圆纹广翅蜡蝉产卵规律及温度对越冬卵发育的影响
2020-12-15金银利马全朝张绍杰尹健史洪中耿书宝
金银利,马全朝,张绍杰,尹健,史洪中,耿书宝*
圆纹广翅蜡蝉产卵规律及温度对越冬卵发育的影响
金银利1,2,马全朝1,张绍杰1,尹健1,史洪中1,耿书宝1*
1. 信阳农林学院,河南 信阳 464000;2. 河南省豫南茶树资源综合开发重点实验室,河南 信阳 464000
为了阐明圆纹广翅蜡蝉(Walker)产卵对茶树的为害规律及温度对其越冬卵发育的影响,通过五点取样法,系统调查了圆纹广翅蜡蝉产卵对茶树的为害情况,并研究了16、19、22、25、28℃恒温对圆纹广翅蜡蝉越冬卵发育的影响。结果表明,圆纹广翅蜡蝉成虫喜欢在直径为(0.23±0.04)cm的茶树幼嫩枝条上产卵,产卵刻痕在枝条上的为害长度为(1.36±0.59)cm。在茶园周围有较多寄主林木且管理较为粗放的马鞍山茶叶试验场阶梯茶园内,茶树枝条的受害率为13.35%,显著高于其他茶园。此外,越冬卵的发育历期,随着温度的升高逐渐缩短,28℃处理的越冬卵发育历期为(28.75±7.03)d,若虫初孵时间较早,孵化率较高。温度与圆纹广翅蜡蝉越冬卵发育速率关系满足方程=0.001 9-0.022,越冬卵的发育起点温度为11.58℃,有效积温为526.32 d·℃。本研究对信阳茶区圆纹广翅蜡蝉越冬种群发生期的预测预报和防控策略的制定具有重要的指导意义。
圆纹广翅蜡蝉;产卵规律;茶树;发育历期;发育起点温度;有效积温
圆纹广翅蜡蝉(Walker)又名圆纹宽广蜡蝉,隶属于半翅目(Hemiptera)广翅蜡蝉科(Ricaniidae),植食性,是农林业的重要害虫[1-6]。据报道,该虫主要分布于我国的福建、贵州、广西、湖南和湖北等地,寄主植物范围十分广泛,以阔叶园林植物为主,主要为害小叶女贞、樟树、桂花、冬青、万年青、日本樱花、迎春花、杜仲、油茶、茶、柑橘、刺槐、肉桂和金银花等多种植物[7-12]。该虫的成虫和若虫均能刺吸取食寄主植物叶片和嫩枝的汁液,若虫常群集在叶片或嫩枝梢上为害,轻则引起枝梢和叶片营养不良,叶片褪绿发黄或蜷缩畸形,重则导致枝叶枯死,严重影响树势[13-15]。
目前,对于圆纹广翅蜡蝉的研究主要集中在其形态特征识别、生物学特性、生态学及防治方法等方面[7-15]。1995年,谭济才[7]对湖南省茶园蜡蝉种类调查研究发现,共有12种蜡蝉危害茶树,其中圆纹广翅蜡蝉为新记录种,但之后,并没有该虫在茶树上发生为害规律的相关研究。据报道,圆纹广翅蜡蝉在园林植物上一年发生1代,一生经过卵、若虫和成虫3个虫态[8-10]。该虫在园林植物嫩枝梢组织中产卵越冬,初产卵刻痕外被一层棉絮状白色蜡丝覆盖,影响枝条生长,产卵部位以上枝条易折断枯死,甚至整枝枯死,严重削弱树势[11-15]。翌年,初孵若虫直接在寄主植物越冬刻痕附近的嫩叶背面和嫩枝梢上聚集为害。
昆虫越冬卵的发育与春季的温度、湿度和光照等气候因素有着密不可分的关系,其中,温度是决定性因素,对昆虫越冬卵的发育历期、孵化率、发育起点温度和有效积温等具有十分重要的影响[16-20]。为此,本文在调查圆纹广翅蜡蝉越冬种群对茶树为害规律的基础上,研究了不同温度对其越冬卵发育历期和孵化率的影响,以期得到越冬卵的发育起点温度和有效积温,旨在为信阳茶区圆纹广翅蜡蝉发生期的预测预报和防控策略的制定提供理论参考。
