秦岭次生辽东栎林新生个体对种群空间格局的响应
2021-01-05吴润琴邹书珍康迪
吴润琴, 邹书珍, 康迪
西南野生动植物资源保护与利用教育部重点实验室,西华师范大学生命科学学院,四川 南充 637002
森林是陆地生态系统重要的组成部分,具有涵养水源、净化空气、保育土壤、调节气候等重要的生态服务功能[1-3]。而森林更新是森林提供可持续生态服务功能的前提条件。关于森林更新的影响因素已经有很多研究,国外De Lombaerde 等认为气候变化通过物理、化学和生物过程改变森林的能量分配、水文循环和大气成分等从而影响森林更新[4],国内闫兴富等在2011—2014 年间研究了光照、温度、密度及动物取食行为对辽东栎幼苗更新的影响[5-8],方坚等分别研究了油松和辽东栎混交林的种间和种内竞争对群落和种群动态的影响[9],Berger (2004)等人观测了加拿大西部森林超过50 年后报道,群落内部竞争产生的驱动力比外部气候因素更能推动森林产生动态变化[10]。但并列分析种群新生个体分别对胸径、密度、竞争等因素的响应的文章却较少。
种群更新对胸径、密度和竞争的响应通过新生个体与周围成树之间的相互作用体现,例如有文章研究发现成树对幼树的限制作用[11-13]。为了更加深入地了解种群内个体大小、密度和基于距离的竞争对幼苗更新的影响,本文以秦岭次生辽东栎种群为研究对象,以长期的连续观察代替时空互代法,设置了长达10 年(观测频次为5 年)的定位观测。采用插值法、密度函数法和基于距离的竞争指数法,对比分析了每5 年新生个体对周围成树胸径大小、密度和竞争格局的响应,为秦岭次生辽东栎林的恢复和近自然管理提供参考。
1 研究位置概况
秦岭山脉,呈东西走向,横贯中国中部,西起甘肃省临潭县北部的白石山,向东经天水南部的麦积山进入陕西。在陕西与河南交界处分为三支,长约1 600 km,为黄河支流渭河与长江支流嘉陵江、汉水的分水岭,海拔由东向西逐渐增高,主峰太白山3 771.2 m。秦岭南北气候呈差异性变化,有“南北植物荟萃、南北生物物种库”之称,以南属亚热带气候,植被以常绿阔叶林为主,以北属暖温带气候,植物以落叶阔叶树为主。辽东栎林占领了秦岭北坡海拔1 800~2 200 m 的广阔地带,多为纯林,伴有少量侧柏、杜梨、山杏等树种。
