Description of a New Species of the Asian Newt Genus Tylototriton sensu lato(Amphibia:Urodela:Salamandridae) from Southwest China
2020-12-30ShizeLIGangWEIYanlinCHENGBaoweiZHANGandBinWANG
Shize LI,Gang WEI,Yanlin CHENG,Baowei ZHANG and Bin WANG*
1 Department of Food Science and Engineering,Moutai University,Renhuai 564500,Guizhou,China
2 CAS Key Laboratory of Mountain Ecological Restoration and Bioresource Utilization & Ecological Restoration Biodiversity Conservation Key Laboratory of Sichuan Province,Chengdu Institute of Biology,Chinese Academy of Sciences,Chengdu 610041,Sichuan,China
3 Biodiversity Conservation Key Laboratory,Guiyang College,Guiyang 550002,Guizhou,China
4 College of Life Science,Anhui University,Hefei 230039,Anhui,China
Abstract The newt genus Tylototriton sensu lato is widely distributed in Eastern,Southeastern and Southern Asia.Previous studies indicated that there still has been several cryptic species in the group.Here,we describe a new species of the genus from Guizhou Province,China.Phylogenetic analyses based on mitochondrial DNA supported that the new species was resolved as an independent clade nested into the Tylototriton sensu lato clade.On morphology,the new species could be distinguished from its congeners by a combination of the following characters:large body size (SVL 76.8-85.2 mm in male and 76.3-87.4 mm in female); head longer than wide; snout truncate in dorsal view; tail length longer than the snout-vent length in males; the distal digit ends,ventral digits,peripheral area of cloaca and the tail’s lower margin are orange; relative length of toes III > IV > II > I > V; the distal tips of the limbs greatly overlapping when the fore and hind limbs are pressed along the trunk; fingertips reach the level beyond the snout when the forelimbs are stretched forward.
Keywords China,molecular phylogenetic analysis,new species,newts,taxonomy
1.Introduction
The Asian newt genusTylototritonsensu latoAnderson,1871 is widely distributed from the eastern Himalayas,Indochina,to central and southern China (Fei and Ye,2016; Frost,2020).The taxonomic assignments of the group have been much controversial.Feiet al.(2005) divided the genus into two species groups,i.e.theT.verrucosusAnderson,1871 species group and theT.asperrimusUnterstein,1930 species group.Dubois and Raffaëlli (2009) upgraded the two groups as two subgeneraTylototritonandYaotriton.Feiet al.(2012)further classified all members ofT.sensu latointo three genera,T.sensu stricto,YaotritonandLiangshantriton(a new genus containing one speciesL.taliangensisLiu,1950).Feiet al.(2016) further classified the members ofT.sensu strictodefined by Fei and Ye (2016) into two subgenera,TylototritonandQianotriton(a new subgenus).Whatever,T.sensu latowas strongly resolved to be a monophyletic group by most molecular phylogenetic studies (e.g.,Nishikawaet al.,2013a,2013b; Wanget al.,2018).
In currently known 25 species ofT.sensu lato,17 species have been described in the last decade,and even ten species have been described in the past five years (Frost,2020).This leaded to the underrepresented systematic arrangements in previous studies especially in the species group with wide distributional ranges.For example,systematic profiles of theT.asperrimusspecies group have been continually changed,and the distributional range ofT.asperrimushas been gradually defined to be more narrowed,because many populations ever identified asT.asperrimuswere found to be separated species (e.g.,T.anhuiensisQian,Sun,Li,Guo,Pan,Kang,Wang,Jiang,Wu,and Zhang,2017,T.broadoridgusShen,Jiang,and Mo,2012,T.dabienicusChen,Wang,and Tao,2010,T.liuyangensisYang,Jiang,Shen,and Fei,2014,andT.wenxianensisFei,Ye,and Yang,1984).More remarkably,many new species were clustered into one apart clade (corresponding to the “T.wenxianensis” clade as Clade V in Wanget al.2018),distinctly from the true “T.asperrimus” clade (corresponding to Clade IV in Wanget al.,2018).In Wanget al.(2018),the “T.wenxianensis” clade containedT.wenxianensis,T.dabienicus,T.broadoridgus,T.liuyangensis,T.lizhenchangiand three putative cryptic species.The three putative cryptic speciesoccur from Dabie Mountains of Anhui Province,Wufeng County of Hubei Province and Libo County of Guizhou Province,respectively.The population from Dabie Mountains was already described as a new speciesT.anhuiensis(Qianet al.,2017).But the other two have not been investigated in detail and furtherly described.
In 2018,we collected some specimens ofT.sensu latofrom Maolan National Nature Reserve in Libo County,Guizhou Province,China.Phylogenetic analyses and morphological comparisons indicated that the population was distinctly different from its congeners.Herein we describe it as a new species.