1 材料与方法
1.1 调查茶园概况
2019年2月下旬,在信阳茶区南湾湖沿线茶园,调查圆纹广翅蜡蝉成虫在茶树[(L.) O. Ktze]上的越冬产卵部位以及产卵对茶树枝条的为害率。调查茶园类型和位置如下:
(1)阶梯茶园:马鞍山茶叶试验基地(32°06′N,114°04′E)、十三里桥小庙村(32°01′N,113°59′E)、浉河港马到岭(32°01′N,113°54′E)和董家河清塘村(32°03′N,113°51′E)。
(2)平地茶园:董家河陈湾村(32°08′N,113°55′E)、浉河港郝家冲村(32°01′N,113°54′E)、浉河港白庙村(31°59′N,113°51′E)和浉河港夏家冲村(32°03′N,113°53′E)。
茶园概况见表1。马鞍山茶叶试验基地和董家河陈湾村茶园管理相对粗放,茶树修剪不整齐,杂草丛生,茶园地头或间作种植林木较多。马鞍山茶叶试验场临近蚕桑试验站,种植有大量桑树,茶园地头还有构树、樟树和花椒树等林木,茶园环境隐蔽。董家河陈湾村茶园在水土保持工程附近,周围种植观赏林木较多,茶树行间间作有桂花树树苗(直径:2~3 cm)。
1.2 圆纹广翅蜡蝉在茶树上的越冬为害规律
于2019年2月下旬,在表1所示的8个茶园中,调查圆纹广翅蜡蝉成虫产卵对茶树枝条的为害率(5)点取样,每点随机选取100个茶枝),并将受害枝条带回实验室,置于4℃冰箱冷藏备用。检测统计圆纹广翅蜡蝉成虫在茶树枝条上的产卵部位,利用游标卡尺测量受害茶枝直径、产卵刻痕在茶枝上的为害长度以及茶枝顶端刻痕到芽梢的距离。为了能够更清晰地观察茶枝上的产卵刻槽,利用尖头镊子将具有产卵刻痕茶枝的表皮慢慢剥开,仔细剔除卵与卵之间的其他覆盖物,使用体视显微镜带屏一体机(型号:SRZ-7045A和3610,上海光学仪器厂)进行镜检并拍照。
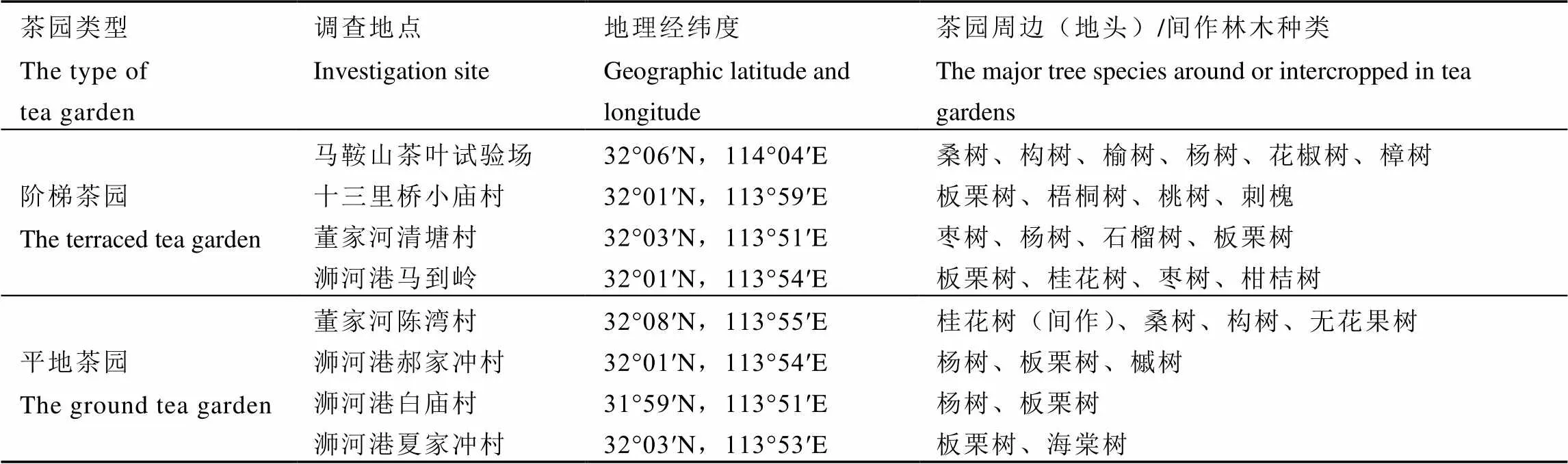
表1 调查茶园概况
1.3 温度对圆纹广翅蜡蝉越冬卵发育的影响
2019年3月初,选取从茶园采回的带有越冬产卵刻痕的枝条,单枝置于50 mL塑料管内,加入约30 mL水,之后用脱脂棉浸入管内并包裹茶枝以保证茶枝接触水而存活,分别置于信阳农林学院昆虫实验室的智能人工气候箱(型号:RTOP-310Y,浙江托普云农科技股份有限公司生产)内,设置5个温度,分别为16℃、19℃、22℃、25℃、28℃,光周期均为L∶D=14∶10,相对湿度RH均为(75±5)%。每天观察圆纹广翅蜡蝉越冬卵的孵化情况,记录若虫孵化时间和数量。每处理3个重复,每重复不少于3个具卵刻痕茶枝。
1.4 数据处理与统计分析
