红砂植物种群空间分布与沙子粒级及地貌起伏之间的关系
2020-12-16刘红梅吕世杰任倩楠刘清泉刘丽英王玉芝周瑶
刘红梅,吕世杰,任倩楠,刘清泉,刘丽英,王玉芝,周瑶
1. 内蒙古自治区林业科学研究院,内蒙古 呼和浩特 010010;2. 内蒙古农业大学,内蒙古 呼和浩特 0100183;3. 赤峰市红山区棚户区改造管理办公室,内蒙古 赤峰 024000
红砂(Reaumuria soongorica)植物种群是草原化荒漠和典型荒漠区重要建群种或优势种之一,具有较强的生态可塑性和抗逆性,其耐干旱、耐贫瘠和较强的集沙能力等生物学特性保证其能够在生态脆弱区生殖繁衍,进而形成草原化荒漠和典型荒漠区特有的景观特征(刘家琼等,1982;马茂华等,1988;付贵全等,2016)。有研究表明,红砂有性繁殖的种子能在沙上良好发芽,而在沙间发芽率较低,在0—0.5 cm埋深发芽率最高(曾彦军等,2004;罗伟祥等,2007)。因此,沙层沙子组成比例有可能对其有性繁殖扩散产生影响,进而影响红砂植物种群空间分布状态。
红砂种群空间异质性明显,阳坡的红砂种群呈现聚集分布,阴坡则多呈现均匀分布;且该研究认为,幼龄红砂多呈聚集性分布,随着年龄增大,空间分布的聚集程度减弱(周资行等,2011)。也有研究表明(李小刚等,2016),红砂在兰州是南北山上的空间分布均匀度较大,而山下空间分布将聚集程度增加。这些聚集与均匀空间分布(或随机分布)特点会影响红砂植物种群空间分布的异质性、斑块性以及扩散过程,反映的是种群扩散、群落演替及生物与环境因子相互作用的生态学过程和及其适应过程(夏素娟等,2018)。红砂根系受沙层水分影响具有较强的形态可塑性(孙百生等,2018),而巴丹吉林沙漠在0.20—0.30 m存在湿沙层,且上部干沙层由于沙子粗度较大,导致下沙层的水分蒸发量下降(赵景波等,2011),使得红砂植物种群能够在此定植扩散成为可能。所以,探讨红砂空间分布与表层沙子粒级及地貌起伏之间的关系,不仅可以全面认识巴丹吉林沙漠红砂种群自然分布的特点和规律,也可初步揭示其空间分布状态形成的演变过程。
巴丹吉林沙漠东缘一年四季干旱少雨,冬春季风沙大(刘红梅等,2015;王猛等,2016),属于典型的荒漠区,红砂作为该区域的优势种群,其空间分布不仅受自身生物特性影响,也受该区域合成风向、地貌起伏条件等因素调控(Keitt,2000;刘冰等,2007);且该地区人为干扰因素较少,保留了红砂植物种群受环境条件和自身生物学特性影响下的空间分布特点和变化规律。开展红砂植物种群空间分布与沙子粒级及地貌起伏之间的相关研究,不仅可为掌握红砂植物种群自然分布提供理论依据和数据支撑,也可为典型荒漠区植被建植以及生态环境保护提供理论指导和科学依据。
