南亚热带混交人工林树种丰富度与土壤微生物多样性和群落组成的关系
2020-12-14宋战超刘世荣胡佳佳明安刚李朝英伍俊廷史作民卢立华蔡道雄
宋战超,王 晖,*,刘世荣,胡佳佳,明安刚,陈 海,李朝英,伍俊廷,史作民,卢立华,蔡道雄
1 中国林业科学研究院森林生态环境与保护研究所,国家林业和草原局森林生态环境重点实验室, 北京 1000912 中国林业科学研究院热带林业实验中心,广西友谊关森林生态系统国家定位观测研究站, 凭祥 532600
土壤微生物在森林生态系统中发挥着不可替代的重要作用,积极参与到森林生态系统养分转化和循环[1],对维持森林生态系统稳定和功能以及植物多样性至关重要[2]。森林土壤微生物群落结构和多样性受林下凋落物性质和土壤根系分泌物等生物因子及土壤环境等非生物因子的共同影响[3- 5]。同时,土壤微生物通过对土壤理化性质和土壤肥力作用,影响森林植被生物量和生产力[6]。
近年来,研究生态系统地上和地下生物多样性之间的联系一直是生态学研究的热点。针对土壤微生物群落与植物群落关系的研究越来越多,但多以草地生态系统或天然林生态系统为主。Zak等[7]研究了植物多样性对土壤微生物的影响,发现随着植物多样性的增加,土壤真菌生物量明显增加。青藏高原天然草原的研究发现,随着植物多样性的增加,真菌α和β多样性分别相应增加[8]。Chen等[9]在我国亚热带森林的不同树种多样性梯度实验研究显示,树种特性是主导土壤细菌和真菌群落多样性和结构变化主要因素,且对土壤真菌的影响比土壤细菌效果更强,而树种多样性并非是主要影响因子。基于我国亚热带森林生物多样性与生态系统功能实验,发现了不同功能型土壤真菌驱动亚热带森林群落多样性的作用模式[10]。有关生物多样性与生态系统功能关系的研究表明,生物多样性是生态系统功能的主要驱动力[11]。不同树种在森林群落中占据不同的高度和空间从而创造了更多的生态位,促进了群落利用阳光等环境资源的能力,提高了森林生态系统生物量和生产力。而我国人工林以单一树种种植为主,由于生物多样性下降,森林各种功能与生产力得不到充分发挥,降低了森林适应能力和稳定性,还会造成地力衰退、森林系统功能退化等问题[12]。如在我国南方地区大面积分布的桉树人工纯林,丛枝菌根真菌的种丰度随种植年份的增加逐渐下降,优势种Funneliformsgeosporus的丰度在2年生和10年生林地中显著降低[13]。但是,树种多样性增加使森林植被组成和生态系统结构、功能趋于复杂,更多树种混交后树种多样性会随之增加以及树种功能特性发生重叠,不同气候区、不同类型森林生态系统植物种类和环境等条件差异也比较大,因此尚需通过研究进一步增强对森林植物多样性与土壤微生物多样性关系以及潜在机制的认知,进而提高有关土壤微生物对“树种多样性与土壤理化性质和肥力”关系影响机制的理解。
