贵州蝙蝠分布新记录
——圆耳管鼻蝠
2020-12-28李彦男张翰博钟韦凌石红艳余文华
李彦男, 岳 阳, 张翰博, 张 欣, 钟韦凌, 石红艳, 吴 毅, 余文华*
(1.广州大学 生命科学学院, 广东 广州 510006;2.绵阳师范学院 生命科学与技术学院, 四川 绵阳 621000)
管鼻蝠属(Murina)隶属于翼手目(Chiroptera)蝙蝠科(Vespertilionidae)管鼻蝠亚科(Murininae),该类群体型较小[1],鼻孔呈管状,为森林型蝙蝠.由于其栖息环境较为隐蔽,所以长期缺乏关注,研究标本缺少.近年来,随着研究手段的进步,特别是蝙蝠竖琴网的使用,该类群物种的多样性与本底情况逐渐清晰,迄今全世界已发现有40种,我国现记录19种[1],圆耳管鼻蝠(Murinacyclotis)由Dobon在1872年命名.国外分布区包括不丹、老挝、泰国、印度尼西亚等东南亚地区[2],国内分布在江西、海南、广东和广西等地[3].目前该类群在中国的分布区范围较为狭窄,种群数量少,因此,我国濒危物种红色名录将其列为“近危”等级[4].
本研究基于形态学和分子系统发育学方法,对采集于贵州省的6只管鼻蝠标本进行物种鉴定,结果显示该标本为圆耳管鼻蝠,为该物种在贵州省首次发现记录.现将结果报道如下.
1 研究材料与方法
1.1 标本采集
贵州省荔波县和榕江县属于亚热带季风性湿润气候区,为典型的喀斯特地貌,森林覆盖率高[5].本研究的6只管鼻蝠标本分别于2016年10月采集于贵州省荔波县和2018年7月采集于贵州省榕江县(标本号GZHU16412♂、GZHU16415♂、GZHU16416♂、GZHU16418♂、GZHU16425♀、GZHU18169♂),均为蝙蝠竖琴网捕获,以上标本保存在广州大学华南生物多样性保护与利用重点实验室.
1.2 标本测量
使用数显游标卡尺(MNT-150,上海美耐特实业有限公司,测量精度为0.01 mm)测量标本外形和头骨(精确到0.01 mm).按照哺乳动物测量标准[6].外形指标包括:体重(body weight,Wt)、头体长(head and body length,HB)、尾长(tail length,TL)、耳长(ear length,EL)、前臂长(forearm length,FA)、胫骨长(tibia length,TIB)和后足长(hindfoot length,HF).头骨测量指标包括:颅全长(great length of skull,GTL)、枕犬长(condylo-cannine length,CCL)、脑颅宽(breadth of braincase,BB)、颧宽(zygomatic width,ZW)、眶间宽(interorbital width,IOW)、上犬齿宽(upper canine width,C1-C1)、上齿列长(maxillary tooth,C-M3)、上臼齿宽(third molar width,M3-M3)、下齿列长(mandibular toothrow,C-M3)和下颌长(mandibular length,ML).
1.3 DNA提取与扩增
总DNA提取:使用DNA试剂盒(AG通用型基因组AG21009,艾科瑞生物)提取总DNA.根据管鼻蝠亚科蝙蝠在NCBI-nt(Genbank)数据库中不同分子标记的信息丰度与覆盖度,筛选并使用657 bp左右的线粒体细胞色素氧化酶亚基I基因和1335 bp左右的核基因重组激活基因2为分子标记.扩增引物分别为COI-F:5′-ACA GCC TAA TAC CTA CTC GGC CAT T-3′和COI-R:5′-AGG CTC GGG TGT CTA CGT CCA-3′,PCR反应总体系为30 μL:Premix TaqTM 15 μL;引物各0.4 μL;模板DNA 1 μL;ddH2O 13.2 μL.PCR反应程序:预变性94 ℃ 240 s;变性94 ℃ 30 s;退火55 ℃ 30 s;延伸72 ℃ 60 s,35个循环;最终72 ℃延伸600 s;4 ℃ 保存PCR产物;另外,RAG2-F:5′-GTA AGG ATT TCT TGG CAG GAG T-3′和RAG2-R:5′-ATC CTG CCC CAC TGG AGT TTT C-3′,PCR反应总体系为30 μL:Premix TaqTM 13 μL;引物各0.5 μL;模板DNA 5 μL;ddH2O 11 μL.PCR反应程序为:预变性94 ℃ 180 s;变性94 ℃ 60 s,退火55 ℃ 60 s,延伸72 ℃ 90 s,合计35个循环;最终72 ℃延伸600 s;4 ℃保存PCR产物.扩增成功的PCR产物送往上海生工生物工程公司进行测序.