1 试验地概况与研究方法
1.1 试验地概况
试验于阿拉善右旗塔木素苏木地区“塔木素野生肉苁蓉及梭梭产籽基地”进行,地理位置为103°25.162′—103°27.546′E,40°33.059′—40°35.531′N,行政区划属于塔木素苏木格勒图嘎查的朝恒扎干。试验地由于地处内陆高原,降水量少,蒸发量大,日温差较大,为典型的温带干旱荒漠性气候。年平均降水量105.3 mm,主要集中在7—8月,年有效降雨 1—2次;年均温 8.5 ℃,绝对最高温接近43 ℃,年蒸发量达到3179.7 mm。无霜期150—165 d,年均风速4 m·s-1,冬春季以西北风为主,8月东北风为主,其他月份西风占主导地位(陈红宝,2011;张翼飞等,2013;王礼恒等,2019),大风之时,易出现沙暴。自然植被稀少,种属贫乏、覆盖率低,地表裸露。植物以旱生和超旱生灌木,半灌木为主,主要有梭梭(Haloxylon ammodendron)、白刺(Nitraria tangutorum)、红砂(Reaumuria songarica),稀有霸王(Zygophyllum xanthoxylon)、沙拐枣(Calligonum mongolicum)和珍珠(Salsola passerina)出现。草本植物很少,尤以一年生“夏雨型”草本植物为主;主要为倒披针叶虫实(Corispermum lehmannianum)、偶有驼蹄瓣(Zygophyllum fabago)、雾冰藜(Bassia dasyphylla)和砂蓝刺头(Echinops gmelini)等植物出现。这些植物具有耐干旱、耐高温、抗风沙的生态及生物学特性(刘红梅等,2015;王猛等,2016)。
1.2 试验设计及数据来源
试验采用大样地调查法:选择围栏内的 1600 m×1200 m大样地为固定试验样地,以西南方向为绝对坐标的原点(0,0),横向为X轴(由西到东),纵向为Y轴(由南到北),每隔200 m设为一条样线,在样线上调查每一株植物,记录植物名称、相对坐标,并测定红砂冠幅直径。红砂植物种群在空间上的相对位置见图1。图1中每一个“+”代表一株红砂。调查共发现2458株植物,分为6个植物种群,其中红砂植物种群886株,占比高达36.05%,仅次于占比44.22%的梭梭种群。

图1 红砂种群个体空间分布位置Fig. 1 Spatial distribution of R. soongorica
