3个山茶品种对干旱胁迫的生理响应
2020-12-25赖路伟黄永芳叶小萍谭健彬许雄坚
谭 莎,赖路伟,黄永芳,叶小萍,谭健彬,许雄坚
(1.佛山市云勇生态林养护中心,广东 佛山 528183;2.旭辉集团,福建 厦门 361000;3.华南农业大学林学与风景园林学院/广东省森林植物种质创新与利用重点实验室,广东 广州 510642)
茶花是山茶科(Theaceae)山茶属(Camellia)植物的统称,其适应性强,开花周期长,且在少花的冬季开花,在园林绿化中广泛应用[1]。茶花种类繁多,但应用在风景园林中的种类较少,园林植物需精细化水肥管理[2],干旱会限制植物的正常生长[3],故选择抗旱性的观赏茶花尤为重要。目前有关干旱胁迫对茶花生理响应的研究鲜见报道[4—5]。本试验主要研究‘红露珍’(C.japonica ‘Hongluzhen’)、‘六角大红’(C.japonica ‘Hongliujiao’)和‘烈香’(C.japonica‘High Fragrance’) 3个山茶品种在干旱胁迫下的生理生化特性,以期揭示茶花耐旱响应机理。3个山茶品种广泛应用于园林绿化[6—8],应用前景广阔,通过研究其干旱胁迫生理响应,可为茶花在风景林中的应用提供理论依据和技术支持。
1 材料与方法
1.1 材料
2017年4月从广州岭南花卉市场分别选取‘红露珍’、‘六角大红’和‘烈香’生长健壮且长势一致、苗高55~60 cm、地径1 cm的2年生扦插苗木各30株,移栽于直径21 cm、高21 cm花盆中,所用土壤为黄心土和营养土以 1:2混合而成。苗木露天栽培,常规管理。放置在华南农业
1.2 试验设计
采用盆栽连续干旱法,于2017年8月9日将试验苗木浇透水,3个山茶品种各随机选取 6株以浇水处理作为对照,其余试验苗木之后不再浇水,对其进行干旱处理,分别在干旱处理4 d、8 d、10 d进行采样,每个品种随机选择 3~6片功能叶,共10~20叶片,每个样品 3次重复。样叶清洗后于-80 ℃超低温冰箱中保存备用。同时,在干旱处理2 d、4 d、6 d、8 d、10 d对3个品种进行土壤含水量测定,每处理3个重复,于早上8:00前测定。
1.3 检测方法
土壤含水量采用美国产MI2X型土壤水分探头测定;叶片相对含水量采用饱和称量法测定;叶绿素含量测定采用80%丙酮提取法;丙二醛(MDA)含量采用硫代巴比妥酸(TBA)法测定;质膜透性采用相对电导率法测定;可溶性蛋白含量采用考马斯亮蓝法测定;超氧化物歧化酶(SOD)活性采用氯化硝基四氮唑蓝(NBT)光化还原法测定;脯氨酸含量采用酸性茚三酮法测定[9]。
1.4 数据处理
采用SPSS 19.0软件作方差分析和Ducan多重比较。采用模糊隶属函数法对3个山茶品种苗木抗旱指标进行综合评价[10]。隶属函数值计算方法如下。
(1) 当测定指标与综合评判结果呈正相关:
U(Xi)=(Xi-Xmin)/(Xmax-Xmin);
(2) 当测定指标与综合评判结果呈负相关:
U(Xi)=1-(Xi-Xmin)/(Xmax-Xmin)。
式中,U(Xi)为某一植物在某测定指标的隶属函数值;Xi为植物某一指标的平均值;Xmax、Xmin分别为所有材料平均测定值中最大值、最小值。
2 结果与分析
2.1 干旱胁迫对土壤含水量的影响