采用Microsoft Excel 2010、SPSS Statistics 16.0(IBM,Chicago,IL,USA)和R语言软件(R version 3.2.3)进行数据统计分析。圆纹广翅蜡蝉成虫产卵对茶枝的为害率和不同温度条件下越冬卵的孵化率采用贝叶斯(Bayesian)方法计算并比较差异[21]。成虫在茶枝顶端的产卵刻痕至芽梢的距离、选择产卵茶枝直径和产卵刻痕在茶枝上为害长度的频数统计采用频数分析(Frequencies)。不同温度下圆纹广翅蜡蝉越冬卵发育历期差异显著性分析采用Kruskal-Wallis tests,多重比较采用Duncan检验。温度()与发育速率()的关系利用线性(=a+b)拟合,得出越冬卵的发育起点温度(=–b/a)和有效积温(=1/a)。
2 结果与分析
2.1 圆纹广翅蜡蝉在茶树上的越冬为害规律
2.1.1 圆纹广翅蜡蝉在茶树上的越冬部位
茶园调查发现,圆纹广翅蜡蝉成虫喜欢在茶树幼嫩枝条组织上产卵越冬,有的产在接近茶树芽梢的部位,其产卵刻痕外面覆盖一层白色的棉絮状蜡丝。去掉白色蜡丝,可见产卵刻痕是单个地环刻在茶树枝条上的,外露木质丝,为害严重的可致枝叶干枯(图1)。
在茶枝顶端的产卵刻痕到芽梢的平均距离为(3.99±2.28)cm(N=78),距离小于1.00 cm的出现频数为5,距离在1.00~6.00 cm的出现总频数为59,但均小于11.00 cm(图2)。
对带有越冬产卵刻痕的茶树枝条直径进行频数分析,发现成虫选择产卵的枝条直径平均为(0.23±0.04)cm(N=78),主要集中在0.18~0.25 cm范围内,频数为50,占总数的64.10%,但受害茶枝直径均小于0.40 cm(图3)。
2.1.2 圆纹广翅蜡蝉成虫产卵为害茶树规律
在南湾湖沿线8个茶园调查发现,圆纹广翅蜡蝉成虫产卵对周围有较多林木种植且管理较为粗放的马鞍山茶叶试验场阶梯茶园内茶树枝条的为害率最高,为13.35%,显著高于其他茶园(<0.05),其次是水土保持工程附近有较多观赏林木种植且茶树行间间作桂花树苗的董家河陈湾村茶园,枝条受害率为2.19%,与其他茶园的枝条受害率差异显著(<0.05)(表2)。这一现象说明了,管理粗放且种植有樟树等寄主林木的阶梯茶园,更容易受到圆纹广翅蜡蝉成虫产卵的为害。

注:a:茶枝上有白色蜡丝覆盖的危害状;b:干枯茶枝上的产卵刻痕;c:接近茶树芽梢的产卵刻痕
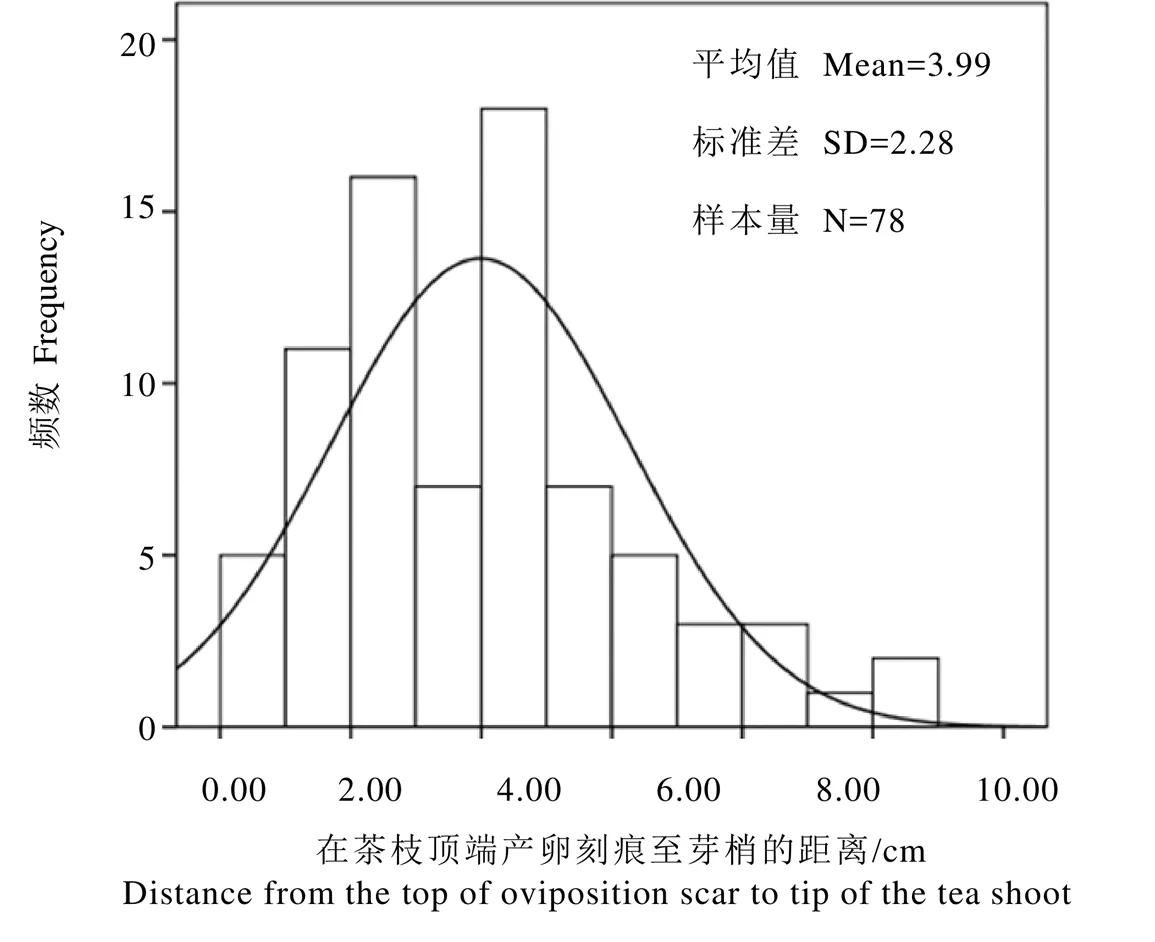
图2 圆纹广翅蜡蝉在茶枝顶端产卵刻痕至芽梢距离的频数统计
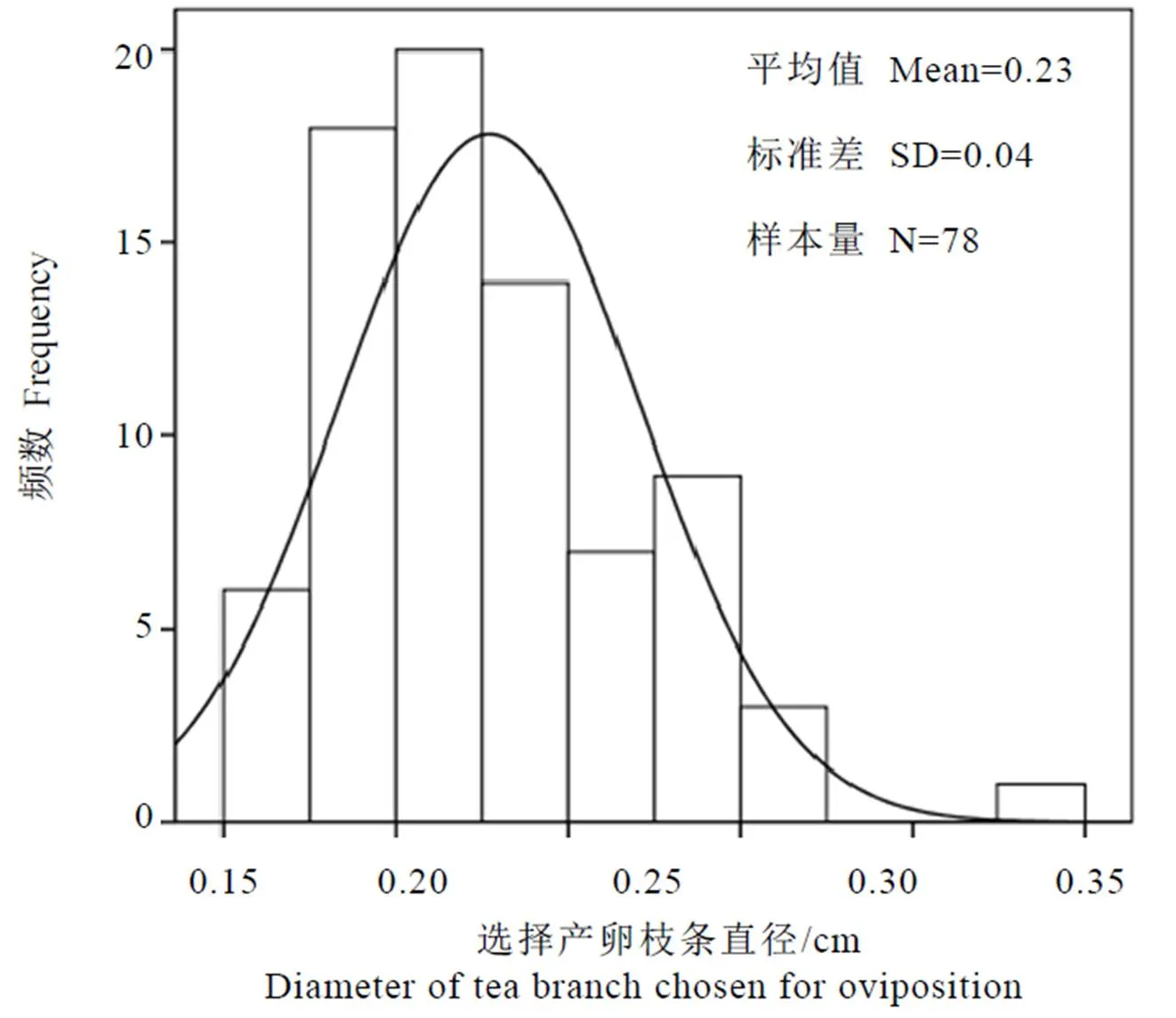
图3 圆纹广翅蜡蝉成虫选择产卵茶枝直径的频数统计
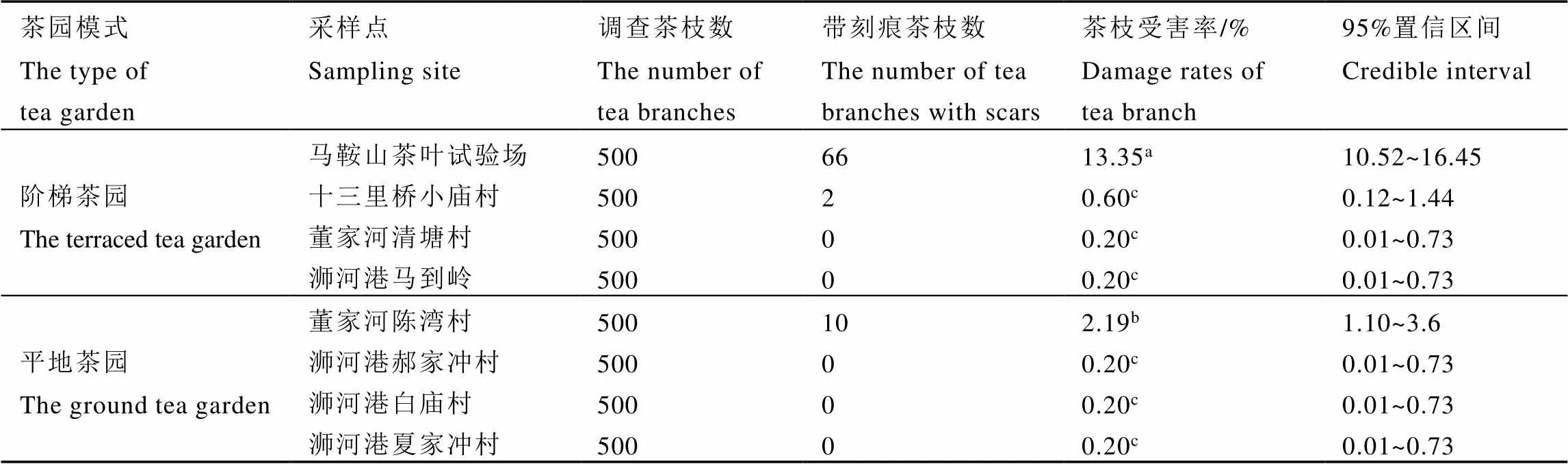
表2 圆纹广翅蜡蝉产卵刻痕对不同茶园茶枝的为害率
注:同列具有相同小写字母者表示在0.05水平上差异不显著
Note: Values followed by the same lowercase letters are not significantly different at the probability of 0.05
对圆纹广翅蜡蝉产卵刻痕在茶树枝条上的为害长度进行频数分析,发现其为害长度平均值为(1.36±0.59)cm(N=78),主要集中在0.50~2.25 cm,频数为71,占总数的91.03%(图4)。用尖头镊子轻轻剥开受害枝条表皮镜检,发现成虫产卵刻痕单个地环刻在茶树枝条组织内,越冬卵也是单个镶嵌排列在产卵刻槽内,外露木质丝(图5)。