在图1顶点和样线交点位置,采用GPS测定该点的海拔高度作为地貌起伏特点的基础数据,然后在该点附近随机采集3个点0—2 cm沙层沙子,面积为10 cm×10 cm,带回实验室混合均匀后,采用激光粒度分析仪(Mastersizer 3000)进行测定,测量范围为0.1—2000 μm,对应美国1991年制定的8级标准(邓继峰等,2017),划分为粗沙(d≥0.5 mm)、中沙(0.25 mm<d≤0.5 mm)和细沙(d≤0.25 mm)(周景山,2013)。
1.3 数据分析
根据图1数据采样方式,采用200 m×200 m大小方格将取样空间进行整合,计算方格内的植物株数(折算成交点和顶点上的密度,plant·200 m-1),高度(cm)、冠幅直径(cm),形成的数据点与沙子取样样点直接对应(n=63)。对红砂植物种群密度、高度、冠幅直径、细沙、中沙、粗沙以及地貌起伏(海拔高度)进行半方差函数分析,在GS+9.0中设置有效滞后距离为1000 m,最小响应距离200 m,采用 Spherical、Exponential、Gaussian 和 Linear模型进行拟合,并记录最优拟合结果的相关参数(刘红梅,2011a,2011b)。
在Surfer 12.5中绘制红砂植物种群密度、高度、冠幅直径、细沙、中沙、粗沙以及地貌起伏的等高线图和集水径流图,分析各指标空间分布的一致性、斑块性以及空间分布趋势(刘红梅等,2015)。
在 SAS 9.2中,采用典型相关分析(调用CANCORR过程)探讨红砂(密度、高度和冠幅直径)与沙子粒级(细沙、中沙和粗沙)之间的相关性,根据获得的典型变量分别与红砂植物种群密度、高度、冠幅直径、细沙、中沙、粗沙以及地貌起伏等进行Pearson相关分析(调用CORR过程)。
2 结果分析
2.1 红砂植物种群数量特征空间分布特点
红砂植物种群密度、高度和冠幅直径的最优拟合模型分别为球形模型(Spherical)、指数模型(Exponential)和指数模型(Exponential),模型的决定系数分别为 1.000、0.785和 0.796;因此采用最优拟合模型能够阐释各指标的空间分布状况(表1)。结构比C/(C+C0)显示,红砂植物种群各指标空间变异处于中等程度,即其空间分布受结构性因素和随机性因素共同控制。密度、高度和冠幅直径自相关尺度分别为931、1554、2213 m,表明其空间分布的斑块性在依次增大。根据分形维数可知,红砂种群密度空间分布的随机性大于高度和冠幅直径,分形维数分别为 1.773、1.851和 1.872。同理分析海拔高度可知,其空间变化的一致性较高,异质性较低,且异质性低于红砂植物种群密度、高度和冠幅直径;空间自相关尺度725 m,空间分布的斑块性小于红砂种群任一指标;分形维数显示其空间分布受随机因素影响较大。