亚热带森林生态系统是我国陆地碳汇潜力的主要来源[14]。我国南方亚热带地区树种多样性丰富,人工林的面积和蓄积均居全国之首。Huang等[15]通过野外多树种新造林实验研究发现随树种多样性增加林分生产力显著增加。本研究面向人工林生态系统固碳增汇功能提升的国家需求,针对土壤微生物群落组成与树种多样性关联机制这一关键科学问题,选取我国南亚热带经近40年近自然管理形成的多树种、多层次、天然更新良好的异龄混交人工林生态系统为对象,采用高通量测序技术研究随树种丰富度变化土壤微生物α多样性和群落组成的变化,以及主要的生物和环境驱动因子,旨在探讨多树种混交如何对南亚热带人工林土壤微生物生物量和群落结构组成产生影响,为进一步揭示树种多样性与地下土壤微生物多样性关系提供科学依据,并为制定该区域人工林通过树种丰富度合理组配调控提升生态系统功能的经营策略提供科技支撑。
1 材料和方法
1.1 研究区概况与实验设计
研究区位于广西壮族自治区凭祥市中国林业科学研究院热带林业实验中心国家林业和草原局广西友谊关森林生态系统定位研究站(22°06′50″—22°07′27″N,106°46′48″—106°47′22″E)。该地区属典型南亚热带季风气候区,海拔约300 m,年平均气温20.5—21.7℃,年平均降雨量1200—1500 mm,多集中在4—9月份。主要地貌类型以低山丘陵为主,土壤以砖红壤和红壤为主,成土母岩主要有泥岩夹砂岩、砾状灰岩和石灰岩等。本研究的试验区在杉木林采伐迹地,自1979年选取21个乡土树种以块状混交布局建立的试验林,经过近40年的人工经营和天然更新,试验区已形成多树种、复层结构的混交人工林,试验区面积100 hm2。
在试验区内随机设置54个20 m × 20 m样地,并对每个样地内胸径(Diameter at breast height,DBH)≥1 cm林木的胸径、树高和坐标进行每木调查。依据每个样地内的树种丰富度将54个样地划分成10组,形成由1—10个树种的丰富度梯度。其中第一组(1个树种)有7个样地,第二组(2个树种)有7个样地,第三组(3个树种)有3个样地,第四组(4个树种)有4个样地,第五组(5个树种)有4个样地,第六组(6个树种)有7个样地,第七组(7个树种)有5个样地,第八组(8个树种)有10个样地,第九组(9个树种)有3个样地,第十组(10个树种)有4个样地,各样地的树种组成详见附件7。
1.2 样品采集与处理

1.3 土壤理化性质、细根生物量和碳化学组分的测定
土壤pH值采用FE20 pH计测定,土水比为1∶2.5。采用重铬酸钾外加热法和凯氏定氮法分别测定土壤有机碳和全氮含量[16]。采用盐酸和硫酸萃取后分光度法测定土壤有效磷含量[17]。采用2 mol/L KCl浸提,靛酚蓝比色法测定铵态氮,紫外分光光度法测定硝态氮[18]。