2.2 温度对圆纹广翅蜡蝉越冬卵发育的影响
2.2.1 对越冬卵发育历期的影响
由图6可看出,22℃和25℃下,圆纹广翅蜡蝉越冬卵的发育历期分别为(51.31±13.98)d和(47.43±8.41)d,两者之间差异不显著(>0.05),但均与其他处理温度下的越冬卵发育历期差异显著(<0.05)。越冬卵在16℃、19℃和28℃下的发育历期分别为(107.97±5.39)d、(66.90±5.35)d和(28.75±7.03)d,三者之间两两差异显著(<0.05),而且,随温度的升高,越冬卵的发育历期逐渐缩短(图6)。
2.2.2 对越冬卵孵化率的影响
由图7可以看出,圆纹广翅蜡蝉越冬卵在16℃、19℃、22℃、25℃和28℃条件下的若虫初孵时间分别是在处理后的第98、55、21、31 d和21 d,除了22℃外,若虫初孵时间是随着温度的升高而缩短的。研究还发现,越冬卵累计孵化率在处理后116、79、67、61 d和53 d分别达到26.79%、34.25%、36.11%、51.75%和66.89%,较低温度的累计孵化率较低,但随着温度的升高,累计孵化率逐渐增加。
采用贝叶斯方法计算并比较分析,发现越冬卵在25℃和28℃条件下的孵化率差异显著(<0.05),且均与其他3个温度下的孵化率差异显著(<0.05),但16℃、19℃和22℃下的越冬卵孵化率无显著差异(>0.05)(表3)。这说明了越冬卵在28℃条件下的若虫初孵时间较早,孵化率较高。
2.2.3 圆纹广翅蜡蝉越冬卵的发育起点温度和有效积温
根据圆纹广翅蜡蝉越冬卵在16、19、22、25、28℃条件的发育历期结果,计算得出了该虫的加权平均发育历期(N)及发育速率(1/N)。结果表明,不同温度()与越冬卵发育速率()的关系满足方程:=0.001 9-0.022(R=0.91;(1, 3)=28.98;=0.012)(图8)。根据有效积温=1/a,发育起点温度=–b/a,得出圆纹广翅蜡蝉越冬卵的发育起点温度=11.58℃,有效积温K=526.32 d·℃。
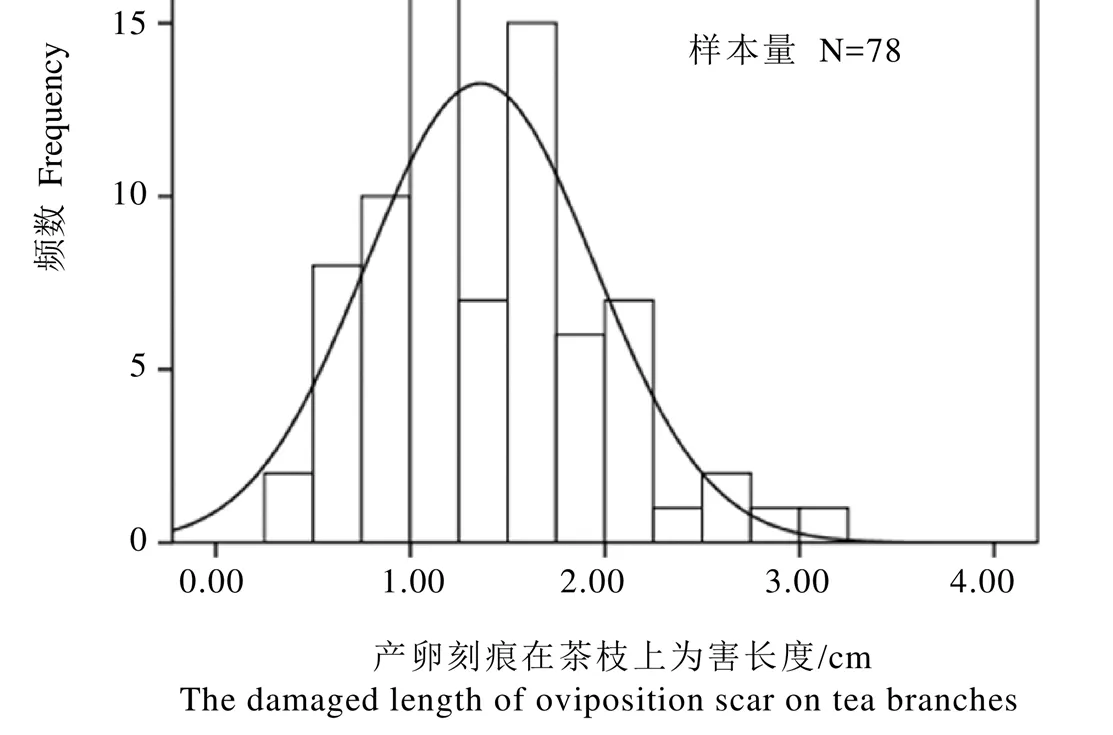
图4 圆纹广翅蜡蝉产卵刻痕在茶枝上为害长度的频数统计
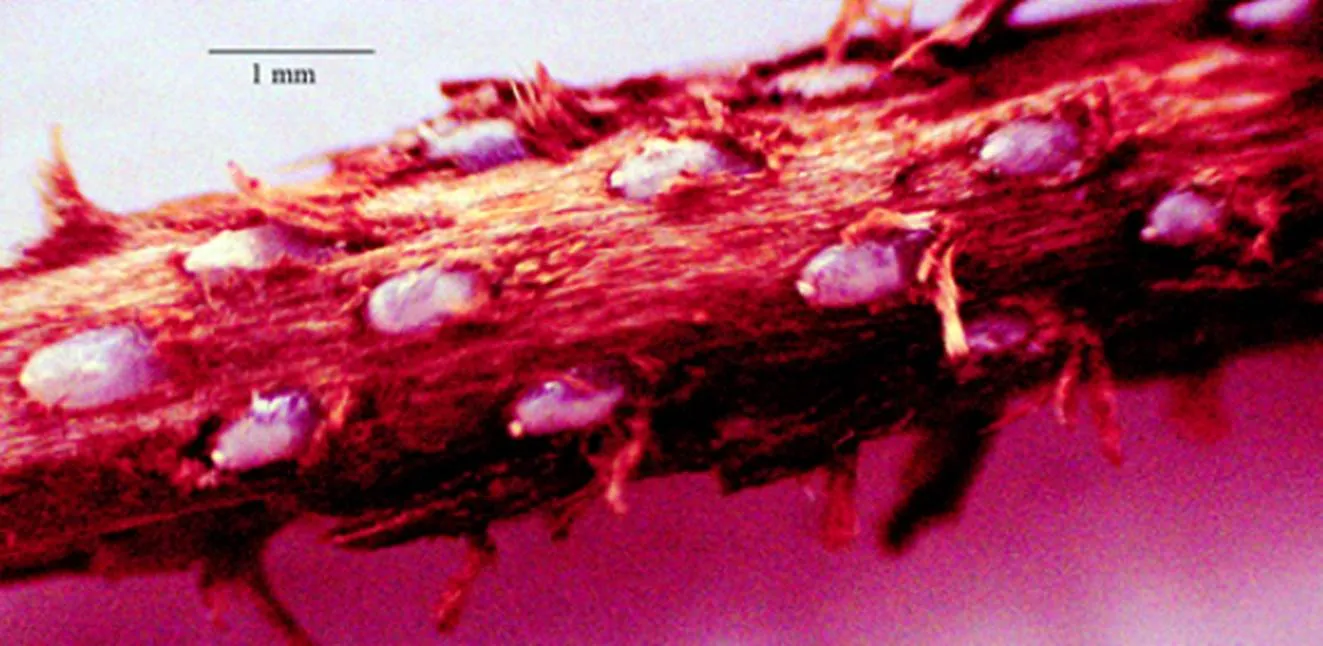
图5 圆纹广翅蜡蝉越冬卵在茶枝内的排列方式及其刻痕

注:图上标不同小写字母,表示在0.05水平上差异显著
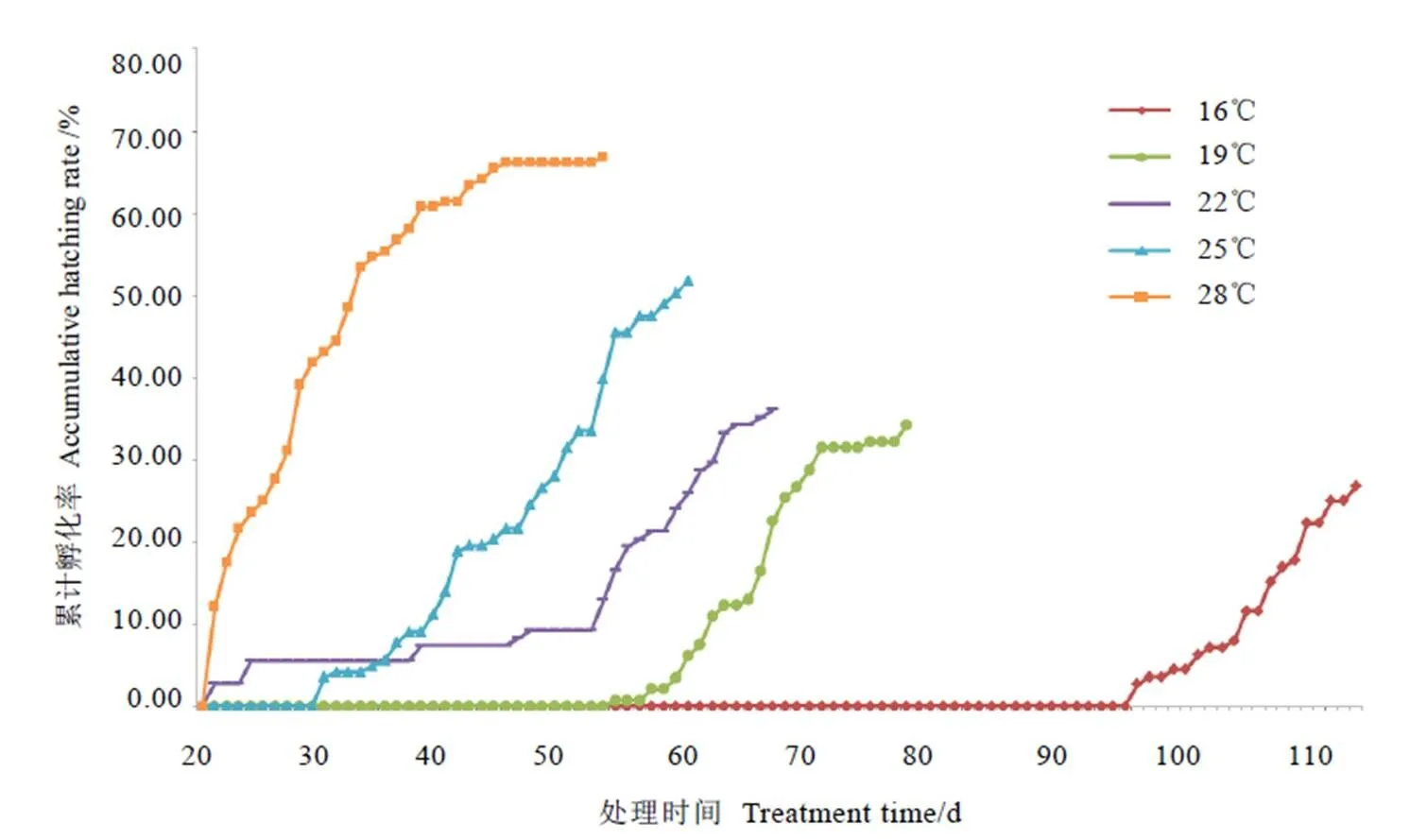
图7 圆纹广翅蜡蝉越冬卵在不同温度条件的累计孵化率

表3 圆纹广翅蜡蝉越冬卵在不同温度条件的孵化率及其差异显著性
注:同列具有相同小写字母者表示在0.05水平上差异不显著
Note: Values followed by the same lowercase letters are not significantly different at the probability of 0.05

图8 温度(x)与圆纹广翅蜡蝉越冬卵发育速率(y)的关系
3 讨论