红砂植物种群密度整体呈西北向东南逐渐增加的变化趋势(图2a,等高线),形成的集水盆存在6个,集水径流3条;对应海拔高度(图2b),红砂植物种群密度空间分布趋势与海拔高度(图2d)空间分布趋势恰好相反;表明海拔高度对于种群密度空间分布存在负相关关系。红砂高度空间分布的差异相对较小;在东北区域存在面积较大且冠幅直径(图2c)较大的分布区(东南分布区面积较小),所以红砂的密度、高度和冠幅直径尽管同属于一个种群,但因指标不同形成的空间分布形式差异较大(等高线变化、集水盆位置以及集水径流分布状况),同时也发现红砂的密度、高度和冠幅直径的集水径流主要分布在海拔高度集水盆周边,由此可以推断,海拔高度形成的集水盆为沙丘高度形成的沙丘或沙带所致,红砂植物种群空间分布状态受试验区内沙丘或沙带形状影响。

表1 红砂植物种群及海拔高度空间变化特征Table 1 Spatial distribution characteristics of R.soongorica population and altitude
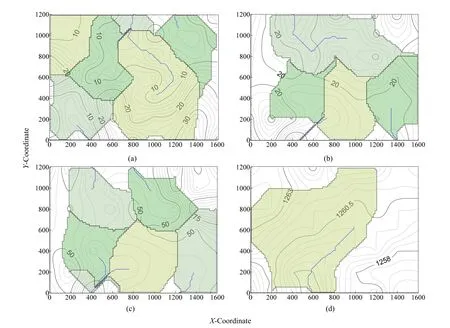
图2 红砂植物种群及海拔高度的等高线图及集水径流图Fig. 2 Contour map and catchment runoff map of R. soongorica population and altitude
2.2 表层沙子粒级空间分布特点
对细沙、中沙、粗沙和海拔高度的最优拟合模型分别为指数模型、指数模型、指数模型和高斯模型(表 2),其中海拔高度的拟合效果最好,决定系数为0.990,而中沙的拟合效果最弱,仅为0.643。由此可见,粗沙空间分布反映的原数据信息较多,而中沙反映的原数据信息较少一些,但这并不影响细沙、中沙和粗沙的空间分布整体趋势。结构比显示,细沙和粗沙空间异质性比较大,其结构比分别为0.566和0.589,但是二者的自相关尺度差别较大,粗沙的自相关尺度是细沙的6倍多,分别为1499 m和246 m。中沙空间异质性较弱,结构比高达0.865,自相关尺度较细沙小,为121 m;说明中沙空间分布的斑块性更为明显。分形维数表征的空间分布状态难以与结构比表征信息相一致。
