基于宏基因组测序的扶桑绵粉蚧内共生菌多样性研究
2020-12-11马福欢蓝翊文何成伟白华菊
钟 勇,马福欢,蓝翊文,何成伟,白华菊
1凭祥海关,广西 凭祥 532600;2友谊关海关,广西 凭祥 532600
胸喙亚目昆虫(包括粉蚧、其他蚧虫、蚜虫、木虱和粉虱)以植物汁液为食,植物汁液营养上极不平衡,虽然富含主要以蔗糖存在的碳水化合物,但脂质和蛋白质含量非常少,因此,它们均需内共生菌提供必需氨基酸和其他营养物质来保证正常的生长发育(Baumannetal.,2000; Moran & Telang,1998;Munsonetal.,1992)。研究发现,粉蚧体内含有初生内共生菌(Fukatsu & Nikoh,2000; Moran & Telang,1998;Munsonetal.,1992)和次生内共生菌(Kanthetietal.,1996;von Dohlenetal.,2001)。在研究昆虫内共生菌的功能及其与宿主间的相互关系时,传统的方法是通过去除内共生菌或者利用营养缺失实验来观察宿主生理生化的变化。然而,内共生菌不能体外培养,这阻碍了进一步深入了解共生菌在宿主体内所起的作用(杨义婷等,2014)。随着PCR技术的发展和测序技术的进步,转录组学、蛋白质组学、基因组学等技术的发展,为昆虫内共生菌研究提供了便利;最新的基因组学工具,特别是新一代高通量测序工具和分析软件,使得内共生菌的大规模宏基因组测序和价格低廉的单个菌基因组测序成为可能,为明确共生菌功能提供了直接证据,同时也促使从种群和群落水平研究细菌、真菌等与昆虫宿主共生关系成为可能,而这些都将为最终发现共生菌与宿主共生关系的潜在机制奠定基础。
扶桑绵粉蚧PhenacoccussolenopsisTinsley隶属半翅目Hemiptera蚧总科Coccoidea粉蚧科Pseudococcidae绵粉蚧亚科Phenacoccinae绵粉蚧属Phenacoccus,是一种重要的入侵害虫,为害棉花GossypiumhirsutumL.和其他重要作物,现已几乎遍布全球(Fuchsetal.,1991;Wang & Yang,2010)。目前,国内外对其研究多集中在生物学特性方面,内共生菌的研究相对较少(黄芳等,2015)。这些报道中,如扶桑绵粉蚧入侵后寄主范围不断扩大(陈哲等,2012)、出现体色变异型种群(朱艺勇等,2011;Zhaoetal.,2014)等可能均有内共生菌参与。同时,不同的研究对扶桑绵粉蚧繁殖方式的报道有较大差异(关鑫等,2011;唐远等,2013; Hodgsonetal.,2008;Huangetal.,2013;Vennilaetal.,2010),其原因可能是不同种群内共生菌感染情况不同,尤其是与生殖有关的内共生菌的感染情况。因此,扶桑绵粉蚧内共生菌多样性的研究,将为共生菌与粉蚧的相互作用研究提供基础,也为研究扶桑绵粉蚧的入侵机制提供新的研究思路。
1 材料与方法
1.1 试虫来源
扶桑绵粉蚧为国家热带和亚热带水果检疫重点实验室(凭祥)使用南瓜Cucurbitamoschata(Duch.ex Lam.)Duch.ex Poiret饲养的室内种群,原寄主为木槿HibiscusrosasinensisL.(广西凭祥)。
1.2 高通量测序
收集试虫,分为2个重复,每重复雌成虫30头;委托深圳华大基因股份有限公司对整虫样本开展微生物宏基因组测序。
1.3 测序数据处理
原始测序数据6.28×109reads。过滤掉低质量序列、接头序列等杂质,从而得到6.12×109的高质量Clean Data。组装过滤后的reads,以得到更长的contig。然后将reads分别比对回每个组装结果以评估组装效果,最后综合考虑N50和比对率情况,选择一个最佳k-mer和其对应的组装结果。最终组装获得1.60×105contigs,组装长度2.56×108bp,最大contig长度89956 bp,最小contig长度300 bp,组装平均长度1600 bp,reads利用率68.32%。