本研究通过不同类型茶园调查,了解了信阳茶区圆纹广翅蜡蝉成虫产卵对茶枝的为害率,发现马鞍山茶叶试验场阶梯茶园的茶枝受害率最高(13.35%),其次是董家河陈湾村平地茶园(2.19%),两者之间差异显著。究其原因,这可能与马鞍山茶叶试验场阶梯茶园管理较为粗放(茶园修剪不整齐)并种植有樟树等圆纹广翅蜡蝉嗜好的寄主林木有关[9-12]。有学者调查发现,圆纹广翅蜡蝉嗜食樟树,该虫对樟树的为害率为90.00%,雌成虫偏向产卵于一年生枝梢组织中,造成大量的干枯枝,尤其是抽梢不整齐和抽梢时间长的植株受害重,而且,园内嗜好寄主林木越多,栽种密度越大,越容易受到广翅蜡蝉科害虫的为害[9-10]。马鞍山茶叶试验场茶树修剪不整齐,导致抽梢不整齐,且环境隐蔽,湿度较大,有利于圆纹广翅蜡蝉的发生为害[9]。调查中还发现,距离樟树较近的茶树受害较重,圆纹广翅蜡蝉可能在这两种嗜好寄主上转移为害。
本研究发现,圆纹广翅蜡蝉成虫喜欢在直径为(0.23±0.04)cm的茶树枝条上产卵越冬,在茶枝顶端的产卵刻痕到芽梢的距离为(3.99±2.28)cm,产卵刻痕在枝条上的为害长度为(1.36±0.59)cm,单个地环刻在茶树枝条皮层组织内,越冬卵单个镶嵌排列在刻槽内,外露木质丝,外面覆盖一层白色棉絮状蜡丝。这与前人报道的圆纹广翅蜡蝉成虫在其他寄主林木上的产卵为害特征基本一致,而且,倾向于选择在当年生茶树幼嫩枝条皮层组织中产卵越冬[6-10]。由此可看出,圆纹广翅蜡蝉与茶园优势枝梢害虫柿广翅蜡蝉(Jacobi)在茶树上的产卵刻痕排列方式不同,柿广翅蜡蝉越冬卵在茶树枝条组织内呈双行互生倾斜排列,且两行越冬卵之间隔着一层薄薄的木质层[22]。另据研究报道,圆纹广翅蜡蝉雌成虫还产卵于园林植物叶脉组织中[9-10],而目前,并未在茶树叶脉组织中发现越冬卵,需要进一步跟踪调查。
昆虫属于变温动物,其体温敏感地随着外界环境温度的变化而变化,且温度是影响昆虫发育、存活和繁殖力的重要环境因子[23-24]。前人研究发现,不同的昆虫其发育的最适温度条件不同[16-19]。所以,研究温度变化对昆虫发育历期和发育速率的影响具有重要的理论和实践意义。本文通过不同恒温处理研究,发现温度对圆纹广翅蜡蝉越冬卵的孵化率、发育历期和发育速率具有重要的影响。越冬卵在28℃条件下的若虫初孵时间较早(处理后的第21 d),孵化率较高(66.67%),与其他温度处理差异显著,而越冬卵在16℃处理下的若虫初孵时间较迟(处理后的第98 d),孵化率较低(27.19%),但孵化率与19℃和22℃差异不显著,这说明了28℃处理较适合于圆纹广翅蜡蝉越冬卵的发育。圆纹广翅蜡蝉越冬卵的发育历期随着温度的升高而缩短,发育速率随着温度的升高呈线性增加,满足方程=0.001 9-0.022。一般来说,在适温区,昆虫发育速率随着温度的增加呈现线性的增加,可以使用直线(=a+b)拟合来描述温度对昆虫发育速率的影响,便于得出昆虫的发育起点温度和有效积温[18-20]。所以,本文根据有效积温=1/a,发育起点温度=–b/a,得出圆纹广翅蜡蝉越冬卵的发育起点温度为11.58℃,有效积温为526.32 d·℃。