2.Materials and Methods
2.1.SpecimenA total of nine specimens of the undescribed species including two females,six males and one larvae (for voucher information see Tables 1 and S1)were collected from Maolan National Nature Reserve in Libo County,Guizhou Province,China (Figure 1).In the field,after taking photographs,the newts were euthanized using isoflurane,and then the specimens were fixed in 10% buffered formalin.Tissue samples were taken and preserved separately in 95% ethanol prior to fixation.The specimens were deposited in Chengdu Institute of Biology (CIB),Chinese Academy of Sciences (CAS).The Animal Care and Use Committee of CIB,CAS provided full approval for this research (Number:CIB2013041103).Field experiments were approved by the Management Office of Maolan National Nature Reserve,Guizhou Province,China (Project number:201102034).
2.2.Molecular data and phylogenetic analysesTotal DNA was extracted from tissue samples of five specimens of the undescribed species using a standard proteinase phenol-chloroform protocol (Sambrooket al.,1989).The mitochondrial 16S rRNA and NADH dehydrogenase subunit 2 (ND2) genes were amplified,the primers P7 (5’-CGCCTGTTTACCAAAAACAT-3’),P8 (5’-CCGGTCTGAACTCAGATCACGT-3’) and SL-1(5’-ATAGAGGTTCAAACCCTCTC-3’),SL-2 (5’-TTAA AGTGTCTGGGTTGCATTCAG-3’) were used for 16S rRNA and ND2 following Simonet al.(1994) and Shenet al.(2012),respectively.They were amplified under the following conditions:an initial denaturing step at 95°C for 4 min; 36 cycles of denaturing at 95 °C for 30 s,annealing at 52 °C (for 16S)/58 °C (for ND2) for 40 s and extending at 72 °C for 70 s,and a final extending step of 72 °C for 10 min.Sequencing was conducted using an ABI3730 automated DNA sequencer in Shanghai DNA BioTechnologies Co.,Ltd.(Shanghai,China).All new sequences were deposited in GenBank (for accession numbers see Table 1).
For phylogenetic analyses,the available sequence data for all species of the genusTylototritonwere downloaded from GenBank (for accession numbers see Table 1).Corresponding sequences of onePachytriton labiatus,onePleurodeles waltland oneEchinotriton chinhaiensiswere downloaded (for GenBank accession numbers see Table 1),andPachytriton labiatuswas used as outgroup according to the previous study (Qianet al.,2017; Wanget al.,2018).
Sequences were assembled and aligned using the ClustalW module in BioEdit v.7.0.9.0 (Hall,1999) with default settings.Alignments were checked by eye and revised manually if necessary.To avoid bias in alignments,the regions of defined sequence conservation were extracted from the length-variable 16S gene fragments using GBLOCKS v.0.91.b (Castresana,2000) with default settings.Non-sequenced fragments were defined as missing loci.
Phylogenetic relationships were reconstructed based on the mitochondrial gene concatenated dataset.Phylogenetic analyses were conducted using maximum likelihood (ML) and Bayesian Inference (BI) methods,implemented in PhyML v.3.0 (Guindonet al.,2010) and MrBayes v.3.12 (Ronquist and Huelsenbeck,2003),respectively.To avoid under-or over-parameterization(Lemmon and Moriarty,2004; McGuireet al.,2007),the best partition scheme and the best evolutionary model for each partition were chosen for the phylogenetic analyses using PARTITIONFINDER v.1.1.1 (Robertet al.,2012).In this analysis,16S gene and each coding position of ND2 gene were defined and Bayesian Inference Criteria was used.As a result,the analysis suggested that the best partition scheme is 16S gene+(first+second) coding positions of ND2 gene+the third coding positions of ND2 gene,and selected TrN+I+G model as the best model for each partition.For the ML tree,branchsupports were drawn from 10,000 nonparametric bootstrap replicates.In BI analyses,the parameters for each partition were unlinked,and branch lengths were allowed to vary proportionately across partitions.Two runs each with four Markov chains were simultaneously run for 50 million generations with sampling every 1,000 generations.The first 25% trees were removed as the“burn-in” followed by calculations of Bayesian posterior probabilities and the 50% majority-rule consensus of the post burn-in trees sampled at stationarity.Genetic distance betweenTylototritonspecies was calculated with uncorrectedp-distance model on ND2 gene using MEGA v.6.06 (Tamuraet al.,2013).