1.4 内共生菌丰度分析
使用MEGAN(version 4.6)对nr比对结果进行处理。MEGAN根据LCA算法对BLAST比对结果根据NCBI物种分类树进行重新梳理,从而得到每条序列的物种注释信息。将同一物种注释的基因丰度累加,得到该物种在样品中的物种丰度。
1.5 内共生菌基因分析
对每个样品使用MetaGeneMark进行基因预测,构建高质量基因集,将reads比对到高质量基因集计算基因丰度。对KEGG(89.1)、COG等数据库的分析,先将基因注释到此类数据库。基于Reads与基因集的比对结果,获取该类功能数据库的定量(丰度)和定性(基因个数)信息。
2 结果与分析
2.1 扶桑绵粉蚧内共生菌种类及其丰度
根据注释结果,样本共鉴定获得29个门,47个纲,105个目,178个科,245个属,299个种。在界分类水平上,扶桑绵粉蚧内共生物测序共注释了古细菌Archaea、细菌Bacteria、真核生物Eukaryota和病毒Viruses;细菌为优势种群,相对丰度达85.16%。在门分类水平上,共注释了子囊菌门Ascomycota、担子菌门Basidiomycota、Candidatus Magasanikbacteria等29个门;其中变形菌门Proteobacteria为优势种群(相对丰度81.36%)。在纲分类水平上,共注释了α-变形菌纲Alphaproteobacteria、β-变形菌纲Betaproteobacteria、衣原体纲Chlamydiia等47个纲;γ-变形菌纲和β-变形菌纲为优势种群,相对丰度远超其他种群(分别为50.04%和27.68%)。在目分类水平上,共注释了交替单孢菌目Alteromonadales、伯克氏菌目Burkholderiales、弯曲菌目Campylobacterales等105个目;肠杆菌目Enterobacterales为优势种群(相对丰度42.65%)。在科分类水平上,共注释了酸杆菌科Acetobacteraceae、芽孢杆菌科Bacillaceae、弯曲菌科Campylobacteraceae、Debaryomycetaceae等178个科;肠杆菌科Enterobacteriaceae为优势种群(相对丰度33.48%)。在属分类水平上,共注释了醋酸杆菌属Acetobacter、芽孢杆菌属Bacillus、Caballeronia等245个属;伯克氏菌属Burkholderia为优势种群(相对丰度13.12%);CandidatusTremblaya相对丰度3.69%排名第二。在种分类水平上,共注释了Acetobactermalorum、脆弱类杆菌属Bacteroidesfragilis、Caballeroniagrimmiae等299个种;相对丰度为2.58%的CandidatusTremblayaphenacola为优势种群(表1)。

表1 扶桑绵粉蚧内共生菌丰度(前10)
2.2 扶桑绵粉蚧内共生菌基因分析
样本所测序列共预测了36505个基因,其中77.27%的基因分布在200~499 nt的范围内(图1)。基因集KEGG注释了776个基因,其中代谢相关基因最多(384个),占49.48%;其次为遗传信息处理基因(207个),占26.68%;最少的为环境信息处理基因(8个),仅占1.03%。代谢相关基因除了综合性代谢全谱基因(157个)外,最多的为氨基酸代谢基因(64个),其次为碳水化合物代谢基因(36个)、辅酶和维生素代谢基因(35个)(图2),分别占28.19%、15.86%和15.42%。基因集COG注释了398个基因,其中翻译、核糖体结构和生物发生的基因最多(110个),占27.64%;其次为氨基酸转运和代谢基因(49个)(图3),占12.31%。