2 材料和方法
2.1 样地设置与数据采集
在秦岭太白山北坡辽东栎林的主要分布区内建立1 个固定观测样地,在样地中建立4 个28 m×28 m 重复样方,10 年间分别在第1、第5、第10 年对4 个样方进行3 次调查(下文统一定义第1 年、第5 年、第10 年为A、B、C)。每次每个样方内均进行每木调查,对所有辽东栎个体进行统一编号,并记录个体的水平坐标、胸径、平均冠幅等信息。从第5 年调查开始,将实际新增的、存活的,高度1 m 以上或基径1 cm 以上的个体定义为更新苗,记录其坐标和基径。
2.2 径级结构的分析
基于树木径级与龄级反应规律的一致性,采用立木径级结构代替年龄结构对3 次调查的种群结构进行分析[14]。参照高贤明等的方法[15]和根据实际情况将调查个体划分为7 个径级:第1 径级为1 cm<DBH<8 cm 的个体;2~6 径级以4 cm 为步长递增;第7 径级为DBH≥28 cm 的个体。根据选用的径级标准,通过Origin9.0 软件完成种群径级结构分析。
2.3 竞争指数的计算
竞争指数用于表示个体受到竞争的强度,反映了对象木所承受的来自周围其他个体的竞争压力,本研究依据的是与距离相关的Hegyi 竞争指数,该指数可以独立地表示处于连续动态发展的特定种群或群落中每个分布个体的竞争大小。本研究的竞争指数的计算方法由Hegyi 的竞争指数演变而来[16,17],其公式如下:
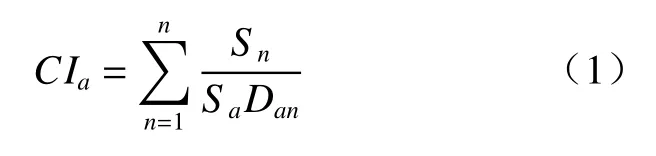
CI表示个体a的竞争指数;n表示目标个体a周围xm 以内的个体数(x为样地中辽东栎的平均冠幅);Sa和Sn分别表示目标个体和竞争个体的胸径大小;Dan表示目标个体和竞争个体的距离。4 个样方的2 个林龄过渡期(第1~5 年、第5~10 年)分别进行计算。
2.4 数据分析与软件
竞争指数计算通过R 软件完成;密度分析与插值分析采用ArcGis10.0 完成;绘图采用ArcGis10.0以及Origin9.0 完成[18-21]。
3 结果与分析
3.1 辽东栎次生林径级结构
