广东石门台假林线的物种组成及优势种种群特征
2020-12-16张琪杜建会李远球李升发邵佳怡
张琪,杜建会, ,李远球,李升发 ,邵佳怡
1. 中山大学地理科学与规划学院,广东 广州 510275;2. 广东省城市化与地理环境空间模拟重点实验室,广东 广州 510275;3. 广东石门台国家级自然保护区管理局,广东 英德 513000;4. 广州地理研究所/广东省地理空间信息技术与应用公共实验室,广东 广州 510070;5. 广东省遥感与地理信息系统应用重点实验室,广东 广州 510070
在亚热带山区,矮曲林作为山顶重要的生态屏障,与其分布海拔之下的竹林、亚热带针叶林及常绿阔叶林等组成了相对稳定的生态系统,其物种组成及群落动态对于山地生态系统的水源涵养、水土保持、生物多样性保护及碳储存等具有非常重要的意义(Grafius et al.,2015;姚永慧等,2017)。但受自然或人为活动干扰,矮曲林被砍伐或烧毁,山顶因缺乏植被保护,加上坡度较陡和强降雨影响,使得该处土壤侵蚀加剧,土层变薄、养分流失。此外,山顶气温较低、辐射强烈、风力强劲、蒸发量大,进而使得森林的次生演替过程受到很大的抑制,原本发育森林的地方被灌丛或草甸所占据,森林则只能出现在距离山顶一定范围的高度(Grubb,1971;姚永慧等,2017;张琪等,2018)。山顶灌草丛与森林接触之处形极规则,视同横线,前人将这种由山顶效应形成的突变,且分布在气候林线之下的森林边界称为假林线(Gersmehl,1973;叶居新,1989)。
假林线在亚热带山地普遍存在,许多学者针对其起源、物种组成和土壤特性等展开调查,对于假林线的群落更新动态进行了初步研究,发现在人类活动中止后,受物种生物学特性和区域生境影响,假林线向上缘草坡均存在不同程度的入侵,但变化不显著(徐捷等,2010;Bowman et al.,2017;张琪等,2018)。树木的种子萌发、幼苗生长和幼树的形成等受树种组成、生境特征及干扰竞争等多个因素影响,任何一个环节出现障碍都可能会影响到森林的天然更新(赵高卷等,2016;张小鹏等,2018)。但假林线分布的区域地处偏远、交通不便,在一定程度上限制了对其进行大范围的深入调查,目前尚不清楚假林线附近的树木在人类活动中止后能否持续更新,难以准确评估中国亚热带山地森林的恢复潜力(张金泉,2016)。种群特征可反映其个体数量的分配情况、形成过程和发展趋势,也可表征树种对生境的适应策略,这对揭示种群的更新和维持机制具有非常重要的意义(费世民等,2008;Li et al.,2020)。本文通过对广东石门台国家级自然保护区内的假林线典型分布区木龙顶样地进行群落物种调查,并对其乔木层优势种的树高、胸径和生物量等进行测量及采样分析,有助于理解人类活动中止后,亚热带山地森林的次生演替过程,这对于准确评估亚热带山地生态系统的服务价值也具有重要的意义。