土样过2 mm筛时,挑选出直径 ≤ 2 mm的细根活体,用水反复冲洗去除附着在根系表面的泥土后,放置烘箱用65℃烘干至恒重,用电子天平称重(精确到0.001 g),依据如下公式计算各样地的细根生物量:
细根生物量(g/m2)=平均每个土芯中细根干重(g)/[π(Φ/2)2× (m2/104cm2)](Φ=5.0 cm)
采用13C固体核磁共振波谱(13C CPMAS NMR)分析土壤有机碳不同化学组分比例。13C固体核磁共振波谱被分为四组化学位移区域,以此区分土壤有机碳的不同组分:烷基碳(δ=0—45,alkyl C),氧烷基碳(δ=45—110,O-alkyl C),芳香碳(δ=110—160,aromatic C),羰基碳(δ=160—210,carbonyl C)。
1.4 土壤微生物总DNA提取和高通量测定
称取约0.25 g经冷冻干燥处理的土壤样品,使用MO BIO PowerSoil(MO BIO Laboratories,Carlsbad,USA)试剂盒提取土壤微生物总DNA,采用NanoDrop分光光度计进行定量提取DNA的质量和浓度,并用1%琼脂糖凝胶电泳对提取的DNA质量进行检测,合格后用于构建文库;以各土壤样品微生物总DNA为模板,以细菌V4和V5区特异性引物(15F 5′-GTGCCAGCMGCCGCGGTAA- 3′;926R 5′-CCGTCAATTCMTTTGAGTTT- 3′)[19]和真菌ITS1区特异性引物(ITS1F 5′-CTTGGTCATTTAGAGGAAGTAA- 3′;2043R 5′-GCTGCGTTCTTCATC GATGC- 3′)[20]进行PCR(polymerase chain reaction)扩增;采用Illumina MiSeq测序平台对PCR扩增产物进行双端测序分析,微基生物科技(上海)有限公司完成测序。
土壤微生物目标片段的PCR产物测序完成后,对有效序列进行质控、拼接和精确去杂等处理,得到优化序列。优质序列利用Usearch软件基于97%的相似水平进行分类操作单元(operational taxonomic units,简称OTUs)。根据Silva细菌数据库[21]和Unite[22]真菌数据库利用 RDP Classifier进行物种注释和分类。利用QIIME(Version l.7.0)软件依据样本文库的OTUs丰度信息计算土壤样品的细菌和真菌多样性(Faith′s Phylogenetic Diversity,Faith′s PD指数)。利用R语言软件VEGAN包对土壤细菌和真菌β多样性进行了距离冗余分析(Distance-based redundancy analyses,db-RDA),并筛选主要解释变量。
1.5 统计分析
2 结果与分析
2.1 树种丰富度与土壤细菌和真菌α多样性的关系
