石油污染土壤多酚氧化酶的动力学及热力学特征
2020-11-25冯程程赵境怡岳中辉
罗 慧, 冯程程, 赵境怡, 岳中辉
哈尔滨师范大学生命科学与技术学院, 黑龙江省普通高等学校植物生物学重点实验室, 黑龙江 哈尔滨 150025
土壤多酚氧化酶(polyphenol oxidase, PPO)是土壤中一类重要的氧化还原酶,能参与芳香族类化合物的分解转化过程,将土壤中芳香族化合物氧化成醌[1],因其对土壤环境的变化较为敏感,学者们常将其作为土壤环境生态恢复的监测指标[2-4]. 石油污染后土壤酶活性变化的研究发现,土壤酶能够迅速反映出污染程度[5-6],且利用植物对石油污染进行修复时,土壤酶也随之发生相应变化[7]. 石油污染后土壤多酚氧化酶的变化主要受石油污染物的浓度和持续时间以及修复植被或修复微生物种类的影响,如王金成等[8]研究表明,石油污泥原位修复25~80 d时,土壤多酚氧化酶活性呈下降趋势;李政等[9-10]研究认为,施用微生物菌剂修复石油污染会增加土壤多酚氧化酶活性;而陈凯丽等[11]研究发现,微生物修复后土壤多酚氧化酶活性在第2周达到最大,之后降低;朱凡等[12]研究了不同树种修复PAHs(多环芳烃)污染土壤后多酚氧化酶活性的变化,结果表明酶活性在处理9个月时升高随后又降低,PAHs含量为10 gkg时会促进酶活性,而当其含量达到50 gkg时,表现为随时间延长,酶活性被抑制,不同树木种类对酶活性的影响没有差异;而王洪等[13]研究认为,植物(黑麦草、苜蓿)修复、微生物修复及二者联合修复均促进了土壤多酚氧化酶活性,其促进程度表现为植物+微生物菌剂>植物>菌剂;王娇娇等[14]发现,甜菜与牧草间作修复PAHs污染土壤后,土壤多酚氧化酶活性提高;还有研究[13,15-16]表明,多酚氧化酶活性与石油降解率之间存在一定的相关关系. 以上研究从不同角度分析了石油污染对土壤多酚氧化酶活性的影响,但大多仅关注了酶活变化的结果,对土壤多酚氧化酶在石油污染土壤中的反应过程机制还较少涉及[17]. 土壤酶促反应的动力学和热力学特征是土壤酶学研究中的重要领域,对探讨土壤酶促反应机制有着重要意义[18-20],研究土壤多酚氧化酶活性的动力学和热力学特征能够判断酶促反应进行的可能性、方向和限度,有利于进一步研究多酚氧化酶在石油污染土壤中的酶促反应速度、过程和能量变化.
鉴于此,该文在已有石油污染对土壤多酚氧化酶影响研究[21-22]的基础上,以距油田工作区中心不同距离的区域代表不同石油污染程度,研究不同温度下各区域内裸地和羊草(Leymuschinensis)修复地的土壤多酚氧化酶活性变化,进而分析土壤多酚氧化酶的动力学和热力学特征,探究其在石油污染土壤中的酶促反应机制及行为特征,揭示其在石油污染土壤中的作用,以期为松嫩草地石油污染退化土壤的生物修复提供科学依据.
1 材料与方法
1.1 研究区域概况
大庆油田位于松嫩平原西部(121°36′E~126°36′E、44°00′N~48°35′N),该区属温带大陆性季风气候,年均气温4.2 ℃,春季干燥,夏季炎热,秋季凉爽,冬季严寒;年均无霜期143 d左右,年均降水量427.5 mm,蒸发量 1 635 mm;土壤类型为苏打盐碱土,pH在8.5以上,盐土和碱土呈斑状分布.
