无性系间的变异,以及立地条件和经营措施[9−10]对木材基本密度和纤维形态的影响。以上研究表明:木材基本密度和纤维形态均存在着较大的遗传变异,在种源间、家系间、无性系间以及株间和株内不同部位均存在明显的变异。其中,木材基本密度的家系遗传力为0.46[11],无性系重复力为0.63[12],受较强的遗传控制。谭健晖等[8]通过对马尾松半同胞家系纤维形态遗传变异的研究,表明晚材的纤维长度和宽度的家系遗传力分别为0.42和0.51,认为家系水平的纤维改良潜力较大。但这些马尾松的材性研究主要集中在中幼龄期,而随着树木年龄的增加,材性变异规律也会随之发生变化。因此,对马尾松幼龄—成熟龄期木材基本密度和纤维形态的变异及相关研究至关重要。本研究以浙江省淳安县姥山林场34年生马尾松无性系试验林为研究对象,选取5个不同产地的50个无性系,研究了木材基本密度和纤维形态在产地间和产地内无性系间的遗传变异、性状间相关及幼龄—成熟期的相关性,同时选育材性优良的马尾松无性系,以提高选择育种的效率和性状改良的潜力。
1 材料与方法
1.1 材料来源
材料取自浙江省淳安县姥山林场34年生马尾松无性系试验林,选取广西、贵州、安徽、浙江和江西等5个不同产地50个代表性的健壮马尾松无性系进行调查。该试验林为双因素巢式分组设计,10次重复。株行距4.0 m×4.0 m。所在地年平均降水量1 430 mm,年平均气温17.0 ℃。土壤由中等肥力的黄土组成。
1.2 研究方法
2017年12月,每个产地选择10个无性系。对每个无性系,随机选择3个分株测量木材性状。2017年12月,利用长45 cm、直径6 mm的生长锥在每个单株胸高处南北向钻取贯穿髓心的完整无疵木芯,用以木材基本密度、纤维长和纤维宽等性状测定。
1.2.1 木材基本密度的测定 先将木芯浸入水中至饱和状态,然后对所取木芯自髓心向外,每5个年轮切成1段,每段编号为i(i=1,2,···,7)。用最大含水量法[13]测量第i段木芯的木材基本密度,然后利用第i段年轮宽度测定值估算其所代表的年轮段面积(Ai),进而计算整个木芯的加权平均密度。其中:Di为第i段木芯的木材基本密度,,M为饱和含水率时试样的质量(g),Mi为第i段木芯绝干时试样的质量(g),Dw为构成细胞壁的木材物质的比重,取平均值为1.53。
1.2.2 木材纤维形态的测定 木材样品的离析采用过氧化氢-冰醋酸法进行。每个无性系在木芯上按幼龄材、过渡年龄和成熟材取样。幼龄材、过渡龄材分别取自髓心向外第0~5年轮和第10~15年轮[14],成熟材在靠近树皮的5个年轮取样。随后加入1∶1的冰醋酸和双氧水离析液,再将试管进行水浴加热直至木芯发白变软,待试管冷却后将离析液倒掉,并水洗5次以上至样品无异味。用玻璃棒随机挑选适量样品于专用量筒中,并利用纤维疏解打散器将试样打散。然后,将充分打散的样品转移至专用测量杯中,并加水至100 mL。最后,设置好测量程序,采用维美德纤维图像分析仪(Valmet FS5)进行自动测量。仪器计算记录纤维平均长度(mm)和纤维平均宽度(μm)。
1.3 数据分析
所有性状通过以下线性模型进行方差分析:Xijk=μ+Pi+Cj(i)+εijk。其中:Xijk为第i个产地第j个无性系第k个表型的观察值,μ是总体平均值,Pi是第i产地的效应(i=1,2,3,4,5),Cj(i)是第i个产地内第j个无性系的效应(j=1,2,···,10),εijk是残差。除总体平均值外,产地间和产地内无性系的效应被认为是随机的[15]。
使用SAS 8.0软件的PROC GLM进行ANOVA分析以检验性状在产地间与产地内无性系间的差异。在上述线性模型的基础上,还估计了产地间(σp2)、产地内无性系间(σc2)和残差(σe2)的方差分量。同时,用SAS软件的PROC VARCOMP和PROC CORR NOSIMPLE分别估算其遗传和表型相关系数并做显著性检验。
2 结果与分析
2.1 马尾松不同产地无性系木材性状的遗传变异