通过对本研究中的人工林树种丰富度梯度序列样地建立不同树种丰富度梯度(1—10个树种)与土壤细菌α多样性之间的相关关系,树种丰富度与土壤细菌α多样性的关系如图1所示。Pearson相关分析的结果表明,土壤细菌α多样性随树种丰富度增加整体间存在增加的趋势,但影响不显著(P>0.05)。通过对土壤细菌α多样性和土壤理化性质关系的分析,发现土壤细菌α多样性与土壤pH和芳香碳组分呈显著正相关(P< 0.05),而与有机碳、铵态氮、碳氮比、烷基碳组分及细根生物量呈显著负相关(P<0.05)。采用逐步回归分析揭示土壤理化性质与土壤细菌α多样性之间的关系,结果表明土壤pH和铵态氮是土壤细菌α多样性变化的主要影响因素(表1)。
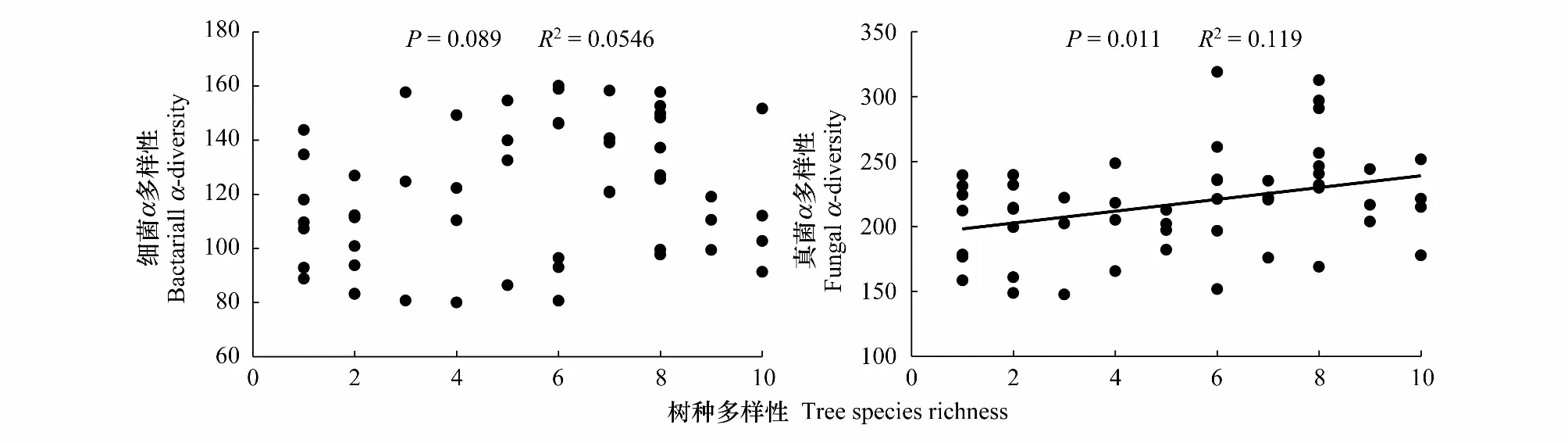
图1 树种丰富度与土壤细菌α多样性和真菌α多样性的关系Fig.1 Relationship of tree species diversity on soil bacterial α-diversity and fungal α-diversity
树种丰富度与土壤真菌α多样性的关系如图1所示。Pearson相关分析的结果表明,土壤真菌α多样性与树种丰富度之间存在显著正相关关系(P<0.05),随树种丰富度增加,土壤真菌α多样性显著提高。通过对土壤细菌α多样性和土壤理化性质关系的分析,发现土壤真菌α多样性与土壤pH和碳氮比呈显著正相关(P<0.05),而与细根生物量呈显著负相关(P< 0.05)。逐步回归分析表明土壤pH和有效磷是土壤真菌α多样性变化的主要影响因素(表1)。

表1 土壤细菌和真菌α多样性与环境因子的多元统计结果
2.2 不同树种丰富度土壤细菌和真菌群落组成变化
对不同树种丰富度土壤微生物群落组成进行PerMANOVA分析,结果发现土壤细菌β多样性在不同树种丰富度之间存在显著差异(表2)。对54个土壤样品进行Illumina MiSeq高通量测序,数据经过滤优化,共得到1964785个细菌优化序列,平均每个样品36385个序列,共得到4769个OTUs。在纲水平上土壤细菌群落共鉴定48纲,相对丰度大于1%的纲共15个。样品中土壤细菌群落主要由酸杆菌纲(13.19%)、α变形菌纲(13.06%)、γ变形菌纲(9.85%)、β变形菌纲(6.00%)和δ变形菌纲(5.61%)组成,其他微生物种类以低丰度(小于5.00%)存在,如:纤线杆菌纲(3.45%)、浮霉菌纲(0.80%)。树种丰富度增加变化改变了土壤细菌β多样性,在不同树种丰富度梯度下α变形菌纲、迷踪菌纲、γ变形菌纲、浮霉菌纲、Ignavibacteria(未命名)和螺旋体纲差异显著(P<0.05)(图2)。
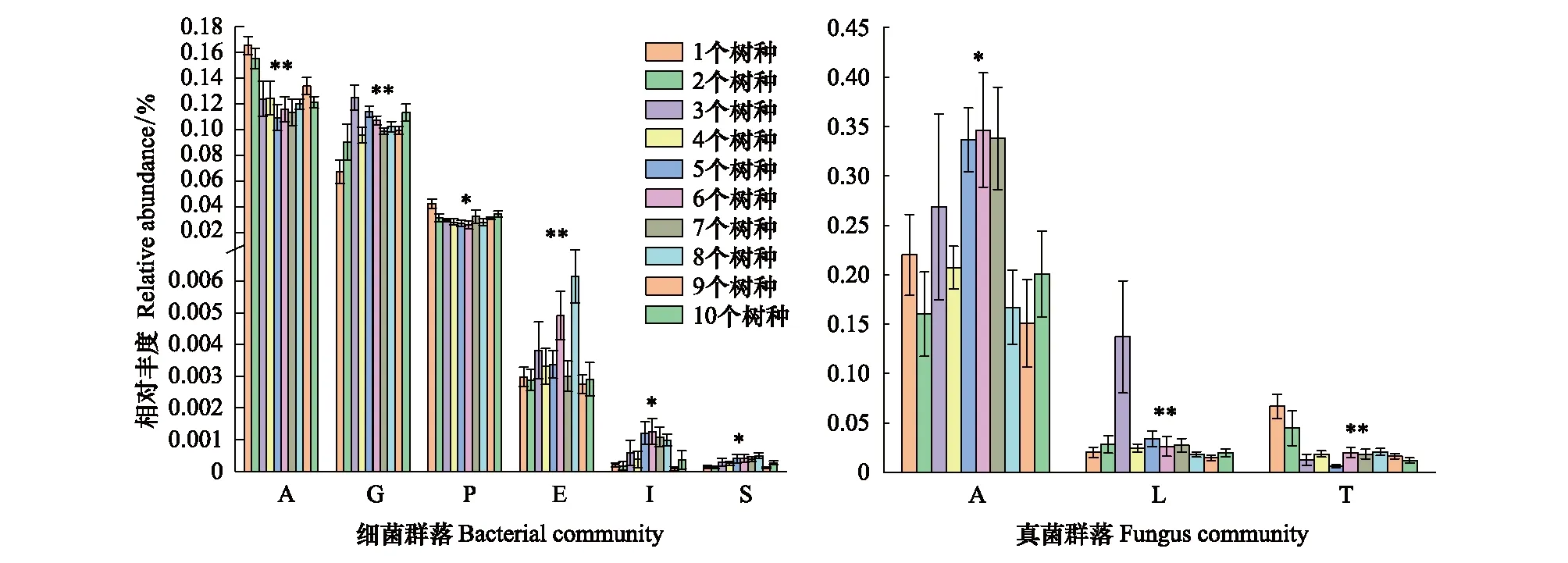