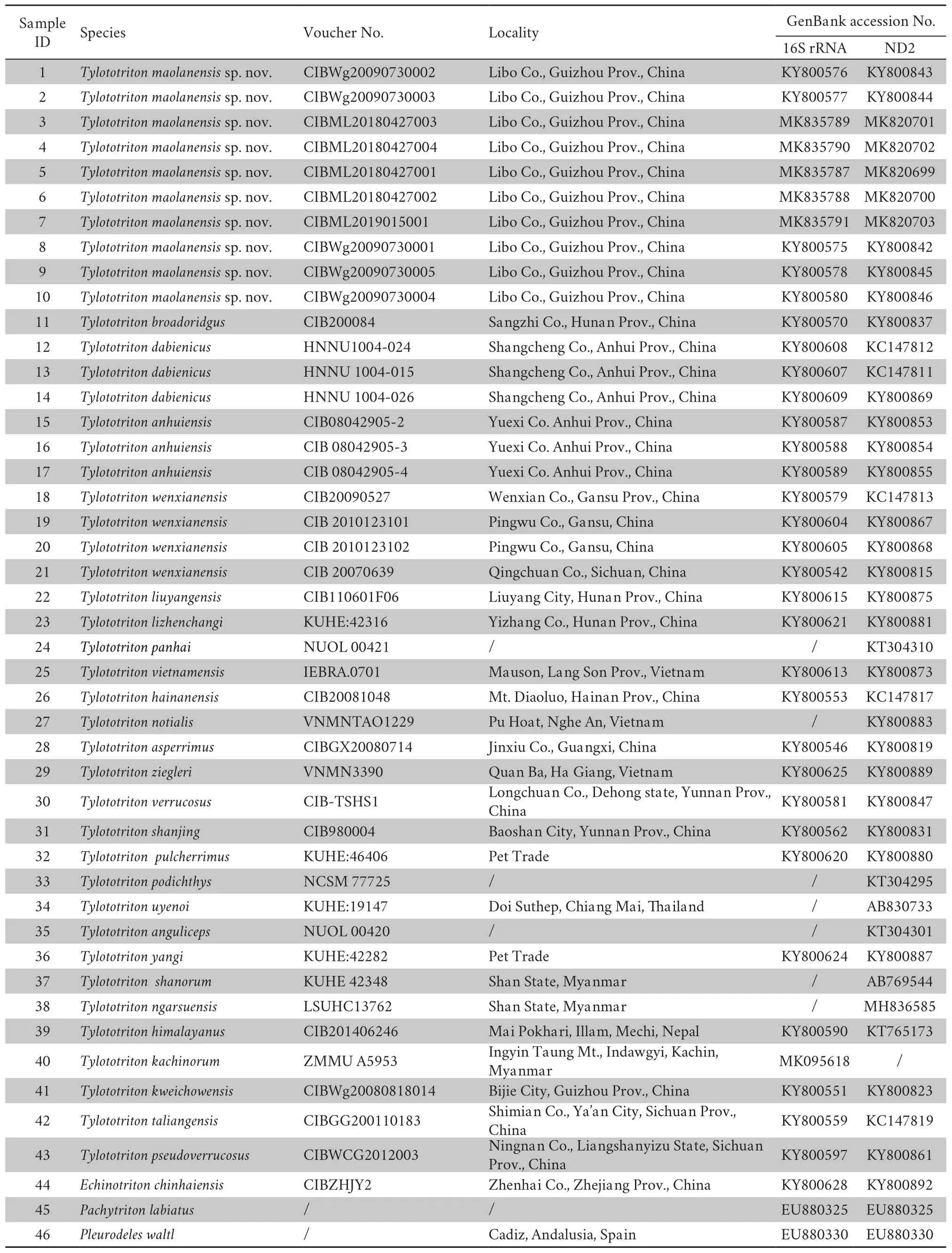
Table 1 Information of samples used in the molecular analyses in this study.

Figure 1 The type locality of Tylototriton maolanensis sp.nov.,Maolan National Nature Reserve,Libo County,Guizhou Province,China.
2.3.Morphological comparisonEight specimens of the undescribed species,sevenT.wenxianensisfrom two localities,threeT.broadoridgusfrom its type locality(Sangzhi County,Hunan Province,China) and fourT.dabienicusfrom its type locality (Shangcheng County,Henan Province,China) were examined and measured(Table S1).The terminology and methods followed Fei(1999).Measurements ofT.anhuiensiswere retrieved from Qianet al.(2017).
Measurements were taken with a dial caliper to 0.1 mm.Twenty morphometric characters of adult specimens were measured:total length (TOL,distance from tip of snout to tip of tail); snout-vent length (SVL,distance from tip of snout to posterior edge of vent); head length (HDL,distance from jugular fold to snout tip); head width (HDW,maximum head width); snout length (SL,distance from tip of snout to the anterior corner of eye); eye diameter(ED,distance from the anterior corner to the posterior corner of the eye); interorbital distance (IOD,minimum distance between the eyes); upper eyelid width (UEW,greatest width of the upper eyelid margins measured perpendicular to the anterior-posterior axis); internasal distance (IND,minimum distance between the external nares); IFE (distance between anterior corner of eyes); IAE(distance between posterior corner of eyes); trunk length(TRL,from gular fold of throat to anterior tip of vent);tail length (TAL,from anterior tip of cloaca to tip of tail);tail height (TH,maximum tail height); length of lower arm(LLA,distance from elbow to tip of wrist); hand length(HL,wrist from elbow to longest finger); the third finger length (FIIIL,distance from base to tip of finger III); thigh length (TLH,distance from groin to knee); tibia length(TL,distance from knee to tarsus); and the third toe length(TIIIL,distance from base to tip of toe III).