方差分析结果(表1)显示:马尾松木材基本密度在产地间和产地内无性系间皆存在着极显著变异(P<0.001)。50 个无性系的木材基本密度均值为 0.461 g·cm−3,变化范围为 0.350~0.636 g·cm−3,变异系数为10.94%。由此可知,木材基本密度的变异较为丰富,有较大的选择潜力。木材基本密度在产地间和产地内无性系间变异分别为总变异的22.88%和26.71%,产地内变异大于产地间,因此在充分利用产地间变异的同时,更应重视产地内优树的选择。木材基本密度在幼龄材、过渡龄材和成熟材的无性系均值分别为0.576、0.452和0.449 g·cm−3,幼龄材基本密度高于过渡龄材和成熟材,这可能是幼龄材松脂类浸提物含量较高的缘故。同时,发现木材基本密度在幼龄材、过渡龄材和成熟材的产地间和产地内皆存在显著(P<0.01)或极显著(P<0.001)差异,并且在幼龄材和过渡龄材、成熟材之间的差异也较显著(P<0.05),但过渡龄材和成熟材差异不显著(P>0.05)。而相对于过渡龄材和成熟材,木材基本密度在幼龄材的变化范围较大,变异系数亦较高,说明马尾松木材基本密度在早期具有较大的选择潜力。
纤维长度、纤维宽度和纤维长宽比在产地间和产地内均存在一定幅度的变异,变异系数从大到小依次为纤维长宽比、纤维长度、纤维宽度,均低于10%,表明这3个性状较为稳定。纤维长度和纤维宽度在产地间和产地内均存在显著(P<0.01)或极显著(P<0.001)差异。纤维长宽比在产地内差异明显,但在产地间差异不显著(P>0.05)。其中,纤维宽度的遗传变异主要来源于产地间,而纤维长度、纤维长宽比的遗传变异主要来源于产地内无性系间,表明纤维宽度通过产地间选择将会获得较大的遗传增益。相反,纤维长度和纤维长宽比通过产地内选择可取到较好的遗传改良效果。纤维长度、纤维宽度和纤维长宽比的无性系均值在幼龄材和成熟材差异均显著(P<0.05),且成熟材的纤维长度和纤维宽度高于幼龄材,而成熟材的纤维长宽比低于幼龄材。纤维长度、纤维宽度和纤维长宽比在幼龄材、过渡龄材和成熟材的变动幅度都较大,表明这3个纤维性状在不同生长期皆存在丰富的变异,具有较大的选择潜力。

表 1 马尾松无性系木材基本密度和纤维形态的方差分析及遗传参数估计Table 1 Result of ANOVA and genetic parameter estimates for wood basic density and fiber morphology of P.massoniana clones
2.2 马尾松无性系木材基本密度和纤维形态间的相关性
2.2.1 马尾松无性系的性状相关分析 由表2可知:木材基本密度和纤维长度、纤维长宽比具有极显著(P<0.001)的中等强度遗传负相关和显著(P<0.01)的弱度表型负相关,并且遗传相关系数均高于其表型相关系数。木材基本密度与纤维宽度呈表型和遗传的正相关,但相关不显著(P>0.05)。说明提高马尾松木材基本密度会降低纤维长度和纤维长宽比,但对纤维宽度没有明显的影响。纤维长度与纤维宽度、纤维长宽比呈极显著(P<0.001)的表型和遗传正相关。
2.2.2 马尾松无性系在幼龄—成熟期的相关分析 从表3可见:幼龄—成熟期的纤维长度、纤维宽度和纤维长宽比呈极显著(P<0.001)的遗传正相关,而纤维长度、纤维长宽比均呈极显著(P<0.001)的弱度表型正相关,纤维宽度在幼龄—成熟期表型相关不显著(P>0.05)。纤维形态在幼龄—成熟期遗传相关系数均大于0.66,幼龄—成熟期表型相关系数为0.138~0.293。遗传相关系数去掉了环境因素的影响,比较真实地反映2个性状间的相关关系,因此纤维长度、纤维宽度和纤维长宽比可作为材性进行早期选择。但这3个性状的表型相关小于其遗传相关,故早期选择时应考虑立地条件、气候等环境因素的影响。幼龄—成熟期马尾松木材基本密度的表型相关系数为0.135,相关系数较小且不显著(P>0.05),说明木材基本密度进行早期预测是不可靠的。
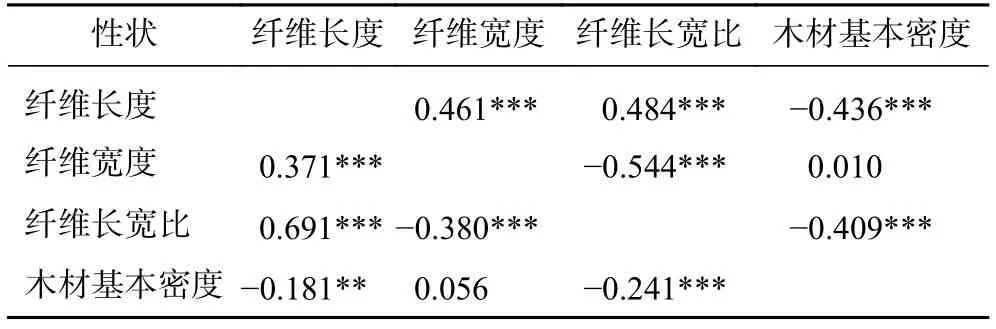
表 2 木材基本密度和纤维形态的遗传相关和表型相关Table 2 Genetic correlation and phenotypic correlation between wood basic density and fiber morphology