随着干旱处理时间的延长,3个山茶品种的土壤含水量明显下降(图1)。在干旱10 d时,3个山茶品种土壤含水量均与对照存在显著差异(P<0.05)。‘六角大红’土壤含水量最低,为8.1%,‘红露珍’次之。‘红露珍’土壤含水量与对照相差最大,差距达45.7%。
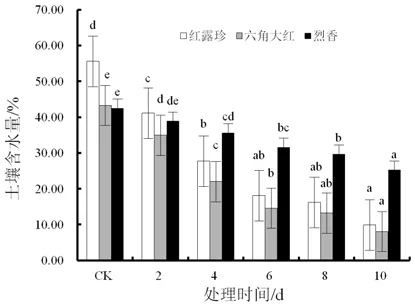
图1 干旱胁迫下3个山茶品种的土壤含水量变化Fig.1 Changes of soil water content of three Camellia cultivars under drought stress
2.2 干旱胁迫对叶片生理生化指标的影响
2.2.1 相对含水量
随着干旱处理时间的延长,3个山茶品种叶片相对含水量变化不同(图2)。3个山茶品种在胁迫前相对含水量相近,干旱胁迫期间,‘红露珍’叶片相对含水量无显著变化,‘六角大红’和‘烈香’叶片相对含水量整体呈下降趋势。在干旱10 d时,‘六角大红’和‘烈香’的叶片相对含水量均显著低于对照(P<0.05)。
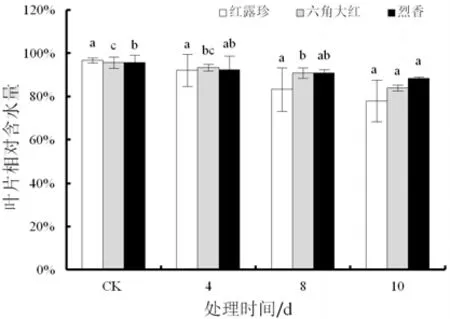
图2 干旱胁迫下3个山茶品种叶片相对含水量变化Fig.2 Changes of leaf relative water content of three Camellia cultivars under drought stress
2.2.2 叶绿素含量
随着干旱时间的延长,3个山茶品种叶绿素含量呈下降趋势(图3)。在干旱4 d时,3个山茶品种叶绿素含量大幅度下降,‘红露珍’叶绿素含量最低,下降 37.82%;‘六角大红’叶绿素含量下降幅度最大,下降47.58%。3个山茶品种干旱4 d、8 d的叶片叶绿素含量差异不显著(P>0.05),在干旱10 d时,各品种叶绿素含量与对照皆差异显著(P<0.05)。

图3 干旱胁迫下3个山茶品种叶片叶绿素含量的变化Fig.3 Changes of chlorophyll content in the leaves of three Camellia cultivars under drought stress
2.2.3 丙二醛(MDA)含量
随着干旱时间的延长,3个山茶品种丙二醛含量逐渐上升(图4)。干旱4 d,‘红露珍’和‘烈香’丙二醛含量上升幅度较大,分别比对照上升34.00%、23.81%。干旱 10 d,‘红露珍’、‘六角大红’、‘烈香’与对照相比上升93.00%、76.84%、63.23%,三者与对照差异皆显著(P<0.05)。
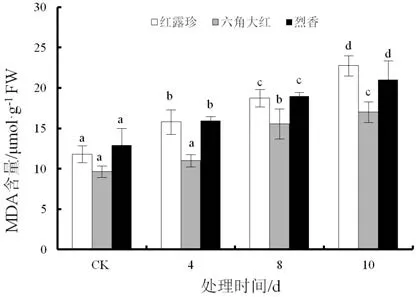
图4 干旱胁迫下3个山茶品种叶片丙二醛(MDA)含量变化Fig.4 Changes of malondialdehyde (MDA) content in the leaves of three Camellia cultivars under drought stress
2.2.4 相对电导率
随着干旱胁迫时间的延长,3个山茶品种的叶片电导率明显上升(图5)。干旱4 d,3个山茶品种的叶片电导率上升幅度基本一致。干旱8 d,3个不同山茶品种电导率上升幅度较大,‘红露珍’幅度最大,电导率增至72.58%。3个山茶品种干旱处理后叶片相对电导率与对照均存在显著性差异(P<0.05)。