1 材料与方法
1.1 地理位置
广东石门台国家级自然保护区位于广东省英德市西北部,北与韶关市的乳源、曲江县相邻,西起波罗镇的赤泥岭,东至沙口镇的江溪,东西宽约43.66 km,南北长约15.48 km,总面积335.55 km2。地势西北高、东南低,兼有平原、丘陵、山地和喀斯特地形的多样性地貌,最高峰船底顶海拔1586.8 m(张琪等,2018)。早期因大炼钢铁,大面积常绿阔叶林被砍伐用于燃料,加上后续村民伐木建屋、毁林种果、种竹笋,使得原始森林遭到严重破坏。英德市人民政府首先在 1998年于石门台成立县级自然保护区,2012年将其升级为国家级自然保护区,并全面实行封山育林,严禁砍伐等措施,使得次生植被得到了良好的保护,除少量的户外活动外,核心区无其它居民居住(张金泉,2017)。
1.2 数据来源及研究方法
1.2.1 植物群落调查
在线路踏查的基础上,选取木龙顶样地作为研究区(24°28′42.1″—24°28′43.0″N,113°14′15.0″—113°14′17.2″E),其海拔为 1118 m,土壤类型以山地草甸土为主。采用相邻格子法,于2017年11月中旬沿着假林线边缘设置6个10 m×20 m样方,并在样方内沿对角线设置2 m×2 m小样格。考虑到研究区生境严酷,疏齿木荷主要以幼苗为主,且后勤难以补给,因此在保证数据测定精度的基础上,以树高≥2 m且胸径≥2.5 cm作为乔木层起测标准,进行每木调查并记录种名、高度、胸径、枝下高、冠幅和林分郁闭度,胸径小于2.5 cm的幼苗只统计其高度及数量;对于灌木层则记录其种名、株数、平均高度和盖度;草本层记录其种名、平均高度和盖度;藤本植物记录其种名。用GPS测定海拔和经纬度,用坡度计测定坡向和坡度。
1.2.2 重要值
乔木层和灌木层重要值(IV)的计算基于相对多度(ra)、相对频度(rf)和相对盖度(rc);草本层以相对高度(rh)替换相对多度(ra)。各参数及植被盖度(C)分别采用以下公式计算,其中真盖度指乔木胸径处的截面积,基盖度指灌木基径处的截面积(乔木层和灌木层重要值根据公式 (6) 计算;草本层重要值根据公式 (7) 计算):

上式中,W为实际测量的植物胸径;

上式中,mi为第i个物种的株数,Σmj为全部物种的总株数;

上式中,fi为第i个物种的频数,Σfj为全部物种的频数之和;

上式中,ci为第i个物种的盖度,Σcj为全部物种盖度之和;灌木层直接用估算的盖度进行计算;

上式中,hi为第i个物种的平均高度,Σhj为全部物种的平均高度。

