铜铅胁迫下水分、温度和施肥对黑麦草富集能力的影响
2020-11-24董馨岚李铭红朱剑飞
董馨岚, 白 哲, 李铭红, 朱剑飞
1 浙江师范大学行知学院,金华 321000 2 浙江师范大学生态研究所,金华 321000
随着工农业生产、城镇化的迅猛发展,工业废物排放和农业化学品的使用正使大量的重金属进入土壤环境,造成土壤污染日趋严重[1]。铅是具有高度毒害性的重金属,与镉、铬、汞、砷合称为“五毒”[2]。矿山开采、金属冶炼、汽车废气是环境中铅的主要来源,而含铅废水的灌溉则是造成农田铅污染的首要原因。铜是植物生长发育过程中的必需元素,近年来逐渐增多的铜矿开采、含铜杀菌剂的频繁使用以及含铜垃圾的不合理排放等成为土壤中铜的主要来源[3]。当土壤含铜量超过一定阈值时,不仅会导致土壤环境质量下降, 还会对土壤中的动、植物产生严重胁迫[4- 5]。据2014年全国土壤污染状况调查公报显示,在调查范围内,铜(Cu)、铅(Pb)重金属的超标率分别达到2.1%、1.5%,是耕地、工业废弃地和工业园区等地的主要污染物。倪中应等[6]以杭州市域范围内27个重金属污染农田土壤为样本,测得土壤中Cu、Pb含量分别在7.65—113.65 mg/kg和5.69—54.25 mg/kg范围间, 其中有机质结合态和氧化物结合态总比例较高, 具有中等程度的生态污染风险。杨清伟等[7]对利用铅锌矿废水灌溉约50年的农田进行调查,结果表明,土壤中Cu、Pb的含量远超背景值,高达1486、680 mg/kg,污染严重。
目前,国内外土壤重金属污染修复技术主要包括:工程技术措施、物理修复、化学修复、生物修复以及农业生态修复[8]。其中,植物修复由于其低成本、无二次污染等优势成为土壤生物修复研究中的热点,而黑麦草(LoliumperenneL.)作为我国南方普遍种植的一种优良牧草,近年来因其生长快,生物量高等特点得到关注。已有研究表明,黑麦草对重金属Cu、Pb具有一定富集能力。张永兰和王友保[9]在黑麦草对铜尾矿的修复研究中发现多年生黑麦草的生长和吸收作用可促进土壤中铜的活化,从而降低土壤铜含量,有效推动铜尾矿基质的修复进程。宣斌等[10]对黑麦草等12种植物的Pb累积特性进行了比较分析,认为黑麦草具有良好的铅修复潜力,可作为铅锌矿区耐性栽培植物。而针对植物富集过程中生物量下降、生长速率低下等诸多限制因素[11],农业生态修复技术所涵盖的农艺修复和生态修复两方面措施可在一定程度上提高植物修复效率。相关研究表明,土壤水分影响作物的生长,并间接影响重金属在土壤中的迁移和分配[12]。温度除了对植物的蒸腾作用、光合作用等各项新陈代谢活动产生一定的影响作用外[13-14],还会影响植物对重金属离子的吸附,如Chao[15]等发现高温预热能在一定程度上降低Cd的毒性。同样,化肥的施用也是改善土壤理化性质,影响株高、茎粗、叶绿素等生理指标,使植物对重金属的吸收富集能力产生变化的关键因素。
因此,本研究以常见牧草黑麦草为供试植物,探究其在不同水分、温度及施肥条件的交互作用下对Cu、Pb单一及复合污染的响应情况,从而筛选出利于黑麦草吸收Cu、Pb的环境因子组合,进一步拓宽黑麦草对重金属Cu、Pb的修复潜力。
1 材料与方法
1.1 供试材料
黑麦草种子购于浙江金华。采用校园周边农田土壤,取地表至20 cm深度土,去杂后置于阴凉通风处自然风干,研磨并过筛(孔径:4 mm)备用。测得原始土壤中重金属Cu、Pb的背景值分别为35.94、78.22 mg/kg,土壤pH值为6.8,有机质5.13 g/kg, 全氮0.72 g/kg, 速效磷18.6 mg/kg, 速效钾203.1 mg/kg。试验以分析纯CuCl2·2H2O和Pb(NO3)2提供所需重金属浓度,根据国家环境土壤质量标准(pH>6.5,三级标准),设置单一Cu、Pb污染浓度为400 mg/kg、500 mg/kg,并组成复合Cu-Pb污染浓度。
1.2 试验设计