本文在阐明圆纹广翅蜡蝉越冬种群对茶树为害规律的基础上,揭示了不同温度对越冬卵发育的影响,得出了越冬卵的发育起点温度和有效积温,不仅对信阳茶区圆纹广翅蜡蝉越冬种群发生期的预测预报具有非常重要的意义,还有助于信阳茶区圆纹广翅蜡蝉绿色防控策略的制定。总体来说,对该虫的防治,必须要从保护茶园的生态系统出发,结合圆纹广翅蜡蝉发生与为害规律,建议采用较为安全有效的综合防控措施:(1)主要是以农业防治为基础,注重茶园建设,强化茶园管理,增强茶树冠层通风透光,统一进行修剪,及时处理茶园周围圆纹广翅蜡蝉嗜好取食和产卵的寄主林木,及时剪除并集中销毁受害的茶树枝梢,以减少越冬产卵场所和越冬虫源;(2)结合圆纹广翅蜡蝉越冬卵发育起点温度和有效积温,预测若虫在翌年春季的发生期,做好监测工作,因地制宜地运用各种防控措施,控制其大暴发。另外,为了更加准确地预测圆纹广翅蜡蝉若虫的发生期,还需要进一步研究监测自然变温对越冬卵孵化的影响[25],从而确定该虫的防治适期。
[1] 周尧, 路进生. 中国的广翅蜡蝉科附八新种[J]. 昆虫学报, 1977, 20(3): 314-322. Zhou Y. Lu J S. On the Chinese Ricaniidae with descriptions of eight new species [J]. Acta Entomologica Sinica, 1977, 20(3): 314-322.
[2] 张汉鹄. 我国茶树蜡蝉区系及其主要种类[J]. 茶叶科学, 2004, 24(4): 240-242. Zhang H G. Fauna of tea fulgora and the major species in China [J]. Journal of Tea Science, 2004, 24(4): 240-242.
[3] 张玉波. 西南地区广翅蜡蝉科昆虫分类研究(半翅目: 蜡蝉总科)[D]. 贵阳: 贵州大学, 2011. Zhang Y B. Taxnomic studys on Ricaniidae from Southwest China (Hemiptera: Fulgoroidea) [D]. Guiyang: Guizhou University, 2011.
[4] 刘沅. 中国广翅蜡蝉科分子系统学研究[D]. 贵阳: 贵州大学, 2016. Liu Y. Molecular systematics studies on Ricaniidae (Hemiptera: Fulgoromorpha) from China [D]. Guiyang: Guizhou University, 2016.
[5] 刘沅, 张玉波, 杨琳, 等. 基于16S rDNA与Cytb序列的疏广翅蜡蝉属三个近似种的分子鉴定[J]. 环境昆虫学报, 2016, 38(3): 557-564. Liu Y, Zhang Y B, Yang L, et al. Molecular identification of three similar species of genusbased on 16S rDNA and Cytb gene sequences (Hemiptera: Fulgoroidea: Ricaniidae) [J]. Journal of Environmental Entomology, 2016, 38(3): 557-564.