1.2.3 多样性指数
本研究采用重要值作为优势度指标以确定植物群落各层的优势种(王伯荪等,1996),以辛普松指数(Simpson Index,D)和香农-维纳指数(Shannon-Wiener Index,H′)对乔灌层及群落的多样性水平进行分析。两种指数计算公式如下:

上述两式中,Pi=Ni/N,其中Ni为样方内第i个物种的个体数,N为样方内所有物种的个体数之和。
1.2.4 疏齿木荷生物量测定
为避免对固定样方内的植被造成干扰,在样地附近选取不同径级的疏齿木荷(Schima remotiserrata)作为标准株。考虑到绝大多数的植株胸径均小于6.5 cm,且样地位于保护区范围内,无法进行大规模破坏性采样,因此对胸径≤6.5 cm的样本,共选择了 11株,使用小铁铲沿着其根系(主要分布在0—20 cm)的延伸方向,采用全挖法将其整株挖出。对于>6.5 cm的样本,以胸径间隔2 cm为径级划分标准,在样地附近选择标准木28株,现场砍伐,以充分代表疏齿木荷的种群特征。所有样品按器官分为树干、树枝和树叶,并在样地迅速称取鲜重,用于构建单株地上生物量与其树高及胸径之间的关系。由于山顶缺乏水源,胸径≤6.5 cm的植株根系泥土粘连较多,因此将其全部带回,仔细清理干净后,测定其鲜质量,并计算根冠比,用于估算胸径>6.5 cm的植株地下生物量。研究区山高路远,运输不便,因此将植株地上样品分器官充分混合,依照比例选取部分样品带回实验室。所有样品均在烘箱内设置70 ℃烘干(24 h)至恒质量,分别记录树干、树枝、树叶和树根生物量,并计算其干鲜重比。最后结合不同径级的立木密度,估算木龙顶样方内的疏齿木荷各个径级的生物量及总生物量。
1.3 数据分析
相关数据采用SPSS 13.0(SPSS,Chicago,IL,USA)进行统计分析,作图使用Origin 9.0(Origin,Northampton,USA)进行绘制。
2 结果与分析
2.1 群落组成及物种特征
2.1.1 群落外貌
木龙顶样地植物群落主要以亚热带常绿阔叶林为主,外貌整齐,林冠浓密,郁闭度达0.85,极少林窗。群落组成中虽有少数落叶树种,但并不构成明显的落叶季相,常与一定数量的针叶树种混交成林。林冠呈圆形或椭圆形,乔木分层不明显,植株低矮,群落高度普遍在 3—5 m,平均树高(3.5±0.9) m,平均胸径 (4.7±2.8) cm,枝下高平均约(1.3±0.6) m,偶见少数针叶树种高于林冠之上,但最高亦不超过7 m。本次调查的1200 m2样地中,高度在5 m以上的树木仅26株。群落内胸径≥2.5 cm的立木平均密度为3900 plant·hm-2,林木生长稠密,多小型或中型叶、革质且多茸毛,植株通常分枝低矮且侧枝发育扭曲,树干多附生有苔藓类植物。藤本种类较少,仅见少量草质和木质藤本。根据以上群落基本特征,可将该群落划入山顶苔藓矮曲林。