在已有单一环境因子对黑麦草富集Cu、Pb能力影响的试验中,曾分别设置不同持水量梯度(40%、50%、60%、70%、80%、90%、100%WHC)、不同温度梯度(5℃、10℃、15℃、20℃、25℃、30℃、35℃)、不同肥料梯度(N、P、K、NP、NK、PK、NPK)进行单一环境因子影响探究。根据试验结果及实际生产可行性选取变量梯度进行交互作用参数设置,如表1所示,所得处理组合列于表2。
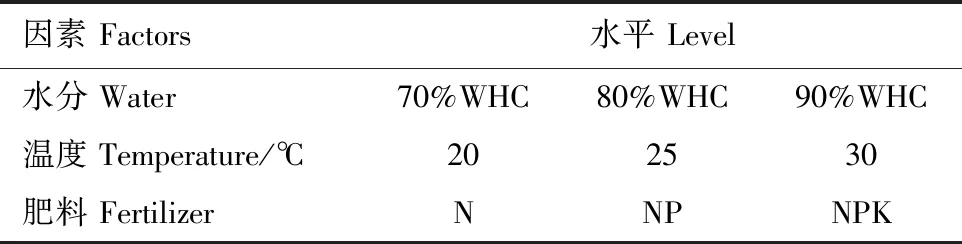
表1 试验设计
挑选籽粒饱满、大小基本一致的优质牧草种子,70%的酒精消毒3 min后用去离子水洗净吸干备用。用高15 cm,直径20 cm的花盆装取1.5 kg土壤,每盆播种种子30颗,每处理重复3次,盆栽数量总计81盆。试验所设置的水分变量指水分占土壤重量的百分比,通过称重法[16]进行控制,定时(5 d)割取各处理植株用以校正幼苗生长对土壤水分变量所造成的误差;通过光照培养箱恒温培养得到各温度变量;通过尿素(46%N)、过磷酸钙(12%P2O5)、氯化钾 (60%K2O)来提供所需N、P、K肥,参照占丽平等[17]研究进行配施。控制其他无关变量保持一致,开始60 d后收获植物和根际周围土样。
1.3 样品分析
取植物样,先用自来水充分淋洗,0.1 mol/L稀盐酸洗净后再用去离子水淋洗2—3次,洗净后用滤纸吸干,称量植株鲜重。将植株地上部与根部分开,105℃下杀青30分钟,90℃烘干至恒重后称量干重,研磨并过60目筛。取植物根部土壤,90℃下烘至恒重,研磨至粉末状后过100目筛。植物样品采用HNO3-HClO4法进行消解,土壤样品采用HCl-HNO3-HClO4法进行消解。最后,采用电子耦合等离子发射光谱法(Inductively Coupledd plasma-Atomic Emission Spectrometry, ICP-AES)测定样品中Cu、Pb含量。
1.4 数据处理
(1)生物富集系数(bioconcentration factor,BCF):植物地上部或根部重金属含量/土壤中重金属含量
(2)转运系数(transfer coefficient,TF):植物地上部重金属含量/植物根部重金属含量
试验所得数据采用Excel 2010、SPSS 20.0等软件进行统计分析处理及绘图。通过SPSS单因素方差分析ANOVA统计分析后,Duncan多重比较法检验各处理间均值差异显著性,一般线性模型(GLM)比较水分、温度与施肥处理之间的二因素、三因素交互作用。
2 结果分析
2.1 Cu、Pb胁迫下环境因子交互对黑麦草生物量的影响
由图1可知,在不同的水分、温度及养分供给条件下,受Cu单一胁迫时,黑麦草在90%WHC、30℃及施NPK(27)时,可获得最大茎叶干重及总生物量(P<0.05),而在90%WHC、25℃、施N(12)时植物根的生长最佳。受Pb单一胁迫时,黑麦草在90%WHC、30℃、施NP(24)时获得最大的茎叶干重,且施N或NPK也能达到较好的生长效果;在90%WHC、25℃、施NPK(18)时获得最大的根干重,在90%WHC、30℃、施N(21)时整体长势最佳(P<0.05),显著优于其他处理组。
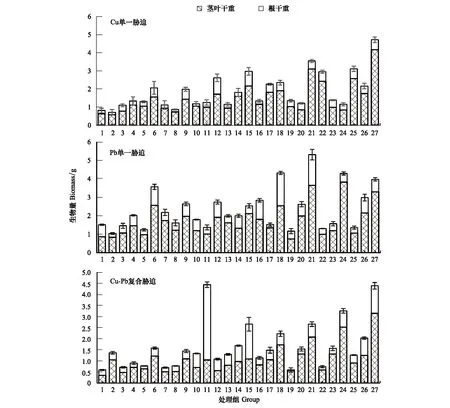
图1 Cu、Pb单一及复合胁迫下不同环境因子对黑麦草生物量的影响Fig.1 The biomass of Lolium perenne L. at (three levels of water,temperature,fertilizer) (different environmental factors) under single and compound stresses of Cu and Pb