1.2 土壤样品采集与处理
对大庆油田工作区进行踏查,发现油田采油、贮存、运输及其他生产过程中会产生大量落地石油,由于土壤对石油类有机污染物具有较强的吸附截留能力,导致大部分污染物被截留在土壤表层,因此土壤表面逐渐形成不同深浅程度的石油污斑. 油田作业区有关部门已选取有代表性的草地,通过选用抗旱、耐盐碱的羊草对盐碱草地土壤石油污染进行了治理改良. 根据踏查情况,将石油工作区作为中心,分别在直径为1、5、15 km的同心圆上确定采样区域,采样区域中包括自然草地和羊草修复草地. 于2016年10月在不同距离的裸地(自然草地中)和羊草修复草地中进行6个样地的土壤样品采集工作. 6个样地均利用S型取样法,每个样地随机选取5个样点,各样点随机选取3个“亚样点”,再混合成一个“复合样点”,共30个“复合样点”,除去土壤表层植被,取0~15 cm土层土样,带回实验室剔除根系等杂物后,自然风干后过2 mm筛,用于土壤理化性质及土壤酶活性测定. 研究样地基本性质见表1.
1.3 土壤酶活性测定方法
土壤多酚氧化酶活采用邻苯三酚比色法测定[23],最适底物浓度为80 mmolL,最适温度为30 ℃. 设置浓度梯度为20、40、80、160 mmolL,设置温度梯度为10、20、30、40 ℃.
1.4 土壤酶动力学和热力学参数的计算方法
1.4.1土壤多酚氧化酶动力学参数计算
酶促反应可用Michaelis-Menten方程[24]来描述:
(1)

表1 研究样地土壤基本理化性质
式中:υ0为酶反应初速度,mg(g·h);S为底物浓度,mmolL;Vmax为酶促反应最大速度,mg(g·h);Km为米氏常数,mmolL.
采用Eadie-Hofstee法将Michaelis-Menten方程中分子、分母同时除以S,得到式(2),以υ0S为横坐标、υ0为纵坐标,做图得到的斜率和截距即分别对应Km和Vmax.
(2)
1.4.2土壤多酚氧化酶热力学参数计算
活化能Ea(kJmol)、温度系数Q10、活化自由能ΔG(kJmol)、活化焓ΔH(kJmol)、活化熵ΔS〔J(mol·K)〕的计算方法如式(3)~(7)所示.
活化能(Ea)由Arrhenius速度方程的积分式求得,以1T为横坐标、lnk(k为酶促反应速度常数)为纵坐标,做图得到斜率为-EaR,求得Ea.
(3)
Q10=e10EaRT(T+10)
(4)
(5)
ΔH=Ea-RT
(6)
(7)
式中:k为酶促反应速度常数;A为指前因子;T为热力学温度,K;R为气体常数,8.314 J(mol·K);N为阿伏加德罗常数,6.023×1023个mol;h为普朗克常数,6.626×10-34Js[24].
1.5 数据处理
采用Excel 2010软件进行数据整理,采用SAS 9.2软件进行单因素方差分析.
2 结果与分析
2.1 土壤多酚氧化酶活性的变化
距石油污染地不同距离的裸地和羊草修复地土壤多酚氧化酶活性变化情况如表2所示. 由表2可见,随着温度的升高和底物浓度的增加,各样地土壤多酚氧化酶活性逐渐增加(P<0.05),在30 ℃或40 ℃、底物浓度为80 mmolL或160 mmolL时达到最大值. 同一温度下,裸地土壤多酚氧化酶活性高于羊草修复地(P<0.05),其中以距油田工作区5 km处裸地(BMP)的酶活性最大;羊草修复地在10 ℃和20 ℃时,表现为距油田工作区15 km处(LRP)的酶活性最大,30 ℃和40 ℃时表现为距油田工作区1 km处(LVP)的酶活性最大.