[6] 芦夕芹, 张云贵. 圆纹宽广蜡蝉的识别及防治[J]. 农技服务, 2007, 24(12): 44, 105. Lu X Q, Zhang Y G. Identification and prevention of[J]. Agricultural Technology Service, 2007, 24(12): 44, 105
[7] 谭济才. 湖南省茶园蜡蝉种类调查研究初报[J]. 茶叶科学, 1995, 15(1): 33-37. Tan J C. Investigation on Fulgorid planthoppers (Fulgoridae) in tea gardens of Hunan Province [J]. Journal of Tea Science, 1995, 15(1): 33-37.
[8] 徐冠军, 钟仕田. 宜昌地区桔园蜡蝉种类调查初报[J]. 华中农业大学学报, 1988, 7(2): 196-198. Xu G J, Zhong S T. A preliminary report on the kinds of lanternflies in citrus orchard in Yichang Hubei [J]. Journal of Huazhong Agricultural University, 1988, 7(2): 196-198.
[9] 顾昌华, 龙正权. 广翅蜡蝉优势种对园林植物的为害调查与防控策略探讨[J]. 中国植保导刊, 2007(12): 23-25. Gu C H, Long Z Q. Investigation and prevention and control strategies of the dominant species of Ricaniidae on garden plants [J]. China Plant Protection, 2007(12): 23-25.
[10] 顾昌华. 铜仁地区广翅蜡蝉种类及主要种生物学、生态学和防治研究[D]. 贵阳: 贵州大学, 2011. Gu C H. Research of species in Ricaniidae and their biology, ecology and control of main species in Tongren [D]. Guiyang: Guizhou University, 2011.
[11] 谢广林, 邹海伦, 王文凯. 湖北省广翅蜡蝉科害虫种类调查初报[J]. 湖北农业科学, 2015, 54(10): 2394-2396. Xie G L, Zou H L, Wang W K. A preliminary report on pest speices of Ricaniidae in Hubei Province [J]. Hubei Agricultural Sciences, 2015, 54(10): 2394-2396.