结果表明,第1 年、5 年和10 年调查时,样地内辽东栎个体数分别为217 株、243 株和267 株,总体表现为增长,平均增长速率为25 株·5 年−1。为了进一步分析种群结构,绘制柱状图,如图1 所示,样地中中小径级(1~4 级,DBH<20 cm)个体较多,大径级(DBH≥20 cm)个体较少,种群结构为增长型。与第1 年相比,第5 年和第10 年时,种群内1 cm<DBH<8 cm 个体减少,而8 cm <DBH<24 cm 个体均有所增加。
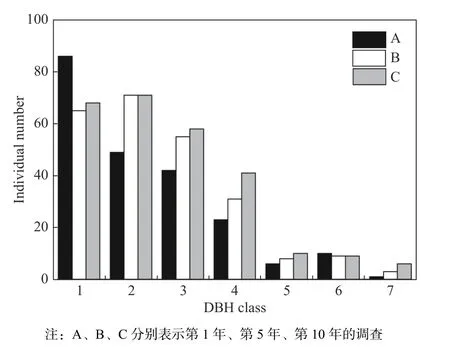
图 1 径级结构图Fig. 1 Diameter structure diagram
3.2 新生个体对胸径格局的响应
采用克里金插值模型对样地中的成树胸径进行二次分析,研究了样地立木胸径大小对新生个体分布的影响(见图2)。结果表明样方1、2、3 中第5 年和第10 年调查时新生个体均分布在平均胸径较小的区域,尤其在有较多成树的样方2 中,此规律更加明显。样方3 立木胸径较为均匀,新生个体的分布规律反而不明显。进一步绘制箱形图(见图3),结果表明:(1)样方2、3、4 中新生个体周围的成树平均胸径小于样方平均胸径,其中成树较多的样方2 最为明显;(2)样地内新生个体周围成树的平均胸径为10.376 cm,低于样地平均水平。

图 2 胸径克里金插值分析图(绿色三角形表示新生个体,下同)Fig. 2 Kriging interpolation analysis chart of the DBH (The green triangle represents a newborn individual, the same below)
3.3 新生个体对密度格局的响应
对样地第5 年和第10 年调查进行核密度分析,研究了样地立木密度对新生个体分布的影响(见图4)。结果表明:样方1、2、3、4 中第5 年和第10 年调查时新生个体均分布在密度较高的区域,尤其是在平均密度较高的样方1 中更加明显。进一步绘制箱型图(见图5),结果表明:(1)样方1、2、3、4 中新生个体周围成树的平均密度高于样方所有成树平均密度;(2)样地内所有新生个体周围的成树平均密度为0.126,高于样地的平均水平。