1.4 构建系统发育树
利用GENEIOUS 4.8.5软件[7]对测序结果进行人工拼接与校对.从NCBI-nt数据库下载管鼻蝠属的COI和RAG2序列,将毛翼管鼻蝠(Harpiocephalusharpia)设置为外群,使用MUSCLE[8]比对排序,通过MEGA 5.0[9]构建最大似然树(Maximum Likelihood Phylogeny)[10],设置Bootstrap值为500估计节点支持度,使用ITOL网页(https://itol.embl.de/)可视化系统发育树.
2 结 果
2.1 形态学特征
为中等偏小体型管鼻蝠,毛发蓬松,鼻部呈不封闭的管状,有对称向上的缺口,鼻吻部有白色触须附着在点状突起上,耳质薄,耳廓圆且平滑,内部有点状结构分布,耳完全展开时无波纹状的嵴,耳折叠时有波纹状的嵴但不明显,耳屏尖细;眼小,眼周缺少毛发附着(图1a);背部毛尖红褐色,中段为金黄色,毛基黑色(图1b);腹部毛发颜色不一,胸部以上为黄褐色,与头部相似,而胸部以下则整体为灰白色,毛基与毛尖颜色差异不大(图1c);体侧膜与第一趾基部相连,后足和胫骨有少量毛发.
头骨结构上,腭桥长,基蝶区有浅的凹陷,犁骨边缘平滑;翼骨突不发达(图1d);矢状嵴和人字嵴不明显,颧弓较为粗壮,鼻骨在前额前微微隆起,中间有凹陷,有眶前孔;齿式为2.1.2.3/3.1.2.3=34,上颌门齿被吻突分割,第1上门齿比第2上门齿小,在侧面观被第2上门齿全部遮挡,第2上门齿与犬齿分开(图1e),第1上前臼齿齿冠比第2上前臼齿略大,第2上前臼齿的齿冠面积约等于犬齿的齿冠面积;第2上前臼齿比第1上前臼齿高但小于上犬齿高,第1上臼齿和第2上臼齿中齿尖不明显,第3上臼齿后跟座退化,仅为第2上臼齿的一半(图1f);下颌骨上隅骨侧面观的轮廓,呈等边三角形状,下颌门齿排列紧密紧挨犬齿,下颌的第1下臼齿齿冠最大,第2下臼齿和第3下臼齿的齿冠依次减小,第3下臼齿的后跟座退化但不如第3上臼齿退化明显(图1g).将本研究的雌雄个体各项指标的测量数据分别与文献[2,11]中圆耳管鼻蝠的数据比对,发现两者高度相符(表1).

图1 圆耳管鼻蝠外部形态和头骨特征(GZHU16412,♂)