图2 不同树种丰富度与土壤细菌群落和真菌群落组成变化Fig.2 Diversity of different tree species and composition of soil bacterial community and fungal community 细菌群落中Aα变形菌纲;G:γ变形菌纲;P:浮霉菌纲;E:迷踪菌纲;I:Ignavibacteria,未命名;S:螺旋体纲;真菌群落中A:伞菌纲;L:茶渍纲;T:银耳纲;**表示显著水平P< 0.01,*表示显著水平P< 0.05

表2 不同树种丰富度梯度下土壤细菌和真菌群落PerMANOVA分析
在不同树种丰富度梯度下土壤真菌β多样性存在显著差异(表2)。真菌测序共得到2068537个优化序列,聚集在8124个OTUs中,平均每个样品38306个序列。在纲水平上土壤真菌群落共鉴定34纲,相对丰度大于1%的纲共8个。样品中土壤真菌群落主要由伞菌纲(23.38%)组成,其他种群丰度相对较低,如茶渍纲(3.07%)、银耳纲(2.84%)。土壤真菌群落中的伞菌纲、茶渍纲和银耳纲在不同树种丰富度梯度下差异显著(P<0.05)(图2)。
2.3 环境变量对土壤细菌和真菌群落组成的影响
采用db-RDA分析来评估环境变量对土壤细菌群落组成的影响。分析结果显示,土壤细菌群落在第一主轴(横轴)主要由土壤pH和芳香碳组分贡献,累计解释变量达到92.86%。第二主轴(纵轴)累计解释变量为5.81%,主要由土壤硝态氮贡献。土壤细菌群落组成与土壤环境因子密切相关,土壤pH、硝态氮和芳香碳组分是树种丰富度梯度间土壤细菌群落组成差异的主要影响因素(图3)。
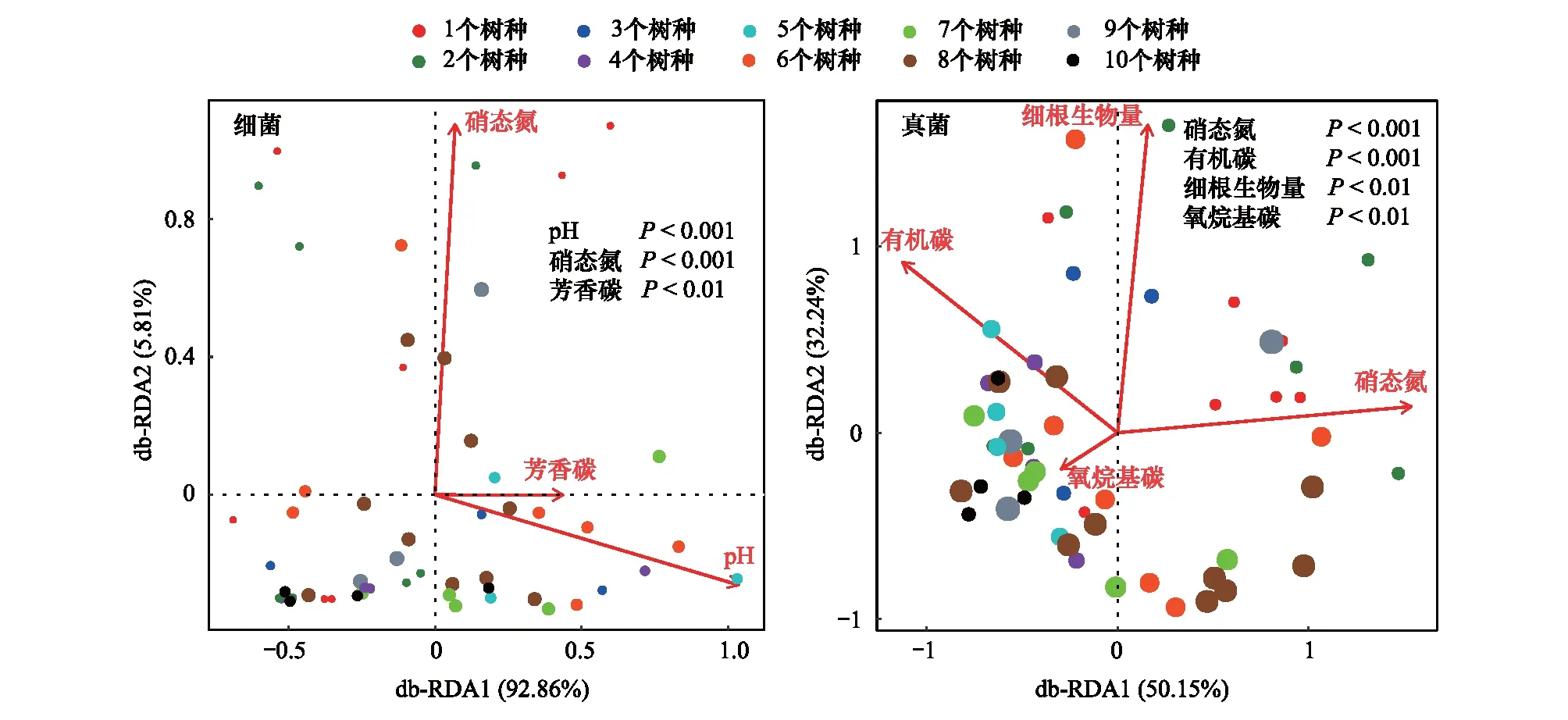
图3 土壤细菌和真菌群落组成与环境变量的db-RDA分析Fig.3 Distance-based redundancy analyses (db-RDA) plot showing the relationship of abiotic factors to community composition of bacteria and fungi
不同树种丰富度梯度下环境变量与土壤真菌群落组成的db-RDA分析如图3所示。两轴共解释土壤真菌群落结构82.39%的变异,其中第一主轴(横轴)累计解释变量为50.15%,主要由土壤硝态氮贡献;第二主轴(纵轴)累计解释变量为32.24%,主要由细根生物量贡献。结果表明土壤硝态氮、有机碳、细根生物量和氧烷基碳组分是影响土壤真菌群落组成在不同树种丰富度梯度间差异的主要因素。
3 讨论
3.1 南亚热带混交人工林树种丰富度与土壤微生物α多样性的关系