2.1.2 群落组成与物种多样性
(1)物种组成
木龙顶样地内共有维管植物53种,隶属33科,43属(表1)。其中,蕨类植物1科1属1种,裸子植物2科2属2种,被子植物30科40属50种。群落物种组成以杜鹃花科6种最多,其次是樟科4种,禾本科、冬青科和百合科各3种,茜草科、金缕梅科、山茶科、山矾科、木兰科和野牡丹科各 2种,其余各科各1种。以亚热带表征科为数量优势科,充分体现了植被的水平地带性分布特征。
木龙顶样方乔木层中的疏齿木荷重要值最高(26.01%),其次为五列木和华南五针松(表2)。样方乔木层平均树高普遍较小,华南五针松的平均高度(4.39 m)略高于五列木(3.44 m)和疏齿木荷(3.21 m),但其数量远低于后两者,相对多度较小。样方乔木层中五列木数量最多,相对多度最大,但其相对盖度及相对显著度明显小于疏齿木荷,因而后者成为样方乔木层的优势种。
灌木层中五列木分布均匀,生势良好(表3),其数量虽略少于广东杜鹃(Rhododendron kwangtungense),但总截面积是后者近10倍,故相对盖度在木龙顶样地的灌木层中位于首位,可判断五列木为样方内灌木层的优势种,而其它物种在灌木层中的生势则较为均衡。
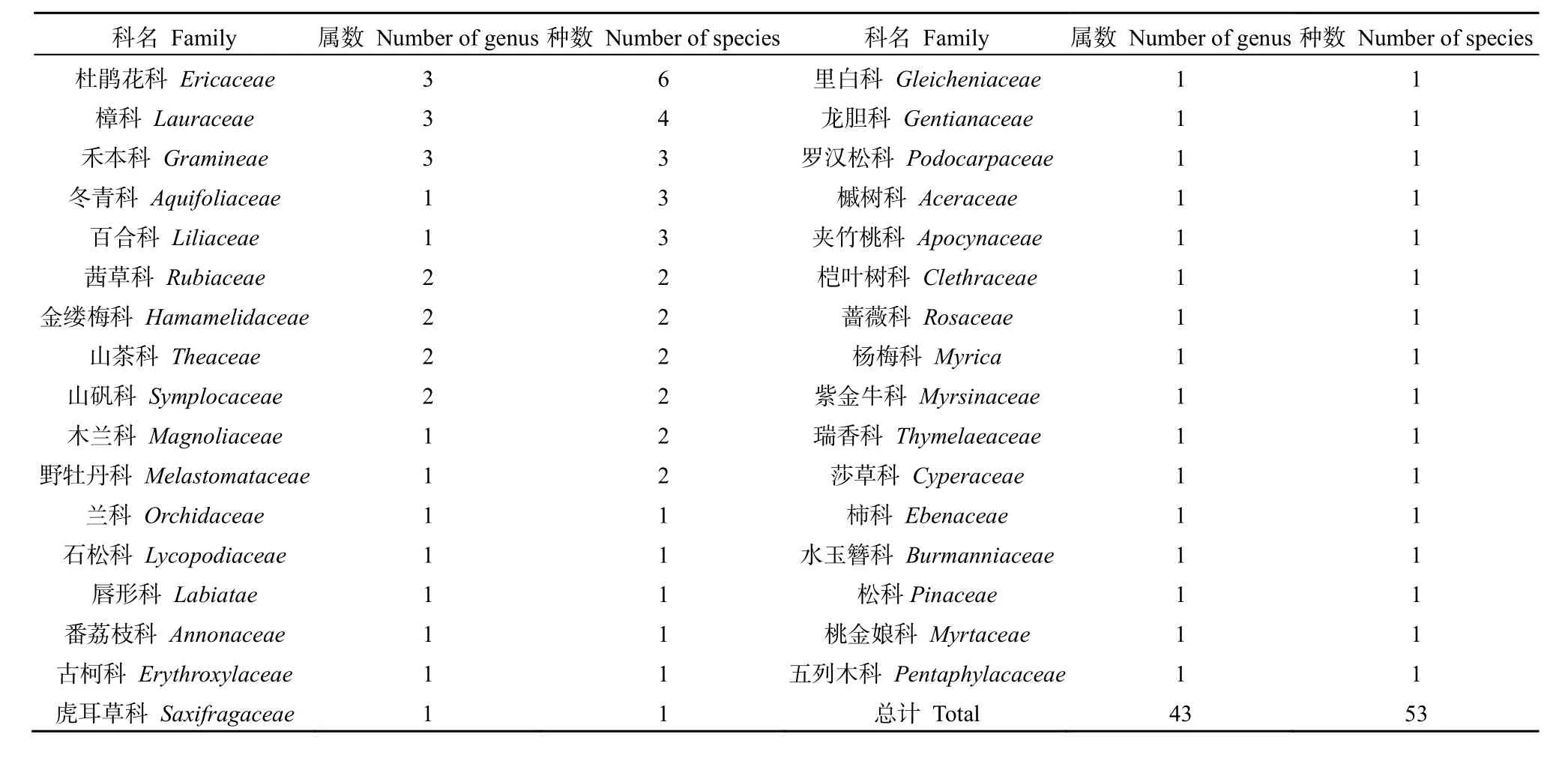
表1 群落维管植物分布特征Table 1 The distribution characteristics of community of vascular plants
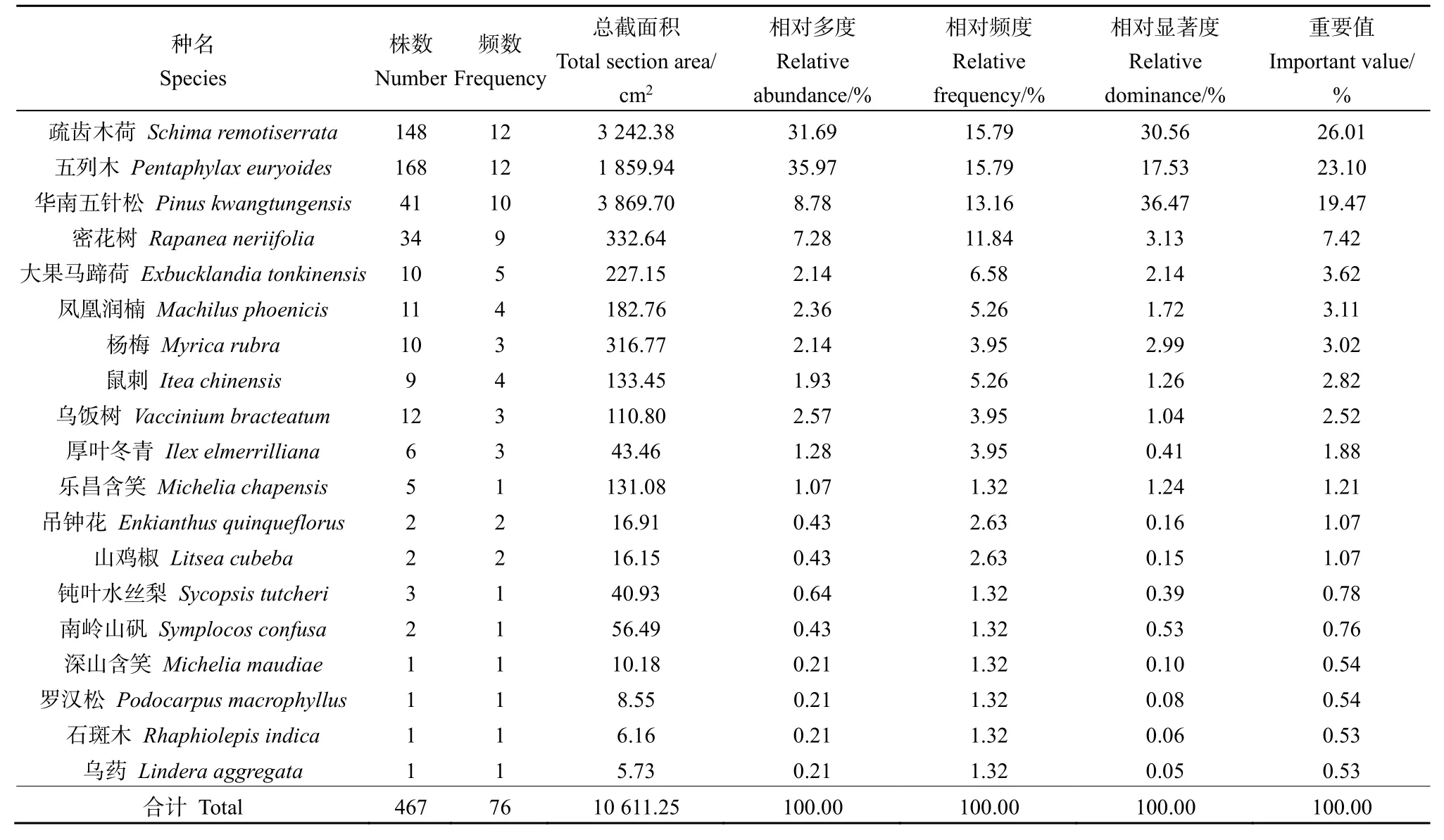
表2 木龙顶样方乔木层特征值Table 2 Important value of tree layer plots at Mulongding Mountain

表3 木龙顶样方灌木层特征值Table 3 Important value of shrub layer plots at Mulongding Mountain
草本层中的野古草(Arundinella anomala)分布均匀(表4),且生势良好,其相对频度与相对盖度均居于样方首位且远大于其余物种,重要值达到47.84%,成为该样方草本层的优势种。
此外,木龙顶样方内的层间植物还包括藤本灌木链珠藤(Alyxia sinensis)、土茯苓(Smilax glabra)、草质藤本藤石松(Lycopodiastrum casuarinoides)、攀援灌木菝葜(Smilax china)、暗色菝葜(Smilax lanceifolia)、羊角藤(Morindaumbellata)等6种。