2.2 土壤多酚氧化酶动力学特征的变化
2.2.1土壤多酚氧化酶酶促反应的Km
Km为米氏常数,是反映酶与底物结合能力的指标,其值越小,表明酶与底物的亲和力越高,酶与底物越容易结合. 由图1(a)可知,随温度升高,距油田工作区5 km处(BMP)、15 km处裸地(BRP)的Km逐渐降低,而距油田工作区1 km处裸地(BVP)的Km先降低(20 ℃时)后升高;距油田工作区不同距离羊草修复地的Km均随温度升高呈先升高后降低的趋势,在30 ℃时达到最大值. 在10 ℃时,羊草修复地Km小于同距离裸地,而在20、30、40 ℃时,同距离样地间Km的变化没有规律.
2.2.2土壤多酚氧化酶酶促反应的Vmax
Vmax为酶促反应最大速度,用来表征酶-底物复合体分解为产物和酶分子的能力,同时也能反映酶的总量,其值越大,表明酶促反应速度越快. 由图1(b)可知,随温度升高,土壤多酚氧化酶的Vmax逐渐增大(P<0.05),在30 ℃或40 ℃时达到最大值;各温度下,土壤多酚氧化酶的Vmax均表现为距油田工作区5 km 处裸地(BMP)最大、距油田工作区5 km处羊草修复地(LMP)最小(P<0.05).