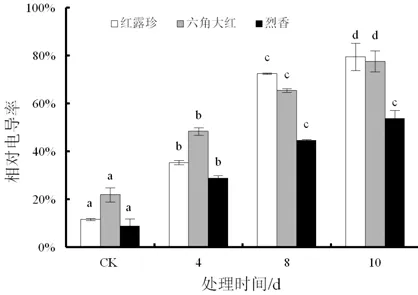
图5 干旱胁迫对3个不同山茶品种叶片相对电导率的影响Fig.5 Effects of drought stress on the leaf relative conductivity of three Camellia cultivars
2.2.5 可溶性蛋白质含量
随着干旱胁迫时间延长,3个山茶品种可溶性蛋白含量逐渐上升(图6)。干旱10 d时,3个山茶品种可溶性蛋白质含量达到最大值,不同品种的增幅有所差异。‘红露珍’在干旱胁迫期间上升幅度不大,增至1.577 mg·g-1FW,‘六角大红’和‘烈香’的上升幅度基本一致,含量分别达 2.82 mg·g-1FW 和2.25 mg·g-1FW,三者皆与对照差异显著(P<0.05)。
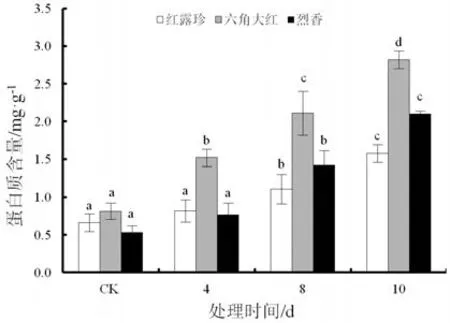
图6 干旱胁迫下3个山茶品种叶片可溶性蛋白质含量变化Fig.6 Changes of soluble protein content in the leaves of three Camellia cultivars under drought stress
2.2.6 超氧化物歧化酶(SOD)活性
随着干旱胁迫时间延长,‘红露珍’和‘烈香’的叶片超氧化物歧化酶(SOD)活性逐渐上升(图 7)。‘六角大红’和‘烈香’干旱初期SOD活性上升,后趋于平稳。干旱10 d时,‘红露珍’SOD活性达最大值,为414.904 U·g-1FW。
2.2.7 脯氨酸含量
随着干旱胁迫时间的延长,3个山茶品种叶片脯氨酸含量整体呈上升的趋势(图 8)。‘红露珍’在干旱胁迫期间上升幅度不大,‘烈香’和‘六角大红’脯氨酸上升幅度较大;干旱4 d,3个不同山茶品种脯氨酸含量与对照相比差异不显著;干旱8 d,3个不同山茶品种的脯氨酸含量急剧升高,‘六角大红’变化最大;干旱10 d,3个山茶品种脯氨酸含量与对照存在显著性差异(P<0.05)。
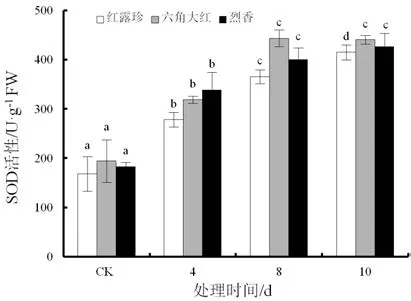
图7 干旱胁迫下3个山茶品种叶片超氧化物歧化酶(SOD)活性变化Fig.7 Changes of SOD activity in the leaves of three Camellia cultivars under drought stress
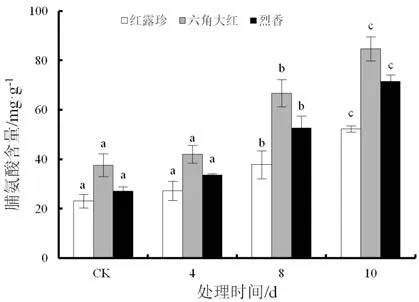
图8 干旱胁迫下3个山茶品种叶片脯氨酸含量变化Fig.8 Changes of proline content in the leaves of three Camellia cultivars under drought stress
2.3 山茶3个品种抗旱性综合评价
以生理生化指标作为抗旱性综合评价指标,对不同品种的抗逆性进行分析,比较3个山茶品种抗旱性。对7个指标的隶属函数分析表明,‘六角大红’综合抗旱能力强于‘红露珍’和‘烈香’(表1)。
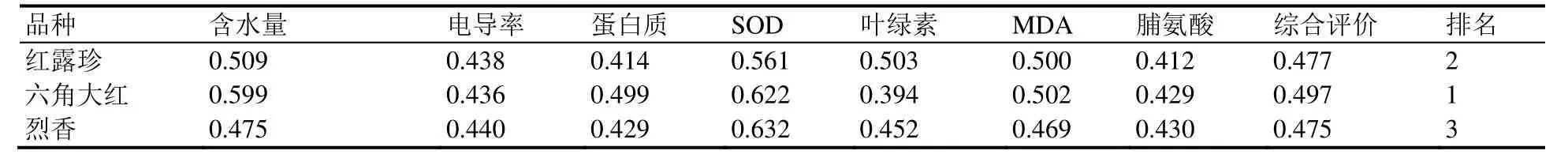