根据以上调查结果可知,样方内共记录有 19种乔木,群落优势种为疏齿木荷,其截面积为88427 cm2·hm-2。灌木层记录物种数 37种,平均高度(1.48±1.33) m。草本层记录物种数8种,平均高度(0.38±0.24) m。依据各层物种的重要值(表2—4),可将该样地植物群落命名为疏齿木荷-五列木-野古草群丛(Ass.S.remotiserrata-Pentaphylax euryoides-Arundinella anomala)。
(2)物种多样性指数
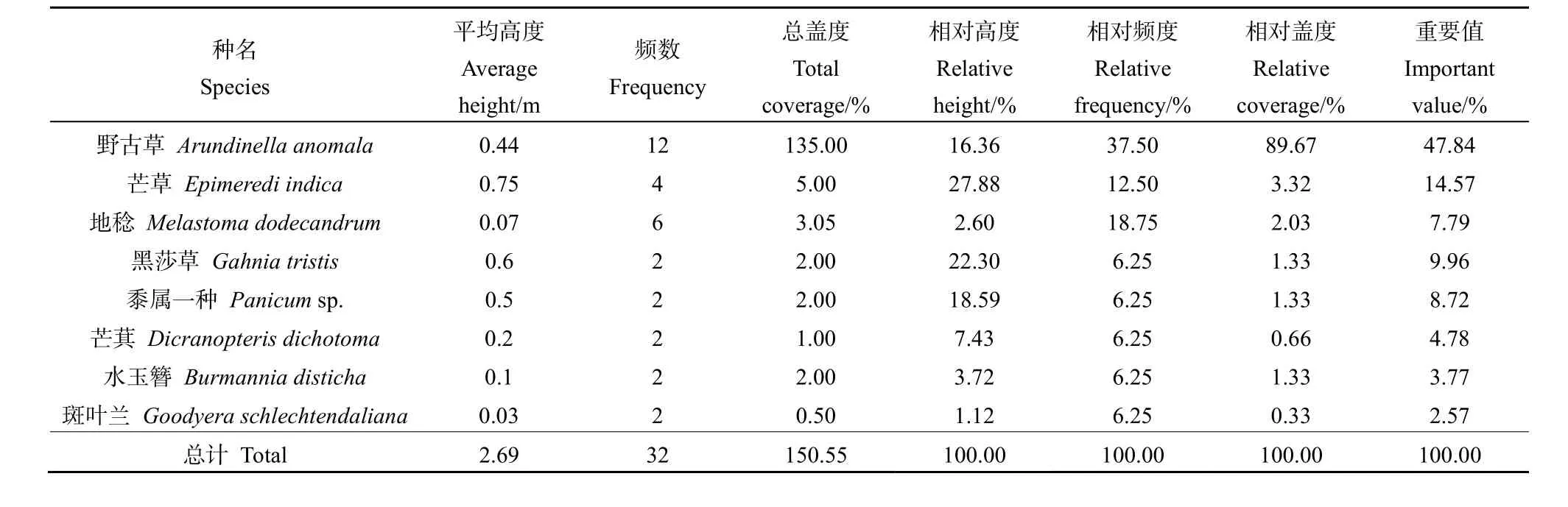
表4 木龙顶样方草本层特征值Table 4 Important value of herb layer plots at Mulongding Mountain
样方植物群落的物种多样性指数D为0.882,H′为 3.652,表明群落物种多样性水平较高。其中乔木层D为0.754,H′为2.624;灌木层D为0.879,H′为 3.679。两种多样性指数在乔木层和灌木层变化趋势一致,均表现为灌木层的物种多样性水平高于乔木层。
2.2 疏齿木荷的种群特征
2.2.1 疏齿木荷种群的结构特征
样地内疏齿木荷种群的平均高度为 (3.22±0.84)m,以小于1.0 m的植株为主,其数量占疏齿木荷种群的88.8%,高度在5 m以上的仅5株,最高为7.0 m,群落整体矮化,分枝较低(图 1A)。疏齿木荷种群的平均胸径为 (4.60±2.25) cm,以胸径≤2.5 cm的植株为主,密度达到15417 plant·hm-2,占疏齿木荷种群的92.3%。随着胸径增长,其个体数量呈现出显著的下降趋势,胸径>2.5 cm的植株较少,其密度仅有1283 plant·hm-2。其中胸径在2.5—6.5 cm范围内的植株占6.8%,而胸径大于6.5 cm以上的植株数量仅占总数的0.9%(图1B)。
2.2.2 疏齿木荷种群的生物量特征
不同径级的疏齿木荷生物量差异较大,除树叶外,其余器官的生物量均随着径级的增加呈现出先上升后下降的趋势(表5)。胸径≤6.5 cm的疏齿木荷总生物量为4.73 t·hm-2,占总生物量的51.88%,而胸径>6.5 cm的疏齿木荷尽管株数较少,但单株生物量大,占到总生物量的48.12%。疏齿木荷各器官的生物量占比也有所不同,从大到小依次为树干、树根、树枝和树叶。树干生物量占总生物量的比例达52.20%,而树叶生物量仅占总生物量的6.01%。本文采样时间为11月中旬,恰好对应了疏齿木荷的换叶期,可能低估了树叶的生物量。依据样方的立木密度调查数据,基于胸径和树高模型估算可得研究区的疏齿木荷地上生物量为7.11 t·hm-2。疏齿木荷根冠比波动范围在 0.15—0.48之间,且随着胸径的增大先快速减小,后逐渐稳定,依据其根冠比可估算疏齿木荷的总生物量为9.10 t·hm-2。