2.2.3土壤多酚氧化酶酶促反应的VmaxKm
VmaxKm用于表征酶的催化性能,反映酶催化反应的速率,其值越大,表明酶促反应速度越快. 由图1(c)可知,随温度升高,土壤多酚氧化酶的VmaxKm呈逐渐增加趋势(P<0.05),在40 ℃时达到最大值,但距油田工作区1 km处裸地(BVP)土壤多酚氧化酶的VmaxKm先增后降,在20 ℃时达到最大值. 在10 ℃和20 ℃时,距油田工作区1 km处(LVP)、15 km处羊草修复地(LRP)和裸地(BVP、BRP)的VmaxKm高于5 km 处的样地(LMP、BMP)(P<0.05);在30 ℃和40 ℃时,距油田工作区5 km处裸地(BMP)的VmaxKm最大,显著大于其他样地(P<0.05).
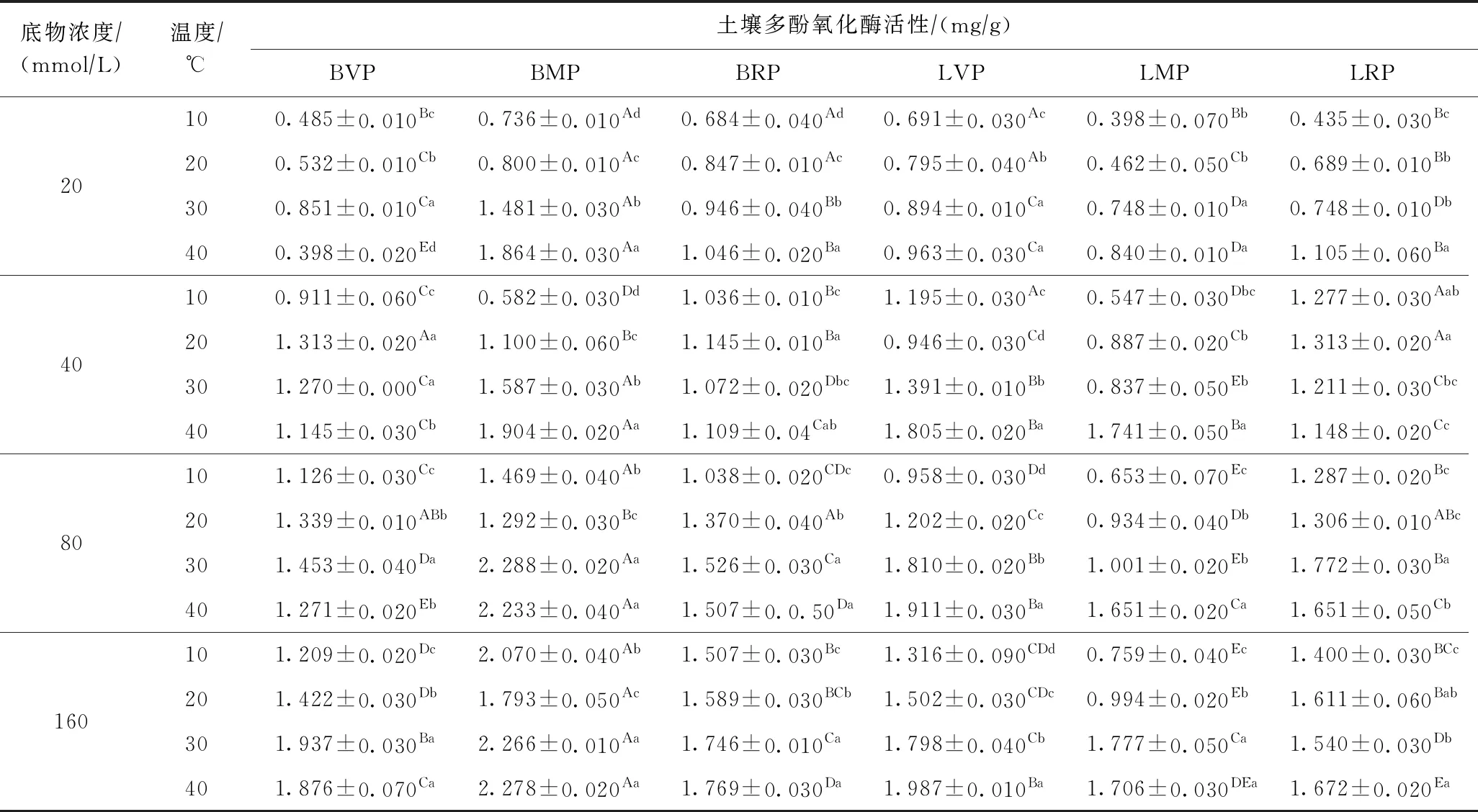
表2 不同温度、不同底物浓度下土壤多酚氧化酶活性的变化情况
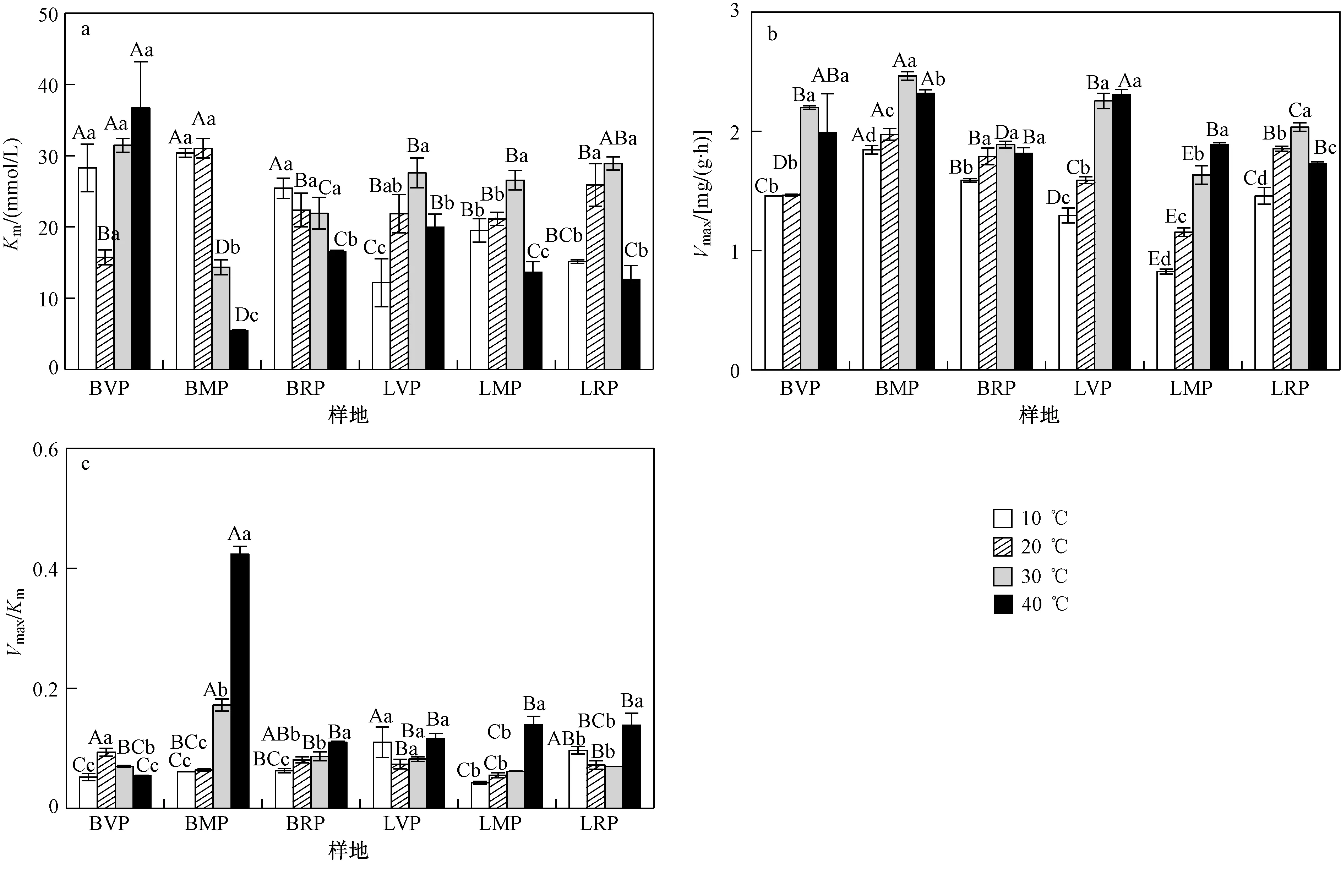
注: 不同大写字母表示同一温度不同样地间土壤多酚氧化酶的Km、Vmax、VmaxKm差异显著(P<0.05);不同小写字母表示同一样地不同温度间土壤多酚氧化酶的Km、Vmax、VmaxKm差异显著(P<0.05).
2.3 土壤多酚氧化酶热力学特征的变化
2.3.1土壤多酚氧化酶酶促反应的Ea和Q10
Ea是土壤酶促反应活化能,用来表征酶促反应进行所需要的反应能障,Ea越大,表明发生反应所需的能障越大,反应速度越慢;Q10为温度每变化10 ℃时酶活性的变化值. 由表3可知,土壤多酚氧化酶的Q10在距油田工作区1 km处裸地(BVP)、1 km处羊草修复地(LVP)随温度升高而逐渐降低(P<0.05),而在其他样地随温度变化的差异不显著. 在不同距离裸地和羊草修复地下,Q10和Ea均表现为距石油工作区1 km处裸地(BVP)最高,5 km处(BMP)、15 km处(BRP)较1km处(BVP)降低(P<0.05),而距石油工作区1 km处羊草修复地最低(LVP),5 km处(LMP)、15 km处(LRP)羊草修复地较1 km处(LVP)增加(P<0.05).