Only one larvae of the undescribed species (voucher number:CIBML2019015001) was collected a nd measured.Ten morphometric characters of the larvae were measured:total length (TOL,distance from tip of snout to tip of tail); snout-vent length (SVL,distance from tip of snout to posterior edge of vent); head length (HDL,distance from jugular fold to snout tip); head width (HDW,maximum head width); snout length (SL,distance from tip of snout to the anterior corner of eye); eye diameter (ED,distance from the anterior corner to the posterior corner of the eye); interorbital distance (IOD,minimum distance between the eyes); internasal distance (IND,minimum distance between the external nares); trunk length (TRL,from gular fold of throat to anterior tip of vent); tail length (TAL,from anterior tip of cloaca to tip of tail); tail height (TH,maximum tail height); length of lower arm and hand (LAL,distance from the elbow to the distal end of the longest finger); and the length of hind leg (HLL,from groin to tip of the longest toe).
For morphometric analyses,in order to reduce the impact of allometry,the correct value from the ratio of each character to TOL was calculated and then was logtransformed.Mann-WhitneyUtests were conducted to test the significance of differences on morphometric characters between the undescribed species,T.anhuiensis,T.broadoridgus,T.dabienicusandT.wenxianensis.The significance level was set at 0.05.Furthermore,principal component analyses (PCA) were conducted to highlight whether the different species were sepa ra ted in morphometric space.Due to only the measurements TOL,SVL,HDL,HDW,TRL,SL,ED,IOD,TAL and TH of maleT.anhuiensisbeing available from Qianet al.(2017),the morphometric analyses were conducted only based on these ten morphometric characters for PCA in male group.The analyses were carried out in R (R Development Core Team).
The undescribed species was clustered into theT.wenxianensisspecies group (Wang et al.2018),and it was compared with the members of this groupon morphology.Comparative morphological data were obtained from literature forT.anhuiensis(Qianet al.,2017),T.asperrimus(Unterstein,1930),T.broadoridgus(Shenet al.,2012),T.dabienicus(Chenet al.,2010),T.liuyangensis(Yanget al.,2014),T.lizhenchangi(Houet al.,2012),andT.wenxianensis(Feiet al.,1984).
2.4.Skull scanningSkulls of three male specimens(voucher numbers:CIBML20180427001,CIBML20180427002 and CIBML20180427004) and two female specimens (voucher numbers:CIBML20180427003 and CIBML20180427006) of the undescribed species were scanned in the high-resolution X-ray scanner (Quantum GX micro-CT Imaging System; PerkinElmer,Boston,MA,USA) in CIB,CAS.The specimens were scanned along the coronal axis at an image resolution of 2000 × 2000.Segmentation and three-dimensional reconstruction of the CT images were made using VG57 Studio Max 2.2(Volume Graphics,Heidelberg,Germany).
3.Results
3.1.Phylogenetic analysesAligned sequence matrix of 16S+ND2 genes contained 1540 bps.ML and BI analyses resulted in essentially identical topologies (Figure 2).The genusT.sensu latowas strongly supported as a monophyly.All samples of the undescribed species occurring from Maolan National Nature Reserve of Guizhou Province,China were strongly nested into one clade,which was sister toT.broadoridgus,and close toT.wenxianensis,T.dabienicusandT.anhuiensis.
Genetic distance on ND2 gene with uncorrectedp-distance model between specimens of the undescribed species was less than 0.5%,much lower than interspecific genetic distances of the genusT.sensu lato(1.2%-14.3%;Table 2).Genetic distances between the undescribed species and its closely related species,T.dabienicus,T.anhuiensis,T.broadoridgusandT.wenxianensiswere 2.8%-3.9% (Table 2),being higher than that between some substantial sister species,for example,T.shanjingvs.T.verrucosus(1.2%),T.shanjingvs.T.pulcherrimus(2.5%),T.verrucosusvs.T.pulcherrimus(2.1%),andT.taliangensisvs.T.pseudoverrucosus(2.7%).Genetic distance on 16S gene and 16S+ND2 data between the new species andT.anhuiensis,T.broadoridgus,T.dabienicusandT.wenxianensisare also in the same level,i.e.,0.8%-1.6% on 16S gene and 2.2%-2.9% on 16S+ND2 data (Table S2).
3.2.Morphological comparisonsIn PCA,for males,the total variation of the first two principal components was 75.3%,and for females was 52.3%.Both in males and females on the two-dimensional plots of PC1 vs.PC2,the undescribed species could be separated from its closely related speciesT.anhuiensis,T.broadoridgus,T.dabienicusandT.wenxiansis(Figure 3).The results of Mann-Whitney U tests indicated that in males,the undescribed species was significantly different fromT.anhuiensis,T.broadoridgus,T.dabienicusandT.wenxiansison a series of morphometric characters (allP-values < 0.05;Table 3).More detailed descriptions of the results from morphological comparisons between the undescribed species and its congeners were presented in the following sections for describing the new species.