图1 扶桑绵粉蚧内共生菌基因长度分布图
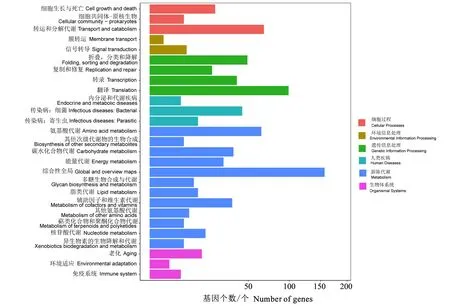
图2 扶桑绵粉蚧内共生菌KEGG通道注释图

图3 扶桑绵粉蚧内共生菌COG通道注释图
3 讨论
本研究采用宏基因组测序技术对扶桑绵粉蚧内共生菌多样性及群落结构组成进行了研究。其中,变形菌门、γ和β变形菌,以及β-变形菌的CandidatusTremblaya是其优势种群,该结果与其他昆虫的报道相似(毛迎新等,2018;Konoetal.,2008;McCutcheon & von Dohlen,2011;Sergioetal.,2015)。
很多变形菌门细菌与营养代谢相关,这些种群丰度占优势的内共生菌很可能在扶桑绵粉蚧的营养代谢中起很大作用。CandidatusTremblaya是一种与绵粉蚧亚科和粉蚧亚科有共生关系的内共生菌,且多数绵粉蚧亚科粉蚧具有独特的内共生菌嵌套体CandidatusTremblayaphenacola(Sergioetal.,2015)。实验结果证明,在扶桑绵粉蚧体内,CandidatusTremblayaphenacola是种的分类水平的优势种群。此外,同属的CandidatusBaumanniacicadellinicola和CandidatusEntotheonellasp.相对丰度排名分别占第6和第10。
研究发现,在基因KEGG注释时,扶桑绵粉蚧基因主要为代谢相关基因和遗传信息处理基因,代谢相关功能基因中,又以氨基酸代谢基因为主,数量远远高于其他代谢基因。在橘臀纹粉蚧Planococcuscitri(Risso)体内,嵌套菌CandidatusMoranellaendobia和CandidatusTremblayaprinceps共同参与了其体内的氨基酸代谢(López-Madrigaletal.,2013;McCutcheon & von Dohlen,2011)。然而,同样的功能在PhenacoccusavenaeBorchsenius体内却由CandidatusTremblayaphenacola一种共生菌完成(López-Madrigaletal.,2014)。为此,可推测扶桑绵粉蚧通过刺吸式口器取食富含糖分却缺乏必需氨基酸的植物韧皮部汁液,内共生菌CandidatusTremblaya为其提供了必需氨基酸等营养物质。同时,研究检测到Drosophilaananassae内共生菌Wolbachia也是扶桑绵粉蚧体内的优势种群之一(相对丰度1.04%)。Wolbachia广泛存在于节肢动物中,对宿主生殖有重要影响。由于内共生菌在昆虫体内的生存环境相对稳定,与自由生活的亲缘菌相比,基因组更小,其中环境信息处理可由其寄主提供(López-Madrigaletal.,2015)。因此,扶桑绵粉蚧内共生菌基因注释时环境信息处理基因所占比例也很小,仅占1.03%。
本研究共预测了36505个基因,而注释的基因相对较少(KEGG,776个;COG,398个)。这可能与研究目标和选取的数据库有关。而注释结果出现B.ubonensis等种类,可能与基因和物种注释中存在的偏好性有关。在基因和物种注释时,本研究选取E值<0.00001的object序列作为该基因或者物种的注释信息,因此存在一定的偏好性,注释丰度较低的基因和物种可能与实际有一定差异。
昆虫内共生菌能为宿主昆虫提供营养物质、消化食物、解毒及提高免疫活性等功能,还影响宿主的生殖方式、适合度、运动行为及抗药性(魏晓莹等,2019)。本研究借助高通量测序和生物信息学技术,初步确定了扶桑绵粉蚧内共生菌菌群的群落结构和优势菌群,并初步分析了这些内共生菌的功能基因情况。但这些内共生菌对扶桑绵粉蚧的生物学特性有哪些影响、在其体内的分布以及与其入侵的关系还需进一步研究。