[12] 段文心, 陈祥盛. 中国5种常见宽广蜡蝉形态比较研究[J]. 四川动物, 2020, 39(2): 204-212. Duan W X, Chen X S. Comparative morphology of five commonspecies from China [J]. Sichuan Journal of Zoology, 2020, 39(2): 204-212.
[13] 程娴, 张书平, 余燕, 等. 天敌与广翅蜡蝉空间关系的研究[J]. 应用昆虫学报, 2019, 56(1): 51-61. Cheng X, Zhang S P, Yu Y, et al. Important natural enemies ofand the spatial relationship between these predators and this pest [J]. Chinese Journal of Applied Entomology, 2019, 56(1): 51-61.
[14] 徐德良. 茶树蜡蝉种群生态及控制技术研究[D]. 苏州: 苏州大学, 2009. Xu D L. The study of tea fulgora popution ecology and control technique [D]. Suzhou: Suzhou University, 2009.
[15] 林江. 柿广翅蜡蝉生物学及其与相近种类的形态学比较研究[D]. 杨凌: 西北农林科技大学, 2011. Lin J. Studies on biology and comparative morphology with other related species of(Hemiptera: Fulgoroidea) [D]. Yangling: Northwest Agriculture and Forestry University, 2011.
[16] 高靖淳. 温度对沙葱萤叶甲越冬卵存活和发育的影响[D]. 呼和浩特: 内蒙古农业大学, 2016. Gao J C. Effects of temperature on the survivorship and development of overwintering eggs of(Coleopter: Chrysomelidae) [D]. Hohhot: Inner Mongolia Agricultural University, 2016.
[17] 何莉梅, 葛世帅, 陈玉超, 等. 草地贪夜蛾的发育起点温度、有效积温和发育历期预测模型[J]. 植物保护, 2019, 45(5): 18-26. He L M, Ge S S, Chen Y C, et al. The developmental threshold temperature, effective accumulated temperature and prediction model of developmental duration of fall armyworm,[J]. Plant Protection, 2019, 45(5): 18-26.
[18] 时培建, 池本孝哉, 戈峰. 温度与昆虫生长发育关系模型的发展与应用[J]. 应用昆虫学报, 2011, 48(5): 1149-1160. Shi P J, Ikemoto T, Ge F. Development and application of models for describing the effects of temperature on insects’ growth and development [J]. Chinese Journal of Applied Entomology, 2011, 48(5): 1149-1160.
[19] 石保坤, 胡朝兴, 黄建利, 等. 温度对褐飞虱发育、存活和产卵影响的关系模型[J]. 生态学报, 2014, 34(20): 5868-5874. Shi B K, Hu C X, Huang J L, et al. Modeling the influence of temperature on development, survival and oviposition of the brown planthopper,[J]. Acta Ecologica Sinica, 2014, 34(20): 5868-5874.
[20] Davidson J. On the relationship between temperature and rate of development of insects at constant temperatures [J]. Cambridge University Press, 1944, 13(1): 26-38.
[21] Hoff P D. A first course in Bayesian statistical methods [M]. New York: Springer, 2009: 1-223.
[22] 金银利,马全朝, 张方梅, 等. 信阳茶区柿广翅蜡蝉越冬种群的发生与为害规律[J]. 茶叶科学, 2019, 39(5): 595-601.Jin Y L, Ma Q C, Zhang F M, et al. Occurrence and damage of overwintering populations of Ricania sublimbata Jacobi in Xinyang Tea-producing Area [J]. Journal of Tea Science, 2019, 39(5): 595-601.
[23] Wang R S, Lan Z X, Ding Y Q. Studies on mathematical models of the relationship between insect development and temperature [J]. Acta Entomologica Sinica, 1982, 2(1): 47-57.
[24] Chen J Y, Niu L M, Li L, et al. Influence of constant temperature on development and reproduction ofViggiani (Hymenoptera: Aphelinidae), a parasitoid of the spiraling whiteflyRussell (Hemiptera: Aleyrodidae) [J]. Neotropical Entomology, 2015, 44(2): 160-165.
[25] Wu T H, Shiao S F, Okuyama T. Development of insects under fluctuating temperature: a review and case study [J]. Journal of Applied Entomology, 2015, 139: 592-599.
Oviposition Behavior and Effect of Temperature on the Overwintering Egg Development of
JIN Yinli1,2, MA Quanchao1, ZHANG Shaojie1, YIN Jian1, SHI Hongzhong1, GENG Shubao1*
1. Xinyang Agriculture and Forestry University, Xinyang 464000, China; 2. Henan Key Laboratory of Tea Plant Comprehensive Utilization in South Henan, Xinyang 464000, China
In order to clarifyoviposition behavior and effect of temperature on the overwintering egg development ofWalker, its occurrence and damage were systematically investigated by five-point sampling method, and the effect of temperature (16℃, 19℃, 22℃, 25℃ and 28℃) on the development of its overwintering egg was also studied. The results show thatpreferred to lay overwintering eggs in tea branches with diameter of (0.23±0.04)cm. The mean length of oviposition scar damaged on tea branch was (1.36±0.59)cm, and the damage rates (13.35%) of tea branch in Maanshan tea garden (managed extensively and more host plants around) was significantly higher than others. The results also indicate that the developmental duration of overwintering eggs decreased with the increase of temperature. The developmental duration of the overwintering eggs was (28.75±7.03)d at 28℃. The initial hatching time of nymph was earlier and the hatching rate was higher at 28℃. The linear relationship between the temperature and the developmental rate ofoverwintering eggs was=0.001 9-0.022, and the developmental threshold temperature was 11.58℃. The effective accumulated temperature was 526.32 d·℃. This study provided a theoretical basis for prediction and control strategies of the overwintering population of this pest.
, oviposition behavior, tea plant, developmental duration, developmental threshold temperature, effective accumulated temperature
S571.1;S435.711
A
1000-369X(2020)06-807-10
2020-06-24
2020-07-13
茶园化肥农药减施增效技术集成研究与示范项目(2016YFD0200900)、河南省科技攻关项目(172102110235)、信阳农林学院科技创新团队项目(KJCXTD-201903)、信阳农林学院青年教师科研基金资助项目(201701007、2018LG002)
金银利,女,博士,讲师,主要从事昆虫生态学研究,jinyinli02@163.com。*通信作者:shubaogeng@163.com