3.3.Description of the new species
Tylototriton maolanensis sp.nov.
Holotype.Figures 4 A,5A,5B,5I,5K,5L,6.Male,CIBML 20180427001,collected on 27 April 2018 by Shize LI from Maolan National Nature Reserve (25.307194° N,107.948764° E,ca.737 m a.s.l.) in Libo County,Guizhou Province,China.
Paratype.Five adult males (CIBML20180427002,CIBML20180427004,CIBML20180427005,CIBML20180427007 and CIBML20180427008),two females (CIBML20180427003 and CIBML20180427006)and one larva (CIBML2019015001) were collected on 27 April 2018 by Shize Li together with the holotype.
Diagnosis.T.maolanensissp.nov.is assigned to the genusT.sensu latobased upon molecular phylogenetic analyses and the following morphological characters:presence of dorsal granules,dorsolateral bony ridges on the head,knob-like warts on dorsolateral body,and the absence of a quadrate spine.
T.maolanensissp.nov.could be distinguished from its congeners by a combination of the following characters:(1) large body size (SVL 76.8-85.2 mm in male and 76.3-87.4 mm in female); (2) head length longer than wide; (3) snout truncate in dorsal view; (4) the tail length longer than the snout-vent length in males; (5) the distal digit ends,ventral digits,peripheral area of cloaca and the tail’s lower margin orange; (6) relative length of toes III > IV > II > I > V; (7) the distal tips of the limbs greatly overlapping when the fore and hind limbs being pressed along the trunk; (8) fingertips reaching to the level beyond the snout while the forelimbs being stretched forward.


Figure 2 Maximum likelihood (ML) tree reconstructed based on the mitochondrial 16S and ND2 gene sequences.Bootstrap supports (BS)/Bayesian posterior probabilities (BPP) were labeled beside nodes.Information of samples 1-46 refer to Table 1.
Description of holotype.Measurements in mm.Figures 4 A,5 A,5B,5I,5K,5L.SVL 80.8; TOL 165.2; head longer than wide (HDL/HDW=1.12); head slightly concave on the top; snout is nearly square,extending beyond the lower lip; naris nearly snout; snout truncate in dorsal view; the supratemporal ridges on the head are long,steep and notable,extending from the dorsal region of the rostral side,and through the interior side of the upper eyelid to the occiput; a pair of short and protruding ridges on middle of supratemporal ridges; two triangular bony ridges on the occiput point laterally idge,distinctly being not connected with the dorsal ridge of body; two bony ridges on the dorsal head surface form a “V” shape and disconnected with the dorsal ridge of body; inter-nasal space is smaller than the inter-orbital space; eyes protrude from the dorsolateral portion of the head; the oral fissure is flat; the joint of jaw articulation lies posterior to the caudal margin of the eyes; fine teeth present on the edge of jaw,vomerine teeth long and prominent,form a “∧”shape which separated from each other at the tip of the“ ∧” shaped ; tongue oval and nearly entirely fixed at bottom but free at both lateral edges; neck is rounded and thick,with distinct neck groove.
Body is stout; two sides of the ridge are slightly concave.Each lateral corner of the dorsal ridge consists of 17 knob-like dorsal warts,and fine transverse striae present between every two knob-like dorsal warts.
Four limbs are relatively slender,and hind limbs are slightly longer than forelimbs; the distal tips of limbs greatly overlapping when fore and hind limbs are pressed along the trunk; fingertips are reaching to the level beyond the snout while the forelimbs are stretched forward;relative fingers length is III > II > IV > I,and relative toes length is III > IV > II > I > V; fingers and digits are with none webbed.
The tail length is longer than the snout-vent length;the tail is notably compressed laterally; the dorsal fin fold of the tail,starting from the tail base,distinctly thin and high; while the ventral fin fold of tail,starting from posterior to the cloaca,is thick and short; the tail height is greater than the width at the tail base,and the distal tail tip is rounded; the cloaca is long and narrow,and the cloacal region is slightly bulbous.
The skin is extremely rough,and body covered with tubercles and warts; the labial margin,distal limbs,ventral limbs and the ventral edge of the tail are smooth; the dorsal ridge,running along the middle of the dorsum and extending from the neck to the base of the tail,is rough and relatively narrow; each lateral corner of the dorsal ridge consists of a row of rough knob-like dorsal warts;on the lateral dorsum of the body,tubercles and warts are big,appearing to form lines that extend from the shoulder to the base of the tail; the ventral tubercles and warts are relatively flat and smaller.
Color in life.Figure 5 A,B,I,K,L.In life,the specimen black or blackish-brown,while the ventral color little lighter; the distal digit ends,ventral digits,the peripheral area of cloaca and the ventral edge of tail are orange; the orange region between the ventral edge of the tail and the peripheral area of the cloaca is connected.
Color in preservative.Figure 4 A.The specimen in preservative is black and the ventral color is blackishbrown.The orange coloration of the distal digit ends,ventral digits,the peripheral area of the cloaca and the ventral edge of the tail fades to lacte.