细沙空间分布在东南区域占比较高(图 3a,78%),集水径流斑块(集水盆)存在两个,积水径流分别经由南北两向流出,其中这两条径流存在很大尺度垂直于合成风向(西北→东南)。中沙在东南区域占比较少(图3b),在北方区域占比较高,形成集水盆3个,集水径流3条,其中东南区域和西面区域的集水径流较长。粗沙在试验区的北方占比较高(图3c),形成集水盆3个,集水径流3条,其中西南和东北两个区域的集水径流较长。由此可见,细沙、中沙和粗沙空间分布状况各不相同,且与海拔高度(图3d)空间分布状态也存在较大差别。

表2 不同粒级沙子及海拔高度空间变化特征Table 2 Spatial distribution characteristics of sand with different particle size and altitude

图3 不同粒级沙子和海拔高度的等高线图及集水径流图Fig. 3 Contour map and catchment runoff map of sand with different particle size and altitude
2.3 红砂与表层沙子及地貌起伏之间的相互关系
红砂空间分布与沙子空间分布存在显著的相关性(r=0.4244,图 4),这种相关性包含了红砂密度、高度和冠幅信息,也包含了沙子不同粒级占比情况。在图4中可以看出,红砂密度与细沙呈正相关性(在相关路“密度→红砂→沙子←细沙”的相关系数均为正值;如果相关系数个数为奇数则为负相关,为偶数则为正相关),而与中沙和粗沙呈负相关。红砂的高度与细沙呈负相关,而与中沙和粗砂呈正相关。红砂的冠幅不受沙子粒径影响,整体与沙子存在负相关趋势。海拔高度能够与红砂的空间分布存在相关性,但不能影响沙子的空间分布状态;且海拔越高,红砂的密度越小,但红砂的高度较高。
3 讨论
3.1 红砂的空间分布特点及其影响因素
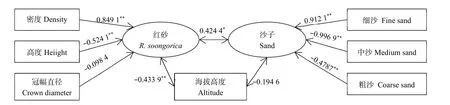
图4 红砂种群、不同粒径沙子及海拔高度之间的相互关系Fig. 4 Relationship among R. soongorica population, sand with different particle size and altitude
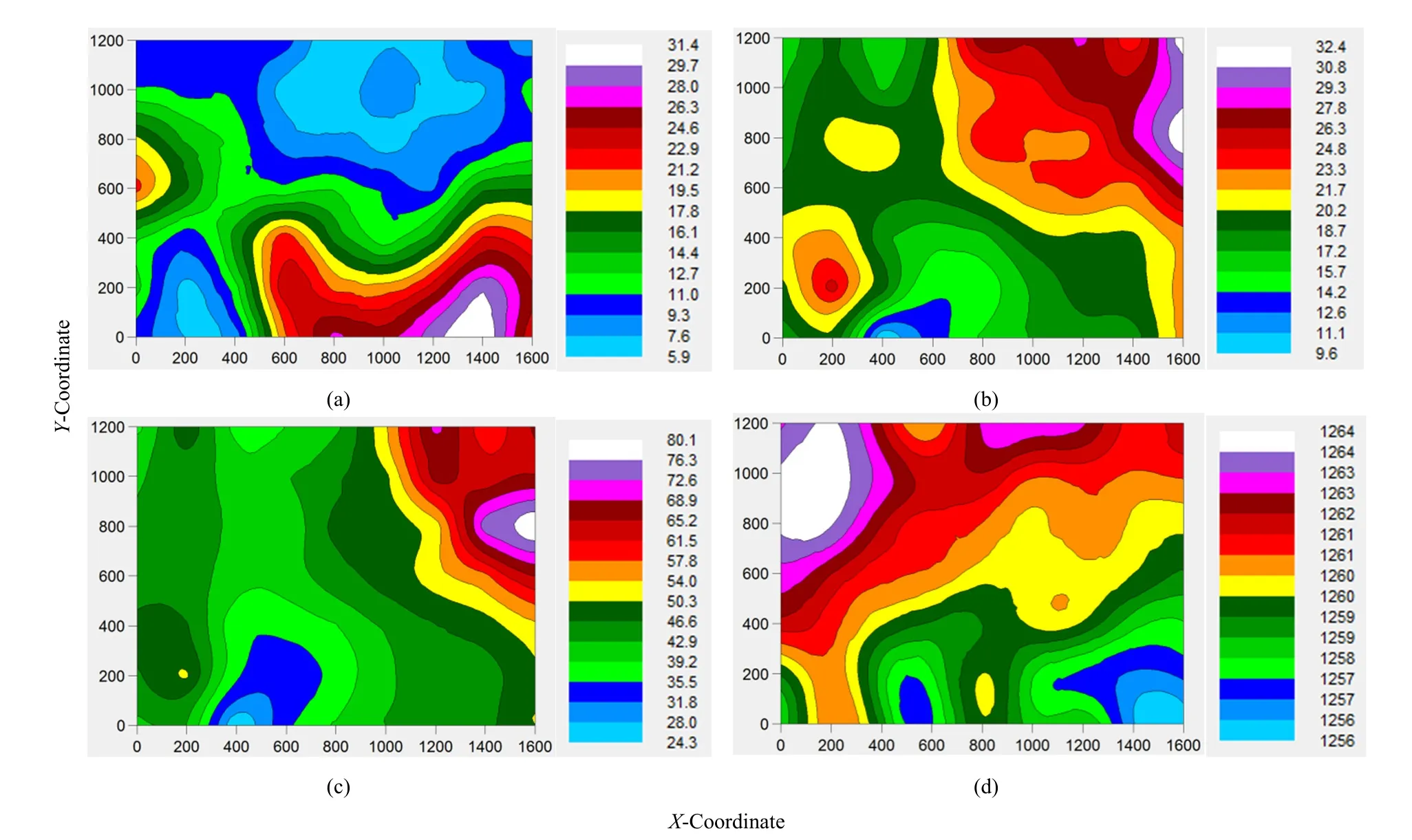
图5 红砂不同指标和地貌起伏空间变化色带图Fig. 5 The spatial distribution color band map of R. soongorica different indexes and terrain