3 讨论
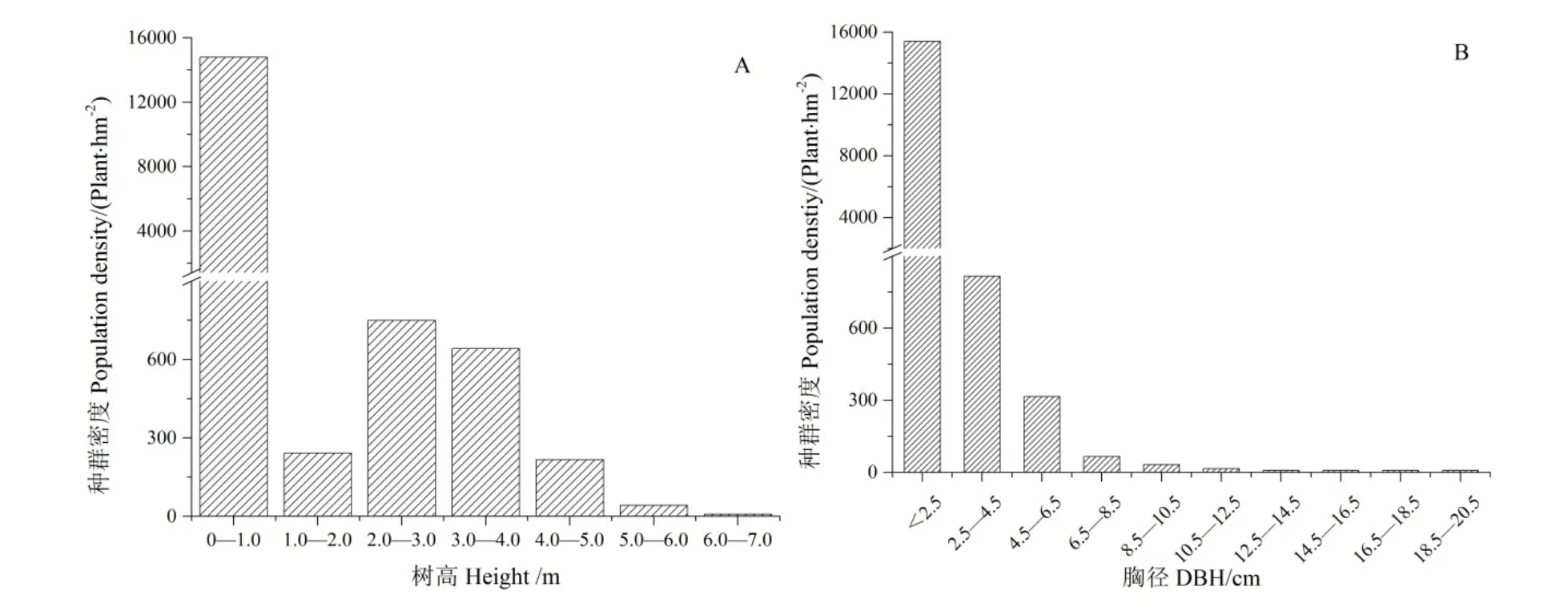
图1 木龙顶样方中的疏齿木荷种群胸径及树高频数分布特征Fig. 1 Population density in height (A) and DBH-class (B) of S. remotiserrata in Mulongding Mountain

表5 木龙顶样方不同径级的疏齿木荷各器官生物量特征Table 5 Biomass of S. remotiserrata among different diameter classes in Mulongding Mountain
森林群落的物种组成受生境影响明显(陈林等,2010)。对木龙顶假林线样方的调查发现,其物种组成主要以乔灌木为主,草本和藤本植物相对较少。这与王厚麟等(2008)对中国亚热带山顶矮林的研究结果比较一致。假林线林冠层的郁闭度较高,往往造成林下光照不足,且枯枝落叶多,不利于草本和藤本植物种子萌发及根系下扎,从而抑制其后续生长(陈林等,2010)。样方内的乔木层高度普遍较矮,且扭曲生长,其平均树高为(3.5±0.9) m。这与陈林等(2010)对南岭国家自然保护区的山顶矮林调查结果较为接近,其乔木层的平均高度为3.23 m,以5 m以下的个体为主。Shiels et al.(2001)对科罗拉多Goliath山地的刺果松(Pinus aristata)和恩格曼云杉(Picea engelmannii)矮曲林研究也发现,其高度大小在0.5—3 m之间。假林线多分布在保护区的山顶或山脊部位(张琪等,2018),该处风力强劲、湿冷多雾、温差大,致密且低矮的林冠层可有效降低风力造成的机械损伤,并能减少辐射加热和土壤热通量交换,林冠层下的热量散失较慢,其叶片组织和根际温度常高于直立生长的高大树木,有利于提高矮曲林的光合作用水平,缓解极端气温对树木生长的影响(Grubb,1971;McIntire et al.,2016;Kašpar et al.,2017)。