Variation.A large proportion of specimens were similar in morphology with the holotype,but some individuals are different.In female individuals the tail length is shorter than the snout-vent length; in some individuals the knob-like dorsal warts on lateral corner of the dorsal ridge are not black but with orange slightly,and in some individuals the knob-like dorsal warts on lateral corner of the dorsal ridge are 13,15,16 or 17 (Figures 5 C-5 F,5 J)
Secondary sexual characteristics.The female cloacal hole is short and its inner cloacal wall has no papilla(Figure 5 I).The male has papilla on its inner cloacal wall and its cloacal orifice long slit (Figure 5 J).

Figure 3 Plots of principal component analyses for Tylototriton maolanensis sp.nov.and its relatives.A,male.B,female.PC1,the first principal component;PC2,the second principal component.
Skull.The skull morphology of the five scanned specimens were almost identical,and thus,one representative(voucher number:CIBML20180427002) was presented(Figure 6) and described as following.Skull wider than long,equipped with numerous sculpture-shaped ridges dorsally.Nasal connects with one another along the midline,with the maxilla laterally,with the premaxilla anteriorly,and with the prefrontal and the frontal posteriorly.The nasal process is short and far away from the frontal.Both the septomaxilla and the lacrimal are absent.The frontal is obviously larger than the parietal.The squamosal is enlarged.The frontosqumosal arch is robust.The pterygoid is not directly meet with the maxilla.The maxilla is long,its anterior margin contacts with the premaxilla,and its posterior end contacts with the quadrate posteriorly.The premaxilla and the vomer form a fontanelle.The vomer is large and gradually narrowed from anteriorly to posteriorly.The vomerine teeth series are arranged in inverted V shape.The parasphenoid wedges between the vomers.
Larva.A larva was collected from the same pone of holotype on 27 April,2018 (Figure 7).Measurements in mm.TOL 55.2,SVL 31.8,HDL14.2,HDW7.6,TRL23.3,LAL7.4,HLL10.2,ED2.1,TAL23.8,TH4.3,IND2.4,IOD3.1,and SL3.2.Body slender.The forelimbs and hindlimbs clearly fully grown up.Head longer than wide,eyes relatively large on the flank and three pairs of external gills having obtuse anterior ends,while the body and tail being laterally compressed.Skin smooth,and except the ventral side of the tail,fingers and toes saffron yellow,body black-brown.Costal grooves for muscles of larvae distinctly.The dorsal fin fold comparatively tall and rises from the region between the posterior head and anterior body.The short and thick ventral fin fold of the tail extends from the cloaca to the end of the tail.
Morphological comparisons.T.maolanensissp.nov.had been identified asT.asperrimus(Wuet al.,1986; Feiet al.,2012; Guet al.,2012),but molecular phylogenetic analyses revealed that this new species was genetically closer toT.anhuiensis,T.broadoridgus,T.dabienicus,T.wenxianensis,T.liuyangensisand T.lizhenchangi.The new species can be distinguished from these closely-related species by a series of morphological characters (Figure 4; Table 3) as follows.
T.maolanensissp.nov.differs fromT.asperrimusby tail length longer than snout-vent length in males (vs.tail length shorter than snout-vent length in males); the distal tip of limbs greatly overlapping when the fore and hind limbs pressed along the trunk (vs.slightly overlapping,meeting or not meeting); fingertips reaching to the level beyond the snout while the forelimbs stretched forward(vs.just reaching to the nostril or eye).
T.maolanensissp.nov.differs fromT.anhuiensisby tail length longer than snout-vent length in males (vs.snoutvent length longer than tail length); the relative toes length I > V (vs.V > I); the distal tips of the limbs greatly overlapping when the fore and hind limbs pressed along the trunk (vs.slightly overlapping); having significantly longer TOL in males; having significantly higher ratios of ED,SVL,TRL,SL,TAL and TH to TOL in males; the transverse striae present between every two knob-like dorsal warts (vs.every two knob-like dorsal warts closely arranged); dorsolateral bony ridge on head steep,long,narrow and prominent (vs.dorsolateral bony ridge on head slightly prominent,gentle and wide); snout truncate in dorsal view (vs.snout obtusely rounded in dorsal view);and having long and strong vomerine tooth (vs.vomerine teeth short).

Figure 4 Photos of the holotype specimen of Tylototriton maolanensis sp.nov.and topotype specimens of its closely related species.A,holotype specimen CIBML20180427001 of Tylototriton maolanensis sp.nov.B,topotype specimen AHU-16-EE-001 of T.anhuiensis. C,topotype specimen CIBSZ20120620026 of T.broadoridgus. D,topotype specimen CIB8042905 of T.dabienicus.E,specimen CIB200811CD03 of T.wenxianensis.A1-E1,dorsal view of whole body.A2-E2,dorsal view of head.A3-E3,view of oral cavity.