表 3 木材基本密度和纤维形态在幼龄—成熟期的相关系数Table 3 Correlation coefficients of wood basic density and fiber morphology between the juvenile wood and the mature wood
2.3 马尾松优良无性系的选择
在林木材性改良中,希望获得密度大且纤维长的优良材料,但只考虑纤维长度,会使单根纤维的成纸强度较大而纤维间结合力较差。纤维长宽比更能科学评价纤维特征与纸张性能的关系。因此,以木材基本密度和纤维长宽比大于群体平均值进行选择,共选出材性优良的马尾松无性系12个(表4)。马尾松优良无性系主要从贵州、浙江和江西3个主产地选出,而在江西产地中,能获得具有最优木材性状的无性系。即在所选择的优良无性系中,木材基本密度最高的无性系是6119,为0.531 g·cm−3;而纤维长宽比最大的无性系是6309,纤维长度为3.23 cm,纤维长宽比达107.81。

表 4 基于木材基本密度和纤维长宽比的优良无性系选择Table 4 Selection for superior clones based on wood basic density and fibre length-width ratio
3 讨论
木材基本密度是林木品质和材性遗传改良研究中的重要指标之一。ZOBEL等[16]指出:木材密度的微小变化(±0.04 g·cm−3)将致使每公顷干物质产量发生巨大变化。本研究表明:木材基本密度的均值为0.461 g·cm−3,在 0.350~0.636 g·cm−3内变异,与其他马尾松的研究基本相符[11,17−18]。其他研究表明:木材基本密度的平均值为0.383~0.491 g·cm−3。木材基本密度在产地间和产地内无性系水平差异显著,且在产地内无性系水平存在较大的遗传改良潜力,因此,在选优时进行产地内无性系的选择能取得显著效果。
纤维长度和纤维长宽比是衡量造纸原料质量优劣的重要指标,与成纸的物理性能有直接关系。对杨树Populus[19]、翠柏Calocedrusmacrolepis[20]等木材研究发现:纤维长度越长,长宽比越大,纸张的撕裂度、抗拉强度、耐破度和耐折度等也越强。马尾松木材纤维长度大于1.6 mm(长纤维),长宽比均值大于90,适宜作为造纸的优良原材料[21]。本研究中,纤维长度、纤维宽度和纤维长宽比均值分别为2.77 mm、28.76 μm、98.58,小于黄光霖等[22]对马尾松的测定结果。究其原因,后者通过显微镜观察来测定,受主观影响相对较大,如选择较长的纤维进行测定,而且考虑到工作量,通常纤维样本量不超过50根,而本研究中使用FS5纤维分析仪进行测定,测定样本量较大,多达几万根纤维,实验结果相对更精确。本研究发现:成熟材的纤维长度和宽度高于幼龄材,而成熟材的纤维长宽比低于幼龄材,与其他对马尾松纤维形态的变异研究结果不一致[23−24],可能是本次研究的取材部位与前人不同所致。本研究还表明:纤维形态的3个指标在产地间和产地内的无性系间皆存在显著(P<0.01)或极显著(P<0.001)差异,说明马尾松纤维性状在产地间和产地内的无性系间存在广泛变异,具有一定的选择基础和改良潜力。
林木生长周期较长,基因杂合度较高,育种进程十分缓慢。为缩短育种年限,早期选择作为筛选优良种源的有效途径而颇受重视。早期选择是根据幼龄期和成熟期的相关性来对林木成熟龄期的目标性状进行选择。幼龄材指髓心附近的树木生长初期形成的木质部,成熟材是指靠近树皮部分的木质部。金国庆等[25]对不同林龄的马尾松研究表明:胸径、树高和材积等生长性状在早期进行选择是可行的。本研究表明:马尾松纤维形态在幼龄—成熟期相关及其紧密,且主要受遗传因素控制。因此,进行纤维形态的早期选择是可行的。但木材基本密度和纤维长度、纤维长宽比呈显著负相关(P<0.01),因此在进行早期选择时,应同时兼顾木材基本密度表现。
4 结论
马尾松不同产地无性系的木材基本密度和纤维形态在产地间和产地内都存在显著差异。马尾松木材基本密度、纤维长度和纤维长宽比在产地内具有较大的改良潜力,而纤维宽度在产地间具较好的改良效果。纤维形态指标比较适合进行早期选择,但应兼顾木材基本密度的选择。本研究共选出12个材性优良的马尾松无性系,供生产造林。