前人有关植物多样性与土壤微生物群落组成的研究主要在草原[25]和天然林生态系统[26]开展,并发现植物多样性增加会提高土壤真菌α多样性。本研究中也得到了相似的结果,在南亚热带经近40年近自然管理,形成的多树种、多层次、天然更新良好的异龄混交人工林生态系统中,土壤真菌α多样性随着树种丰富度增加而提高(图1)。通过人工混交不同乡土树种,进而构建成自然恢复并天然更新的混交人工林生态系统中树种丰富度与土壤微生物α多样性的关系趋近于天然草地和森林所呈现的规律的现象一般性的解释是有更高物种丰富度的生态系统能更有效地利用各种资源,有更高的生产力并创造了更多的生态位从而容纳了更多的土壤微生物(互补假说)[5,27]。但在本研究中还发现不同树种丰富度下土壤细菌α多样性差异不显著,说明了树种丰富度与土壤真菌α多样性的相关性较之与土壤细菌α多样性的相关性更明显。Johnson[28]等研究认为细菌多样性和植物多样性不相关,这与本研究结果一致。有研究发现,土壤微生物更依赖于某种特定植物的存在(单一假说),而不是植物多样性本身[29- 30]。因此,在下一步的研究中,将更深入的开展树种功能特性对土壤微生物功能类群影响机制的研究,并进一步辨析地下微生物过程的变化是由树种丰富度还是树种特性主导。
本研究中相关分析结果表明,土壤细菌和真菌α多样性与土壤理化因子呈现不同程度的相关性,说明该区域土壤理化因子是影响微生物数量的关键因素。Shen等[31]在长白山研究了6种典型植被类型,分析了随海拔梯度土壤细菌群落组成和多样性的变化趋势,发现土壤细菌分布主要受土壤pH驱动。朱平等[32]通过研究祁连山4种不同植被类型对土壤微生物的影响也得出了相似的结论。在本研究中,试验区内土壤细菌α多样性与土壤pH和铵态氮显著相关(P<0.05),印证了土壤pH同样是影响本研究中南亚热带多树种人工林生态系统树种丰富度之间土壤细菌群落分布主要因子。本研究还发现树种丰富度增加显著提高了土壤pH,从而改善了研究区内土壤微生物生存环境,可能是土壤微生物多样性增加的原因。庄林杰等[33]研究了从湖泊到岸边深层土壤中土壤微生物变化情况,发现细菌多样性与土壤铵态氮呈负相关,与本研究结果一致。其原因可能是土壤中铵态氮增加导致土壤酸化[34],而通常认为土壤细菌群落在中性pH条件下丰富度和多样性较高[35],本研究中pH与土壤细菌多样性的正相关关系也为此提供了依据。在森林生态系统中树种丰富度增加会改变森林凋落物输入量和凋落物分解过程,进而影响土壤中养分含量。有研究表明,真菌群落相较于细菌群落更直接的依赖于树木凋落物,且许多真菌与树种根系共生形成菌根,使得真菌类群与树木的生物营养相互作用更强[36],这可能是本研究中真菌α多样性受土壤有效磷影响的原因。
3.2 南亚热带混交人工林树种丰富度变化影响土壤微生物群落组成