T.maolanensissp.nov.differs fromT.dabienicusby tail length longer than snout-vent length in males (vs.snoutvent length longer than tail length); the relative toes length I > V (vs.V > I); the distal tips of the limbs greatly overlapping when the fore and hind limbs pressed along the trunk (vs.slightly meeting or not meeting); fingertips reaching to the level beyond the snout while the forelimbs stretched forward (vs.reaching to the eye); having significantly higher ratios of ED,TLH,TL and TAL to TOL in males; by dorsolateral bony ridge on head steep,long,narrow and prominent (vs.prominent but gentle and wild); by snout truncate in dorsal view (vs.snout obtusely rounded in dorsal view); by the bony ridges on the head extending from the dorsal region of the rostral side,and through the interior side of the upper eyelid to the occiput(vs.extending from the dorsal region of the rostral side to the posterior of orbit); by having a bigger angle formed by ends of vomerine teeth (vs.the angle smaller); by the separated tip of “∧” shaped vomerine teeth (vs.tip of “∧”shaped vomerine teeth connected).

Figure 5 Photos showing color variation in Tylototriton maolanensis sp.nov.in life.A and B,dorsal and ventral view of the holotype CIBML20180427001.C and D,dorsal and ventral view of the female paratype CIBML20180427003.E-H,dorsolateral view of the female paratype CIBML20180427006,and male paratypes CIBML20180427002,CIBML20180427004,and CIBML20180427008.I and J,cloaca of the holotype CIBML20180427001 and paratype CIBML20180427003.K and L,the ventral view of the hand and foot of the holotype CIBML20180427001.
T.maolanensissp.nov.differs fromT.wenxianensisby having a longer body length in males (minimum total length > 151 mm vs.maximum total length < 134 mm in the latter); head length longer than wide (vs.length equal to wide); tail length longer than snout-vent length in males (vs.snout-vent length longer than tail length); the relative toes length I > V (vs.V > I); the distal tips of the limbs greatly overlapping when the fore and hind limbs pressed along the trunk (vs.slightly meeting or slightly overlapping); fingertips reaching to the level beyond the snout while the forelimbs stretched forward (vs.reaching to the nostril); having significantly higher ratios of TOL and having significantly lower ratios of HDL,HDW,TRL,SL,ED,IFE,IAE,IOD,IND,HL,TLH,TL and TAL to TOT in males; by snout truncate in dorsal view (vs.snout obtusely rounded in dorsal view); by dorsolateral bony ridge on head steep,long,narrow and prominent(vs.prominent but gentle and wild); by the separated tip of “ ∧” shaped vomerine teeth (vs.tip of “ ∧” shaped vomerine teeth connected).
T.maolanensissp.nov.differs fromT.broadoridgusby having a longer body length in males (minimum total length > 151 mm vs.maximum total length < 141 mm in the latter); tail length longer than snout-vent length in males (vs.snout-vent length longer than tail length); the relative toes length I > V (vs.V > I); the distal tips of the limbs greatly overlapping when the fore and hind limbs pressed along the trunk (vs.slightly overlapping or just meeting); having significantly higher ratios of TOL,and significantly higher ratios of SVL,HDL,HDW,TRL,ED,IFE,IAE,IOD,LLA,TLH,TAL and TH to TOL; by dorsolateral bony ridge on head steep,long,narrow and prominent (vs.prominent but gentle); by having a bigger angle formed by ends of vomerine teeth (vs.the angle smaller).
T.maolanensissp.nov.differs fromT.liuyangensisby having a longer body length in males (minimum total length > 151 mm vs.maximum total length < 147 mm in the latter),tail length longer than snout-vent length in males (vs.shorter),the relative toes lengths I > V (vs.V> I),the distal tip of limbs greatly overlapping when the fore and hind limbs pressed along the trunk (vs.slightly overlapping,meeting or not meeting),the distal tip of limbs greatly overlapping when the fore and hind limbs pressed along the trunk (vs.slightly overlapping,meeting or not meeting).
T.maolanensissp.nov.differs fromT.lizhenchangiby head length longer than wide (vs.wider than long or equal in the latter),the relative toes lengths I > V (vs.V > I).

Figure 6 Skull of the holotype specimen CIBML20180427001 of Tylototriton maolanensis sp.nov.A,dorsal view.B,ventral view.(1) premaxilla; (2) prefrontal; (3) quadrate; (4) ceratobranchial 1; (5) nasal; (6) maxilla; (7) frontal; (8) squamosal; (9) parietal; (10) ceratohyal; (11) prootic-exoccipital complex; (12)orbitosphenoid; (13) vomer; (14) parasphenoid; (15) prearticular-angular; (16) pterygoid; (17) hypobranchial 1.

Figure 7 Photos of larva CIBML2019015001 of Tylototriton maolanensis sp.nov.in life.A,dorsal view.B,lateral view.C,ventral view.