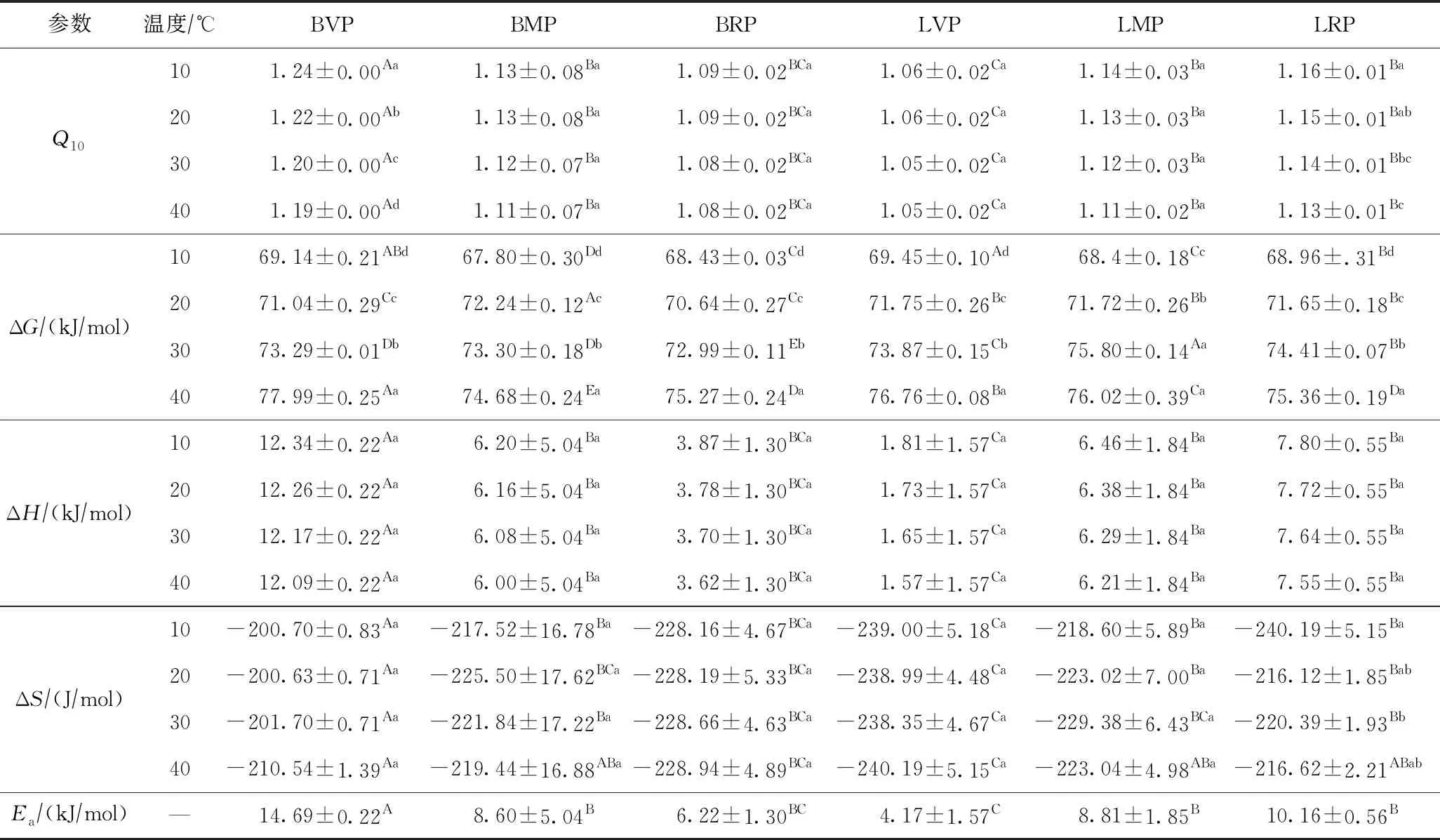
表3 不同温度下各样地土壤多酚氧化酶热力学参数
2.3.2土壤多酚氧化酶酶促反应的ΔG、ΔH、ΔS
ΔG是酶促反应活化自由能,用来反映反应物达到过渡态时所需要的自由能;ΔH是酶促反应活化焓,通常用以表征使酶的活性部位与反应物发生互补时需要从外界所获取的能量;ΔS是酶促反应活化熵,能反映体系无序性量度复合物形成过程中的结构变化特征及程度. 土壤多酚氧化酶的ΔG、ΔH、ΔS如表3所示,由表3可知,随温度升高,土壤多酚氧化酶的ΔG逐渐增大(P<0.05),而ΔH、ΔS随温度变化没有显著差异. 同一温度下,ΔG在各样地间的变化没有规律性;ΔH、ΔS与Q10和Ea的变化规律相一致.