试验区地貌起伏特点是由于该区域冬春季盛行西北风,这是形成沙丘或沙带的主要影响风向,辅以东北风和西风形成图 5d沙丘或沙带空间分布形式,且这一分布状况垂直于合成风向(刘冰等,2007;刘红梅等,2015)。红砂的密度(图5a)空间分布区域在东南,而株高(图 5b)和冠幅(图5c)的空间分布却在东北方向,但是海拔高度却在西北方向最高。这表明红砂的密度、株高、冠幅以及地貌起伏空间变化的决定因素不同,但受结构性因素和随机因素影响程度一致(表 1结构比,Cambardella et al.,1994)。红砂的密度分布较高区域位于主风向(西北风)东南方,此区域海拔较低,细沙经风吹和沙带阻挡容易在这里聚集,这表明红砂萌发需要风沙相对稳定的环境条件,且细沙相对中沙和粗砂更容易锁住水分(赵其文,2006;蔡永坤等,2014),红砂种子被毛也使得其萌发过程有利于利用水分条件(罗伟祥等,2007),使得红砂植物种群在此区域具有较高的发芽率,导致此区域密度较高。然而,红砂生长发育适应的环境条件与种子萌发条件存在差异,其在东北方向表现较高,因此根据生长发育状况,可以推断红砂入侵研究区沙带的方向为东北,其种子成熟,受合成风向影响发生位移,聚集在海拔较低、水分条件相对较好的区域,并形成高密度聚集分布区;这与罗伟祥等(2007)的研究结果一致。从入侵方向看,刘红梅等(2015)研究认为梭梭由西南入侵沙带,经合成风向影响在主风向前端(沙带东南向)建植,起到防风固沙作用。本研究红砂由东北方向入侵,经合成风向综合影响,也在主风向前端(沙带东南向)形成高密度分布区,阻止沙带迁移,进而起到防风固沙作用。所以,此区域不同植物种群空间分布状态对沙带的影响有不同的生态学过程,受自身生物学特性和环境条件综合作用。
3.2 不同粒径沙子空间分布状态及其影响因素
研究区细沙空间分布状态正和此区域常年风向相似,西北风为此区域主风向,西南风和东北风在此区域也经常出现,导致细沙(图6a)占比70%—77%的空间分布与该区常年风相重合,也与该地区较高海拔区域大面积重合(图6d),所以细沙空间分布状态受地貌因素和风向因素双重控制。然而,细沙占比最高分布区却在西方和东南区域,所以细沙占比最高区域也受地貌因素和风向因素双重控制。刘红梅等(2015)研究该区域地貌起伏变化特点时认为,合成风向是形成该研究区域沙丘或沙带的主要影响因素,所以在该区域合成风向是细沙空间分布的主要因素,而地貌起伏是该区域细沙空间分布的次要因素,二者综合作用导致最高区域分布区主要呈现出垂直于主风向(刘冰等,2007)。在细沙分布区存在较为密集的红砂植株,其具有阻挡沙尘降落的功能,使得细沙占比进一步增加。中沙(图6b)和粗沙(图6c)空间分布主要集中在北方,且与地貌次高区域(横坐标1000,纵坐标1200附近)重合,说明东北风和西北风出现的频度和强度较高,而主风向西北风形成研究区最高区域位于西北方;细沙较中沙和粗沙容易被风吹动,合成风向导致该区域细沙被吹走的比例较大,同时受东北风和西北风叠加作用,导致该区域海拔较高且中沙和粗砂占比较大。因此,该研究区域不同粒级沙子空间分布形成的分布特点,是由该研究区合成风向、地貌特点和植被状况共同决定。