Ecology.T.maolanensissp.nov.is currently known from Maolan National Nature Reserve (25.155°-25.347° N,107.869°-108.761° E) in Libo County,Guizhou Province,China.It inhabited in the puddles on a hillside or at the foot of a hill (Figure 8),at elevations of about 737 m a.s.l.Two sympatric amphibian species,Microhyla heymonsiVogt,1911 andHylarana latouchiiBoulenger,1920,were found in the type locality.
Etymology.The specific namemaolanensisis a Latinize toponymic adjective that refers to Maolan National Nature Reserve,Libo County,Guizhou Province,China,where the new species was firstly found.For the common name,we suggest Maolan Knobby Newt (English) and Mao Lan You Yuan (茂兰疣螈,Chinese).
4.Discussion
Most cryptic congeners in the genusT.sensu latoare difficult to be distinguished from each other due to the superficial similarities in morphology.This resulted in considerable challenges in field identification,which in turn cause ambiguities in taxonomy arrangement (Fei and Ye,2016; Feiet al.,2006,2012).T.maolanensissp.nov.had been initially identified asT.asperrimus(Wuet al.,1986; Guet al.,2012; Feiet al.,2012).In this study,phylogenetic analyses suggested that the new species was separated from its congeners.Genetic distance between the new species and its closely-related species(T.dabienicus,T.anhuiensisandT.broadoridgus) were much higher than that between many sister species inT.sensu lato.Indeed,there were some subtle morphological differences between the new species and its congeners with comparisons.Hence,all evidences supported thatT.maolanensissp.nov.is a valid species.Wang et al.(2018)indicated that the distributional range ofT.wenxianensisspecies group (containingT.anhuiensis,T.broadoridgus,T.dabienicus,T.wenxianensis,T.liuyangensis,T.lizhenchangiandT.maolanensissp.nov.) was basically in the north of Nanling Mountain series,and theT.asperrimusspecies group was distributed in the south of Nanling Mountain series.Maolan National Nature Reserve is located in the boundary of them.However,in our surveys in recent many years in Maolan National Nature Reserve,we did not find individuals ofT.asperrimusspecies group.Of course,we could not still exclude the possibility of the distribution of theT.asperrimusspecies group in Maolan,and deep field surveys in this area and/or adjacent areas are needed.

Figure 8 Habitats of Tylototriton maolanensis sp.nov.in the type locality,Maolan National Nature Reserve,Libo County,Guizhou Province,China.A,landscape.B,micro-habitat.
Tylototritonnewts were indicated to be poor dispersers and highly associated with specific habitat requirements for breeding and larval development (Zamudio and Wieczorek,2007).InT.wenxianensisspecies group,exceptT.wenxianensis,each species (T.anhuiensis,T.broadoridgus,T.dabienicus,T.liuyangensis,T.lizhenchangiandT.maolanensissp.nov.) has very a narrow distribution,and all of them are indicated to be separated (Wang et al.2018).This indicated that local adaptation might impede the gene flow between them,and would cause great differentiation of species finally resulting in the formation of the new species (Eckertet al.,2008).
Currently,to our knowledge,T.maolanensissp.nov.live in the pond with broad-leaved forests or on the mountainside of Maolan National Nature Reserve.Thus,it is likely that other populations ofT.maolanensissp.nov.may be discovered in the nearby areas such as Mulun National Nature Reserve.We need to conduct deep surveys for investigate its distribution and population status because of increasing human activities and climate changes.
AcknowledgementsWe are grateful to editors and reviewers for their working on the manuscript.We thank Jing LIU for his help on collecting samples.Collections in field were permitted by Administration of Maolan Natioanl Nature Reserve.This work was supported by National Natural Science Foundation of China (No.31960099),Basic research project of science and technology department of Guizhou Province (No.[2020] 1Y083),Science and technology support project of science and technology department of Guizhou Provincial(No.[2020] 4Y029),Guizhou Provincial Department of Education Youth Science and Technology Talents Growth Project (Nos.KY [2018] 455 and KY [2018] 468).Forestry Science and Technology Research Project of Guizhou Forestry Department (No.[2020] 13).
杂志排行

Asian Herpetological Research的其它文章
- Characterization of Skin Symbiotic Bacteria of Sympatric Amphibians in Southeastern China
- Taxonomic Revision of Raorchestes menglaensis (Kou,1990) (Amphibia:Anura),with Descriptions of Two New Species from Yunnan,China
- Reevaluation of the Holotype of Odorrana schmackeri Boettger,1892(Amphibia:Anura:Ranidae) and Characterization of One Cryptic Species in O.schmackeri sensu lato through Integrative Approaches
- Toad-headed Lizard Phrynocephalus forsythii (Squamata,Agamidae) as a Potential Ring Species Inferred from Population Genetic Differentiation
- The Effect of Attendance Patterns on the Lek Mating System in the Omei Treefrog (Rhacophorus omeimontis)
- No Male Preference for Large Females in the Asian Common Toad(Duttaphrynus melanostictus):Effect of the Sex Ratio and Breeding System