表1 干旱胁迫下3个山茶品种叶片7个指标隶属函数值Table 1 Subordinate function values of 7 indexes of three Camellia cultivars under drought stress
3 结论与讨论
本试验在8月开展,此时高温少雨,空气相对湿度较低,土壤水分减少主要由蒸发和植物蒸腾作用引起。随着干旱胁迫加剧,3个山茶品种栽培土壤含水量均显著低于对照,‘烈香’土壤含水量最高,初始值与‘六角大红’相差不大,随着干旱胁迫处理,差距越来越大。这可能与叶片数量和面积有关,‘烈香’叶面积小于‘红露珍’和‘六角大红’,说明植物形态结构在一定程度上间接影响抗旱能力。
叶片相对含水量(RWC)与植物代谢活动密切相关,RWC是衡量植物抗旱性的常用生理指标[11]。有研究表明,叶片 RWC保持较高水平是对干旱适应的表现[12]。本试验中,3个山茶品种叶片相对含水量下降缓慢,表明3个山茶品种的土壤相对含水量不决定其叶片相对含水量。
叶绿素含量在一定程度上反映植物的同化能力及其生长状况[13],干旱胁迫对光合色素有显著影响,使叶绿素分解速率大于合成速率,影响植物光合作用。抗旱性强的植物叶绿素含量降低的幅度相对较小[14]。本试验中,随着干旱胁迫延长,3个山茶品种叶绿素含量均下降,‘六角大红’下降幅度最大。
逆境胁迫对植物细胞质膜有重要影响,通常表现为膜结构遭到破坏,选择透性丧失,从而导致电解质和一些小的有机化合物大量渗入[15]。丙二醛(MDA)是植物在逆境环境中抵抗逆境时的产物。相对电导率和 MDA含量变化是脂膜过氧化程度和植物对逆境条件反应强弱的重要指标[16]。当植物细胞膜损伤破裂,细胞内电解质外渗,相对电导率越高,表明植物损害越大,植物抗旱性越弱。本试验中,3个山茶品种相对电导率随着干旱胁迫的延长而增大,丙二醛含量呈不同程度的上升趋势。
可溶性蛋白和脯氨酸含量是植物体内重要的渗透调节物质,也是植物耐旱性的重要指标[17]。在植物遭到逆境胁迫时可平衡细胞代谢,以维持细胞内环境的稳定[18]。可溶性蛋白具有较强的亲水胶体性质,直接影响细胞的保水性,蛋白质含量增幅越大,越有利于水势降低,增加植株吸水能力和抗旱性[19],本试验中,3个山茶品种可溶性蛋白含量均增加,‘六角大红’表现尤为明显,可溶性蛋白含量与抗旱性呈正相关[20]。脯氨酸作为氧化还原剂,可消除植物体内氨积累所造成的伤害[21],一定范围内能维持细胞膨压稳定[22]。研究发现,植物干旱胁迫初期脯氨酸含量逐渐增加后下降[23—24]。本试验显示,随着干旱胁迫时间延长,3个山茶品种脯氨酸含量逐渐上升,干旱处理10 d,3个山茶品种叶片脯氨酸含量与对照存在显著性差异,表明3个山茶品种具有一定的耐旱能力。
超氧化物歧化酶(SOD)是保护酶系统的重要酶之一,有助于保护细胞,减轻应激造成的损害。在干旱胁迫下,SOD酶活性相对下降,可通过 SOD活性变化指示植物的抗旱性能[25]。本试验中,‘红露珍’SOD活性随着干旱胁迫延长而升高;‘六角大红’和‘烈香’在干旱8 d后趋于平缓,表明3个山茶品种在轻度干旱下通过抗氧化系统提高抗旱能力。
植物的抗旱性是一个复杂的综合性状,是从植物的形态构造、生理生态特征及生理生化反应到细胞、光合器官及原生质结构特点的综合反映[26]。本试验3个山茶品种抗旱性的综合比较表明,‘六角大红’抗旱性最强,‘烈香’最弱。