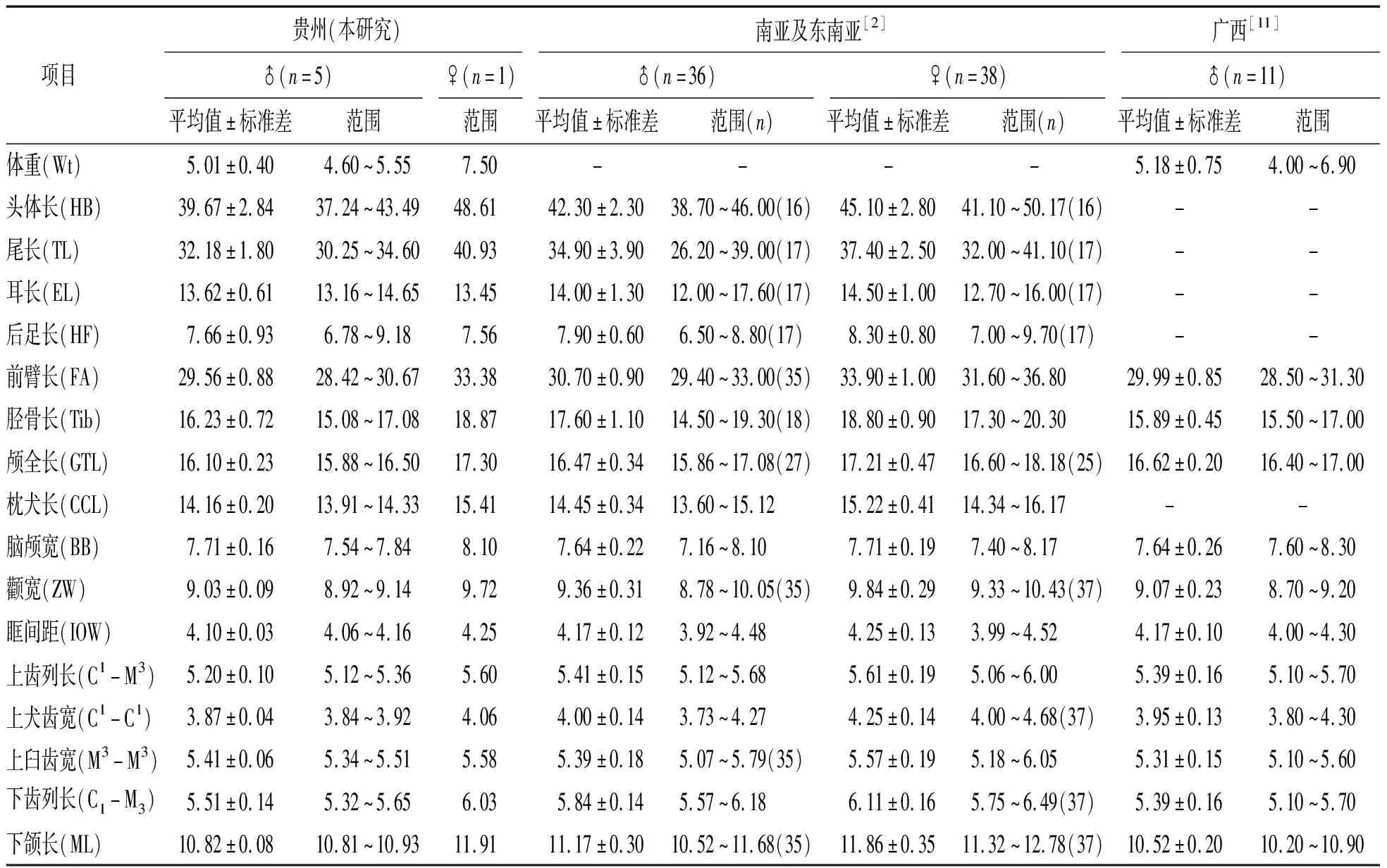
表1 不同地区圆耳管鼻蝠的外形及头骨特征(g, mm)
2.2 系统发育分析
在系统发育树中,本研究样本序列与NCBI-nt中的圆耳管鼻蝠序列聚集成高支持度的一支(自展值100).同时,在线粒体基因COI的亲缘关系上,本研究样本序列不与NCBI-nt上的圆耳管鼻蝠COI序列相混杂,而是聚集在同一支上(图2a);核基因RAG2系统发育树同样提示本研究样本序列与NCBI-nt中的圆耳管鼻蝠聚为高支持度的一支(图2b).因此,基于线粒体基因COI和核基因RAG2构建的系统发育树均支持该批标本为圆耳管鼻蝠.
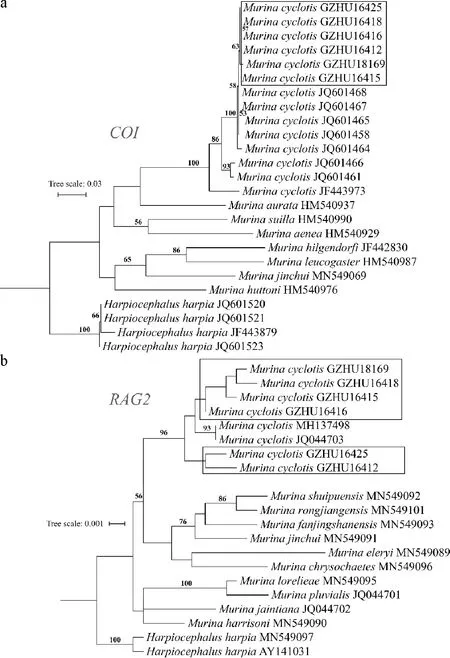
图2 基于线粒体COI基因(a)和核基因RAG2(b)的系统发育树
3 讨 论
在外形上,本研究标本鼻孔呈管状,符合翼手目蝙蝠科管鼻蝠属的特征.其上犬齿和臼齿宽几乎相当,左右齿列呈平行状,第2上门齿与上犬齿分离,上门齿的高度低于上犬齿的高度,同时上犬齿的齿冠面积等于或者大于上前臼齿,与“cyclotis-type”特征相符,而非“sullia-type”[12].根据文献报道该类群管鼻蝠在国内仅三种,分别为:中管鼻蝠(Murinahuttoni)、哈氏管鼻蝠(Murinaharrisoni)及圆耳管鼻蝠,但哈氏管鼻蝠和中管鼻蝠的第1上臼齿和第2上臼齿都有明显的中齿尖[13-14],本研究标本的上臼齿无中齿尖(图1d),与圆耳管鼻蝠相符[2];其外形与头骨的度量数据也与圆耳管鼻蝠吻合(表1).此外,分子系统学结果同样支持本批研究标本为圆耳管鼻蝠.综上,鉴定在贵州采集的标本为圆耳管鼻蝠.
圆耳管鼻蝠模式产地位于印度,国外分布在东南亚等地区,包括:尼泊尔、缅甸、越南、泰国、老挝和印度等国家[12].在我国该物种目前分布于华南区域,包括:江西、广东、广西和海南等省份,但其种群状况未明,标本数量较少[15-17].在贵州发现该物种的分布尚属首次,进一步补充了该物种的本底资料,为贵州省翼手目分布新记录.值得注意的是该种蝙蝠在亚洲广泛分布,且已有形态学地理变异的报道[2],故后续有必要进一步进行细致的物种厘定和亚种分化研究.