图 3 胸径分析箱图Fig. 3 Box plot of DBH
3.4 新生个体对竞争格局的响应
采用克里金插值模型对样地中的所有立木的竞争指数进行分析,研究了样地立木竞争大小对新生个体分布的影响(见图6),结果表明:样方1、2、3 第5 年和第10 年调查中大部分新生个体分布在中等竞争水平的区域,个别分布在竞争水平较高区域的边缘,样方4 中各个区域的竞争强度差异不大。进一步采用箱型图(见图7)分析结果表明:(1)样方1 中新生个体周围的成树竞争指数小于样方所有立木平均竞争指数;样方2 中第5 年时新生个体周围的成树竞争指数大于样方平均水平,第10 年时小于样方平均水平;样方3 新生个体周围的成树竞争指数小于样方平均水平;样方4 第5 年时新生个体周围的成树竞争指数明显大于样方平均水平,第10 年时接近样方平均水平。(2)样地内新生个体周围的成树竞争指数均值为16.425,低于样方1 和2 的平均水平,高于样方3 和4 的平均水平,没有明显规律。
4 讨论
林木的空间格局是生物和环境、生物与生物间长期相互作用的结果,是影响森林更新的重要因素[22]。森林空间格局的形成和变化除了受到外部环境影响外,还受到森林内立木大小、密度和其相互距离的影响,生态学家常依赖上述特征来研究森林更新、推断森林动态[23]。本文发现秦岭次生辽东栎种群小径级个体数量多而大径级个体数量较少,种群处于增长状态。但在第5 年与第10 年调查时,小径级个体数量较第1 年时少,其更新潜力有下降趋势。
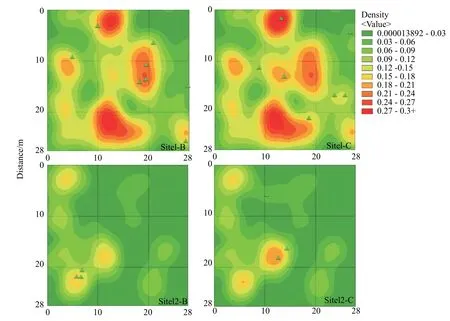
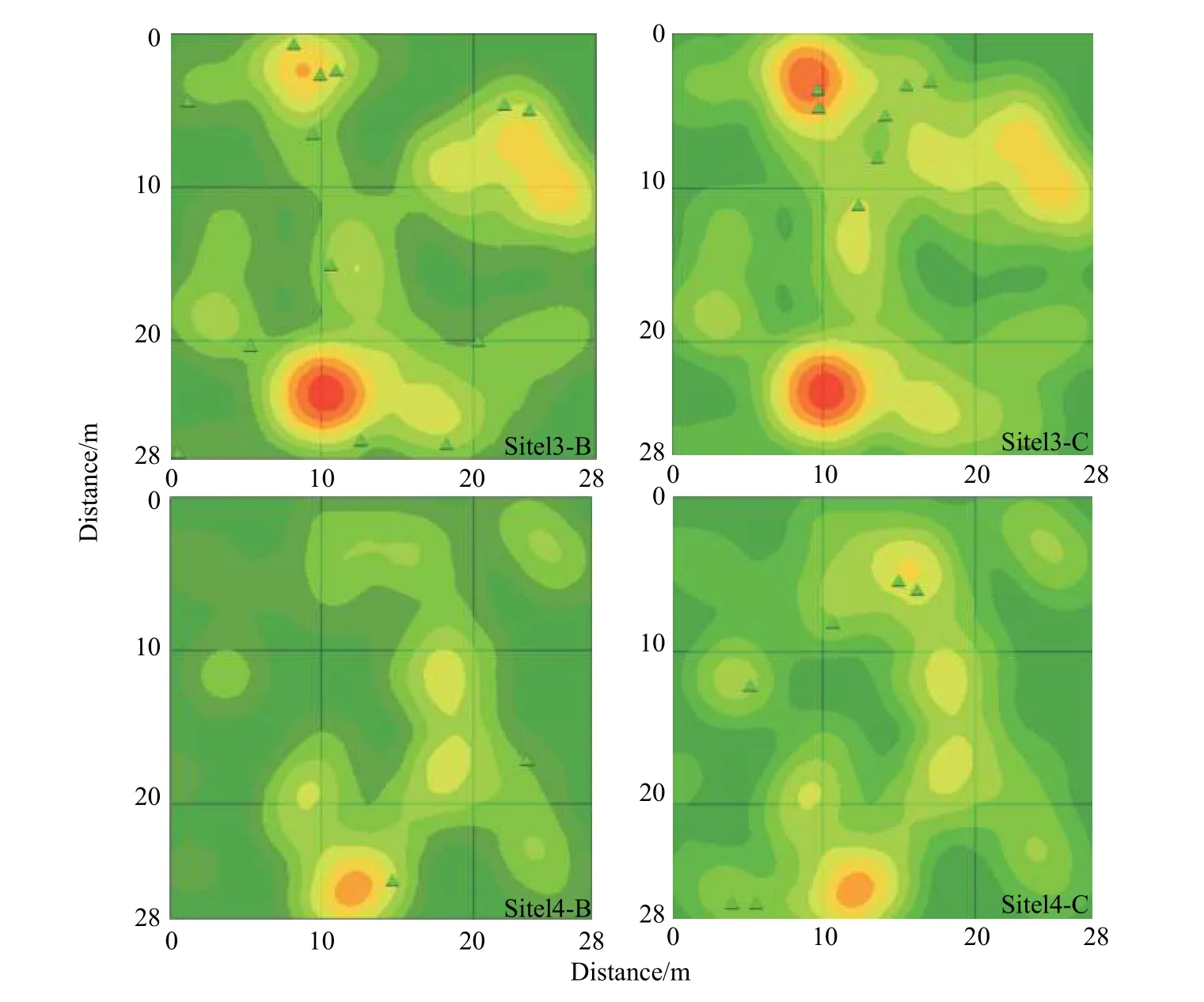
图 4 核密度分析图Fig. 4 Nuclear density analysis chart

图 5 密度分析箱图Fig. 5 Box diagram of density