树木在不同的生长阶段受限因素存在差异,进而影响到其种群的结构特征(王树力等,1993)。对疏齿木荷种群的树高特征研究发现,其呈现为反“J”型,以高度小于1 m的植株为主,1—2 m高度的植株占比急剧减小。说明疏齿木荷的天然更新存在瓶颈现象,其小于1 m的植株死亡率较高,只有少部分植株能够存活,并进入到下一生长阶段。这与宋立奕等(2013)对云南元谋干热河谷膏桐(Jatropha curcas)种群的天然更新研究较为一致,其认为膏桐幼苗可能与高度近1 m的多年生草本植物扭黄茅(Heteropogon contortus)产生了强烈的竞争,导致高度小于1 m的膏桐幼苗生长受到了很大的抑制。木荷属植物的更新主要与自然或人类活动形成的林窗有关(Tang et al.,2020)。对研究区的调查发现,草本植物中重要值较高的如芒草等平均高度可达到0.75 m,灌木层平均高度为1.48 m,其与高度小于1 m的疏齿木荷幼苗共同竞争空间和光照等,在一定程度上可能抑制了疏齿木荷的幼苗成长为幼树。但一旦突破这一高度,其在与草本及灌木的竞争过程中将占据优势,且生长力明显提高,进而更新形成相对稳定的种群(胡尔查等,2013)。对样方内的疏齿木荷调查也证实,高度在 2—3 m的植株比例再次显著增加(图 1A)。因此从幼苗向幼树阶段的转变就成为假林线附近疏齿木荷更新的关键,未来需要通过适当的幼苗抚育,促使其持续更新(Tesfaye et al.,2002)。5 m以上的个体占比仅有0.25%,主要在于随着幼树逐渐成长为成熟林,其对资源的需求及对环境胁迫的敏感程度均在增加,加上逐渐衰老,导致大树存活的数量极为稀少(宋立奕等,2012)。
幼苗更新的早期受种源制约,而之后的更新过程则与生物和非生物要素均有关(邵佳怡等,2019)。对疏齿木荷种群的胸径研究发现,样方中92.6%的疏齿木荷胸径小于2.5 cm,极少有植株胸径大于 10.5 cm,这与尹爱国等(2004)和陈林等(2010)对粤北山顶矮林的研究结果较为一致,其乔木层大部分植株的胸径均在10 cm以下。这主要与疏齿木荷的生物学特性有关,其单株结实量大,且随风扩散能力强(Tang et al.,2020),可确保疏齿木荷幼苗更新充分,也可能是其成为假林线森林群落优势种的关键(宋立奕等,2013)。但随着幼苗生长,个体对资源的需求也不断增加,在资源有限的情况下,进而导致其个体数量随着胸径增加出现锐减(赵欣鑫等,2017),其中胸径小于等于2.5 cm 的植株密度达到 15417 plant·hm-2,而大于 2.5 cm的植株密度仅有1283 plant·hm-2。疏齿木荷的种群结构因而表现为基部较宽、顶部极窄的金字塔型结构,树木的更新和死亡率在短时间尺度上达到了平衡,种群增长性较低(宋立奕等,2012;Chhetri et al.,2015;吴语嫣等,2019)。