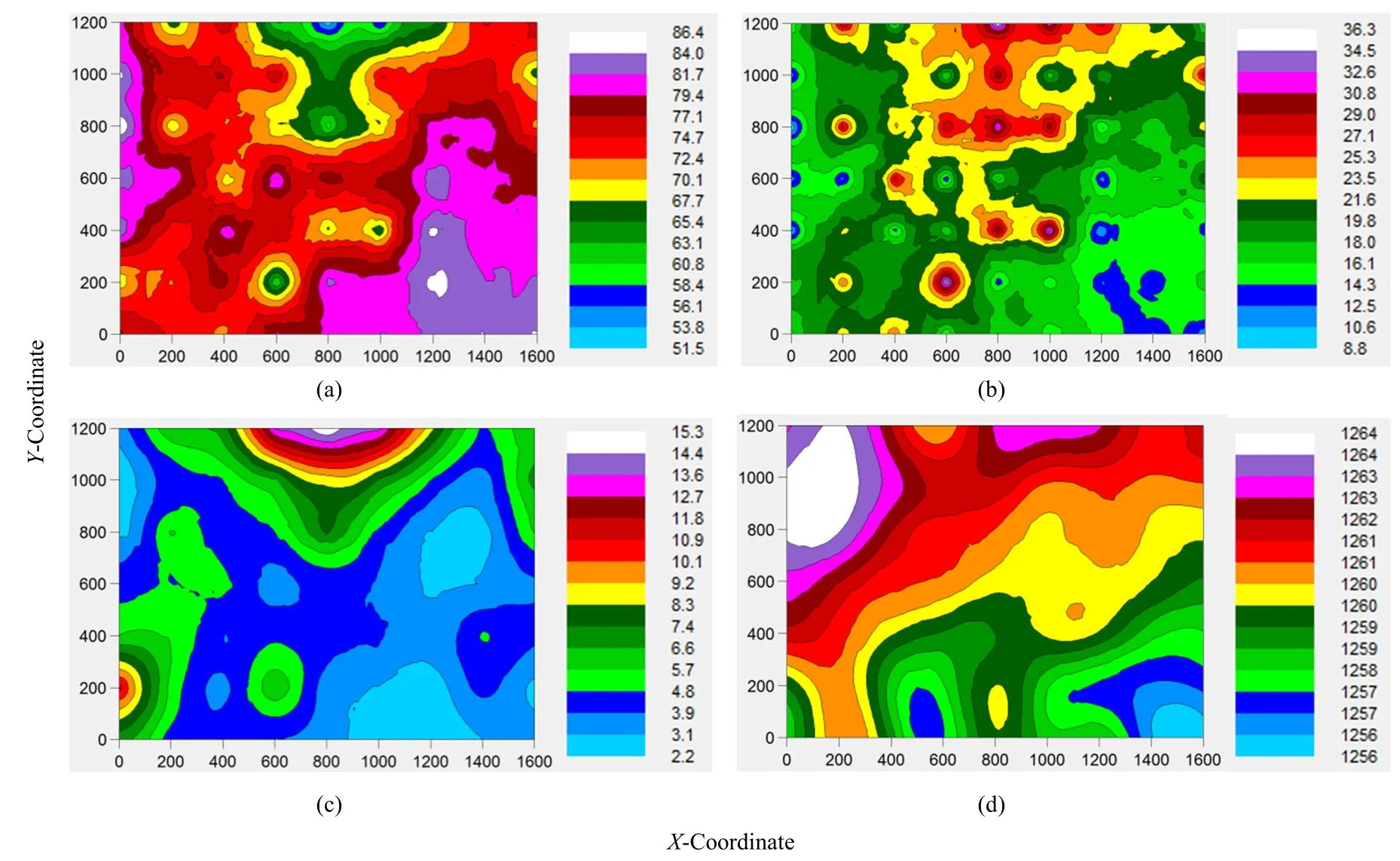
图6 不同粒级沙子和地貌起伏空间变化色带图Fig. 6 The spatial distribution color band map of sand with different particle size and altitude
3.3 红砂的空间分布与沙子粒径和地貌起伏的关系
典型相关显示,红砂植物种群密度空间分布与细沙呈正相关,在空间分布图(图 6)上显示,高比例细沙分布区与高密度红砂分布区重合。首先,红砂萌发和幼苗生长需要相对较高的水分条件,而细沙能够锁住较多的水分,为红砂提供必要营养物质和水分供给(赵其文,2006);其次,该区域两边高,受季节风向影响细沙能够对红砂种子进行轻度覆沙,为红砂种子萌发和幼苗生长提供必要条件。当红砂能够定植之后,由于其自身具有富集流沙的能力,形成大的灌丛,为自身的无性繁殖创造了条件(曾彦军等,2002)。所以在东北区域红砂株高和平均冠幅比较好的原因是种群个体适应环境的结果,也是因为存在流沙掩埋的结果。综合来看,红砂空间分布与沙子粒径直接相关,但是不同粒径影响的红砂指标并不一致;这与红砂定植前后对环境的要求不同所致。该研究区域风向作用形成现有地貌特征,地貌特征与风向共同作用决定沙子粒级空间分布特点,进而决定红砂植物种群空间分布状态。同时我们发现,结构比和分形维数在沙子粒级上和海拔高度上难以一致,原因是在研究决定性因素上存在差异,突出重点不同,半方差函数主要研究不同尺度上的空间结构,该分形维数分析的是没有尺度的空间结构(吕世杰等,2014)。
4 结论
红砂植物种群的密度、高度和冠幅空间异质性处于中等强度(结构比在25%—75%),斑块尺度依次增大,分别为931、1554、2213 m。中沙斑块尺度较小,仅为121 m;粗沙斑块尺度是细沙的6倍多,分别为1499 m和246 m,二者空间异质程度高于中沙。红砂植物种群密度空间分布与细沙空间分布紧密相关(P<0.050),且高密度分布区与高比例细沙分布区重合。受红砂植物学特征和生物学特性影响,红砂在定植前后对环境的要求不同。该研究区域风向作用形成现有地貌起伏特征,地貌起伏特征与风向共同作用决定沙子粒级空间分布特点,进而与红砂植物种群生物学特征相互作用形成其空间分布现状。