3 讨论
3.1 石油污染对土壤多酚氧化酶动力学及热力学特征的影响
土壤多酚氧化酶活性对石油污染的响应大多表现为激活效应[25-26],该研究发现,石油污染裸地土壤多酚氧化酶活性高于羊草修复地,说明石油污染在一定程度上促进了土壤多酚氧化酶活性,同时在裸地中距油田工作区5 km处(BMP)酶活性最大,即随石油污染程度的降低,酶活性表现为先升高后降低,这与以往研究结果[26]基本一致. 究其原因:由于距油田工作区5 km处裸地(BMP)多酚氧化酶的Vmax、VmaxKm大于1 km(BVP)和15 km处裸地(BRP),Vmax、VmaxKm增加,土壤多酚氧化酶与底物结合能力较强,酶含量增多,酶催化反应中酶-底物复合体的分解比形成更容易进行,酶促反应的潜势较高,酶促反应速度加快,酶催化性能较好,酶与石油污染物充分反应,从而加快石油的降解过程,石油降解后的物质又被土壤微生物利用,作为微生物繁殖的营养来源和多酚氧化酶反应的基质,提高多酚氧化酶总量[9,17],进一步增强了酶活性. 但当靠近石油工作区(石油烃含量为5.3%~7.5%)[27]时,石油污染加重,表现为距石油区1 km处(BVP)的多酚氧化酶活性、Vmax、VmaxKm降低,这是因为石油中的有毒物质抑制了微生物的合成与分泌,土壤多酚氧化酶与底物结合能力下降,酶催化反应中酶-底物复合体的形成强度较大,酶促反应的潜势较低,酶促反应速度减慢,从而降低了酶的活性[28]. 因此通过动力学特征的变化(酶促反应作用机制)研究可以解释土壤多酚氧化酶在中度石油污染土壤(石油烃含量为5%左右)中的变化规律[27].
该研究还发现,热力学参数Q10、Ea、ΔH、ΔS的最大值均出现在距石油工作区1 km处裸地(BVP),5 km 处(BMP)、15 km处(BRP)降低,说明随石油污染程度的降低,土壤中多酚氧化酶酶促反应发生的能障较低,所需吸收外界能量较小,反应进行的方向性和强度较大,酶底物复合物形成过程中的有序性较高,能够促进土壤多酚氧化酶与底物复合体的快速合成,增加土壤多酚氧化酶量,加速裸地中土壤多酚氧化酶酶促反应速度及催化效率,从而使酶活性增大,这也从酶促反应过程中能量变化的角度解释了石油污染对土壤多酚氧化酶的影响. 综合来看,石油污染后,盐碱土壤多酚氧化酶活性的增加是通过动力学和热力学特征的变化来调控的,但其调控与石油污染浓度的关系有待做进一步深入探讨.