本文发现秦岭次生辽东栎林种群新生个体的分布受区域种群胸径和密度的影响较为明显,即更新个体容易在周围成树平均胸径较小和平均密度较高的位置分布。辽东栎幼苗移栽过程中,增加密度有助于根系的生长从而提高成活率[24,25]。辽东栎虽属阳性树种,但幼苗却偏好更有利于其自然萌发的庇荫条件,遮阴促进辽东栎幼苗活力指数的增加,促进幼苗苗高和叶面积的生长[26],周围成树密度较高正好为新生个体提供一定的遮阴条件。辽东栎幼树在小尺度趋向于聚集分布[27],除了辽东栎种子的外形和传播特点的原因外,可能还因为在一定空间内呈聚集分布形成遮阴环境,提高幼苗存活率。王巍等(2000)研究发现辽东栎实生苗的密度会随幼苗年龄增大逐渐降低,而萌生更新多基株数的生物学特性却会显著提高局域种群密度[28],辽东栎自然更新过程中以萌生苗为主,这可能是新生个体周围成树平均胸径小而密度高的原因之一。此外,动物的搬运和取食作用也会对辽东栎新生个体的萌发和分布产生影响,有研究表明辽东栎种子54.5%被啮齿动物搬运,而且被搬运后以土壤埋藏为主要贮藏方式[29-31]。但种子被动物搬运后有可能分散贮藏也可能集中贮藏在洞穴或者灌木的根部[32,33],这个过程比较复杂,难以人为控制,但可以通过适度的人工管理,在利于新生个体分布的林分进行人工补苗,使种群更新良好。
本文发现次生辽东栎林种群新生个体周围成树的竞争强度差异较大,无明显规律。其他文章中认为中度的、适当的竞争有利于辽东栎的更新[34],随着个体胸径的增大竞争指数减小,相反胸径小,竞争指数大,自身竞争能力弱,对周围更新苗的压力就小[35,36],因此新生个体多数分布在周围成树胸径较小的区域。
在种群发展的不同阶段或是同一阶段的不同区域内,胸径、密度和竞争是动态变化的。但较之于竞争强度,密度和胸径对辽东栎新生个体分布的影响更为明显,这两个因素也更容易测算和人为调控。因此,在森林的近自然更新中,辅以适度的人工管理,充分考虑种群胸径和密度空间格局的异质性,以达到更好的经营管理效果。
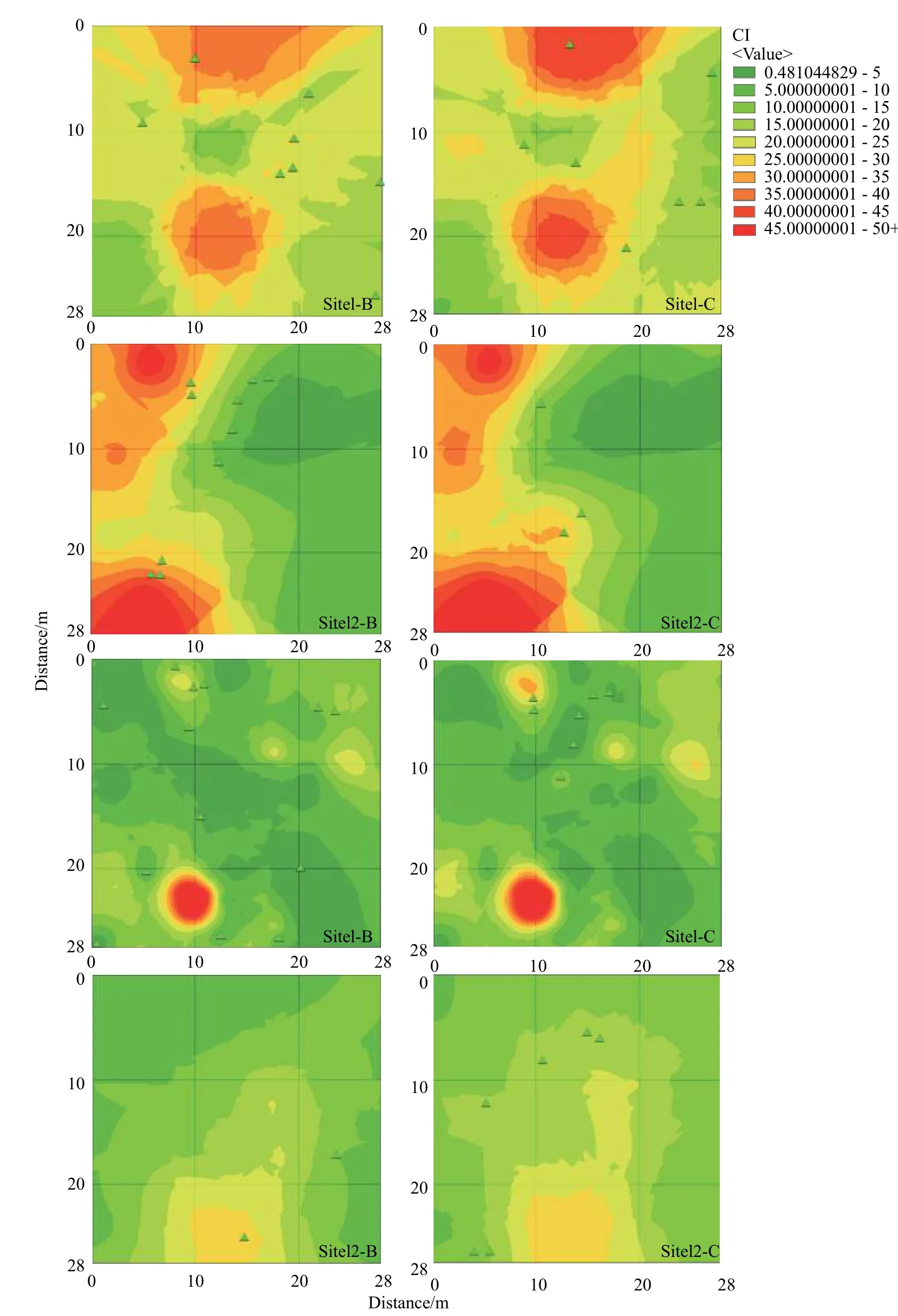
图 6 竞争指数克里金插值分析Fig. 6 Kriging interpolation analysis of competitive index

图 7 竞争指数箱图Fig. 7 Box diagram of competition index