假林线所在的矮曲林生长受生境影响较大,其普遍低矮,且扭曲生长,因此基于胸径和树高的生物量估测模型能够更好地估算疏齿木荷的生物量(Poorter et al.,2012;Hunter et al.,2013)。本文的估算结果表明,疏齿木荷的地上生物量为 7.11 t·hm-2,其远低于付甜等(2014)对三峡库区八种亚热带森林乔木层地上生物量的估算结果,说明人类活动中止后,假林线所在的植物群落长势较差,树木的天然更新不容乐观。疏齿木荷的根系生物量占总生物量的比例为 21.87%,略大于陈章和等(1996)采用全挖法对黑石顶南亚热带常绿阔叶林地下部分生物量的测定结果(20.54%)。这主要与山顶土层浅薄和土壤贫瘠有关,疏齿木荷的幼苗一开始倾向于将更多地生物量分配到根系,以确保水分和养分的供应,但同时也削弱了其与相邻草灌木竞争空间和光照的能力,不利于其后续的更新(张文辉等,2008)。依据根冠比估算可知,本研究区疏齿木荷的总生物量为9.10 t·hm-2,也远低于Fang et al.(1998)对亚热带森林典型树种的总生物量估算结果,主要与当前疏齿木荷种群以幼苗为主,成熟林占比相对较少有关,其碳储存相对于同气候区偏低,碳汇潜力较大。未来若能对现有树木进行适当的人工抚育和科学管理,促进其幼苗向成熟林的演替,则有望增强这一地区的碳汇功能(胡海清等,2015)。Bastin et al.(2019)对全球森林恢复的评估中也指出,因早期退化而以稀疏灌丛或草地为主的亚热带山地可能是未来全球森林恢复的潜在区域之一,并有望缓解当前的气候变化。
4 结论
本文通过对广东石门台国家级自然保护区假林线群落的物种组成及乔木层优势种疏齿木荷的种群特征进行研究,结果表明:
(1)受山顶效应影响,假林线的群落结构简单、总体矮化,形成了典型的亚热带山顶常绿苔藓矮曲林,其乔灌草的优势种分别为疏齿木荷、五列木和野古草。
(2)疏齿木荷的高度分布表现为反“J”型,胸径以中小径级为主,其幼苗更新充分,但死亡率较高,大径级植株极少,形成了典型的底部较宽、顶部极窄的金字塔型结构,种群增长性较低,其中从幼苗到幼树的过渡成为研究区假林线附近疏齿木荷种群更新的关键。
(3)疏齿木荷的地上生物量和总生物量均低于同气候区的森林,其林分质量较差,未来需要通过适当的人工抚育,提高成熟林所占比重,从而增强这一地区的碳汇功能。
气候变暖背景下,不同物种的响应存在差异,进而会影响到假林线附近的物种组成及其种群特征。未来需要在亚热带山地选择典型样地,采用固定样方对于假林线群落的物种组成、种群特征和非生物要素等开展长期系统的定位观测,研究不同抚育措施下,假林线优势种疏齿木荷幼苗的更新潜力,这些都有助于理解亚热带山顶矮林的次生演替过程,并对准确评估其生态系统的服务价值也尤为重要。