3.2 升温和植物修复对土壤多酚氧化酶动力学及热力学特征的影响
温度对土壤酶活性具有直接作用,是影响土壤酶活性的重要环境因子. 一般而言,在一定温度范围内土壤酶活性随着温度的升高而增加[29],刘霜等[30]研究发现,随温度升高,长白山土壤中β-1,4-葡萄糖苷酶和β-1,4-N-乙酰葡糖氨糖苷酶的Km降低,VmaxKm增加;鼎湖山土壤中β-1,4-葡萄糖苷酶和β-1,4-N-乙酰葡糖氨糖苷酶的Vmax、VmaxKm增加. ZHANG等[31]发现,温度升高会促进东北地区4种土壤中脲酶、磷酸酶、芳基硫酸酯酶的动力学特征,降低土壤酶热力学特征. 该研究发现,随温度升高,石油污染土壤中多酚氧化酶的Km逐渐减小,Vmax、VmaxKm逐渐增大,ΔH、ΔS呈逐渐减小的变化趋势,这与以往的研究结果[30-31]基本一致,说明温度升高(30 ℃、40 ℃)能加快石油污染土壤中多酚氧化酶分子运动,促使酶与底物进一步结合,降低土壤多酚氧化酶酶促反应在石油污染土壤中的反应能量,增大酶活性,促进酶反应的催化作用,提高酶-底物过渡态的有序性,利于向生成产物的方向转化,使土壤多酚氧化酶在石油污染土壤中更容易与酚类物质结合进行催化反应及物质循环过程. 另外,土壤多酚氧化酶的Q10在距油田工作区1 km 处裸地(BVP)和羊草修复地(LVP)随温度升高而逐渐降低,说明土壤多酚氧化酶酶促反应在低温时对温度的敏感性较大,而随温度升高,酶促反应速度对温度的敏感性下降,升温提供的热量在酶的热变性过程中消耗较少,酶活性相应增加较多,这与German等[32]研究土壤β-酶葡萄糖苷酶活性的Q10与年均温呈负相关的结果相互印证.
同时该研究还发现,从低温(10 ℃和20 ℃)到最适温度(30 ℃)或高温(40 ℃)的变化过程中,羊草修复地土壤多酚氧化酶活性、Vmax、VmaxKm都有升高趋势,Ea、ΔG呈降低趋势,这可以认为是羊草修复后升温加速了石油污染土壤中多酚氧化酶酶促反应的进行,提高了多酚氧化酶的生物反应速率,反应所需的活化能和自由能降低,酶促反应过程中的能障减小,反应自发程度提高,促进了土壤多酚氧化酶在石油污染盐碱草地中发挥其作用. 但羊草修复地土壤多酚氧化酶活性、Vmax、VmaxKm大都小于裸地,这可能是因为羊草有一定的吸附石油的作用,可以在一定程度上“补偿”多酚氧化酶的作用;而羊草修复后的Ea、ΔG、ΔH、ΔS均在距石油工作区1 km处(LVP)最低,低于裸地,说明石油污染羊草修复后反应发生的能障较低,反应过程中分子构型变化所需的力减少,易于构型的改变,所需吸收外界能量较小,酶-底物复合体过渡态有序性较高,能够促进土壤多酚氧化酶与底物复合体的快速合成,增加土壤多酚氧化酶量,加速裸地中土壤多酚氧化酶促反应速度及催化效率,从而使酶活性增大,这对提高植物修复石油污染物的研究有一定积极意义[33].
4 结论
a) 不同含油量石油污染裸地和羊草修复地的土壤多酚氧化酶活性及动力学参数Vmax和VmaxKm在10~40 ℃温度范围内随温度升高而逐渐增大,在30 ℃或40 ℃时达到最大;在同一温度下,石油污染裸地土壤多酚氧化酶活性高于羊草修复地,且在含油量为5%左右的中等石油污染裸地土壤中酶活性最高,Km和VmaxKm在各样地间无规律性变化,Vmax表现为距石油工作区5 km处裸地最大、羊草修复地最小.
b) 不同含油量石油污染裸地和羊草修复地多酚氧化酶的热力学参数Q10、ΔH、ΔS在10~40 ℃温度范围内随温度升高的差异不显著,ΔG呈逐渐增加趋势;在同一温度下,Q10、Ea、ΔH、ΔS均表现为距石油工作区1 km处裸地最大、羊草修复地最小.