本研究的多树种混交人工林生态系统中树种丰富度增加显著改变了土壤微生物群落组成,与以往在我国亚热带天然林开展的研究结果相一致[9]。土壤微生物分布广泛且适应性强,但在特定区域的土壤微生物群落组成受到林下特定环境因素的调控。许多研究表明,在森林生态系统中植物群落多样性对土壤微生物群落结构具有显著影响[28,37]。土壤细菌群落以酸杆菌和α变形菌为主,但就具体细菌菌群而言,随树种丰富度增加菌群并没有表现出明显增加或者减少的趋势。Xia[38]等人对我国典型森林土壤细菌群落的研究发现,土壤细菌丰度排在前五位的菌群分别为放线菌、酸杆菌、α变形菌、疣微菌和浮霉菌。本研究中不同树种丰富度下土壤细菌群落中α变形菌差异性显著。由此可见,树种丰富度增加通过改变相对丰度较高的土壤细菌菌群使土壤细菌群落组成差异性显著(图2)。在本研究中土壤真菌群落组成变化也表现出相似的现象,伞菌纲作为土壤真菌群落中的重要组成菌群,在不同树种丰富度梯度下差异性显著(图2)。
人工林不同树种组成可通过影响森林生态系统中凋落物数量和质量,根系生物量、根系周转和根系分泌物等对土壤理化性质和土壤养分产生不同的影响,进而改变土壤微生物的生物量和群落组成[3-4,36]。土壤pH参与土壤生物地球化学循环过程并对土壤理化性质具有重要的调节作用,是影响细菌群落组成的重要因子。本研究中db-RDA分析表明土壤细菌群落组成差异中的98.67%能够被土壤pH、硝态氮和芳香碳组分三个环境变量所解释,其中土壤pH占主要贡献(图3),结合土壤pH对土壤细菌α多样性的显著影响,说明了土壤pH是影响土壤细菌群落组成和多样性的主要因素。适宜的pH为土壤微生物提供了良好的生存条件,土壤pH值能够通过影响土壤基质的组成、化学性质从而改变土壤微生物多样性和群落组成。本研究中土壤硝态氮、有机碳、细根生物量和氧烷基碳组分是导致土壤真菌群落差异的主要影响因素(图3)。Wang[24]等人研究表明在南亚热带氮肥添加导致土壤中铵态氮和硝态氮含量增加,从而改变土壤pH,这可能是土壤硝态氮影响土壤真菌群落组成的原因。土壤有机质作为土壤理化性质的重要指标之一,是土壤养分循环转化的核心。土壤有机碳含量的变化能够影响土壤肥力和土壤微生物群落的变化[39]。王淼等[40]研究结果也表明土壤理化性质特别是土壤养分状况的差异是土壤微生物群落结构差异的主要影响因素。树种丰富度增加,往往会对根系的分布和生物量产生较大影响[41],通过改变根系分泌物从而导致土壤微生物群落变化。字洪标等[42]通过对青海7种不同林分土壤微生物群落结构的研究表明,细根生物量是导致土壤微生物群落差异的因素之一,与本文研究结果相一致。
4 结论
本研究选取了南亚热带多树种混交人工林为研究对象,研究了人工林树种丰富度对土壤微生物多样性和群落组成的影响。结果表明,树种丰富度增加对土壤微生物多样性和群落结构具有一定的影响,其中土壤真菌表现出比细菌更加敏感的响应。土壤pH是影响土壤细菌多样性和群落组成的主要影响因子,土壤碳氮组分和细根是影响土壤真菌多样性和群落组成关键因素。研究结果说明了南亚热带不同乡土树种块状混交后天然更新形成多树种混交林过程中,树种组成和多样性的变化通过改变土壤理化性状和根系生物量对土壤微生物群落有显著影响。