当受到Cu-Pb复合胁迫时,黑麦草地上部分的生长与Cu、Pb单一胁迫时相比存在明显的抑制作用,但仍在90%WHC、30℃及施NPK(27)时取得茎叶干重最大值,根干重在80%WHC、25℃及施N(11)处理时显著高于其他处理组(P<0.05),且取得总生物量最大值。
2.2 Cu、Pb胁迫下环境因子交互对黑麦草Cu、Pb富集量的影响
由图2可以看出,在不同环境因子组合下,黑麦草不同部位对Cu的吸收量有较大差异,大体上体现为根部富集量大于地上部。在80%WHC、25℃及施N(11)时,黑麦草地上部在Cu单一胁迫下具有最大富集量444.49 mg/kg,而在Cu-Pb复合胁迫时,黑麦草在70%WHC、25℃及施NPK(16)时对土壤中Cu的富集量显著提高,具有所有处理中的最大地上富集量465.18 mg/kg。黑麦草根部在Cu单一胁迫下,在90%WHC、25℃及施NPK(18)时具有最大富集量1343.1 mg/kg,而在Cu-Pb复合胁迫下,在80%WHC、30℃及施NP(23)时才取得最大富集量1274.78 mg/kg。
图2 Cu、Pb单一及复合胁迫下不同环境因子对黑麦草Cu富集量的影响Fig.2 The Cu enrichment in shoots and roots of Lolium perenne L. at (three levels of water,temperature,fertilizer) (different environmental factors) under single and compound stresses of Cu and Pb
由图3可以看出,在不同环境因子组合下,黑麦草不同部位对Pb的吸收量有较大差异,但与Cu富集规律类似,也体现为根部富集量大于地上部。在80%WHC、30℃及施NPK(17)时,黑麦草地上部在Pb单一胁迫下具有最大富集量557.46 mg/kg,而在Cu-Pb复合胁迫下,黑麦草在80%WHC、30℃及施N(20)时对土壤中Pb的富集量显著提高,具有所有处理中的最大地上富集量597.66 mg/kg。黑麦草根部在Pb单一胁迫下,在90%WHC、25℃及施NP(15)时具有最大富集量1154.62 mg/kg,而在Cu-Pb复合胁迫下,在80%WHC、30℃及施NP(12)时才取得最大富集量1496.83 mg/kg。
根据不同环境因子将27组试验进行分类统计,结果如下:在Cu单一污染时,黑麦草地上部分别在温度25℃(266.76 mg/kg)、持水量80%(167.44 mg/kg)、施NP(166.99 mg/kg)时,根部分别在30℃(825.43 mg/kg)、持水量80%(854.78 mg/kg)、施N(768.00 mg/kg)时具有最大富集量。在Pb单一污染时,黑麦草地上部分别在温度30℃(316.19 mg/kg)、持水量90%(261.18 mg/kg)、施NPK(331.58 mg/kg)时,根部分别在25℃(806.27 mg/kg)、持水量90%(697.24 mg/kg)、施NPK(878.84 mg/kg)时富集效果最佳。在Cu-Pb复合污染时,黑麦草地上部分别在温度25℃(204.68 mg/kg)、持水量70%(208.09 mg/kg)、施NP(188.20 mg/kg)时对Cu的富集量最大,在温度25℃(410.79 mg/kg)、持水量80%(349.17 mg/kg)、施NP(388.81 mg/kg)时对Pb的富集量最大;根部分别在30℃(1021.82 mg/kg)、持水量70%(903.57 mg/kg)、施N(918.27 mg/kg)时对Cu的富集效果最佳,在30℃(1161.63 mg/kg)、持水量80%(1307.28 mg/kg)、施N(1182.54 mg/kg)时对Pb的富集效果最佳。数据表明,在应对不同的重金属胁迫时,黑麦草对温度、水分及肥料的最适范围表现出差异性,当三者共同作用时,还会产生交互影响,形成最适处理组合。
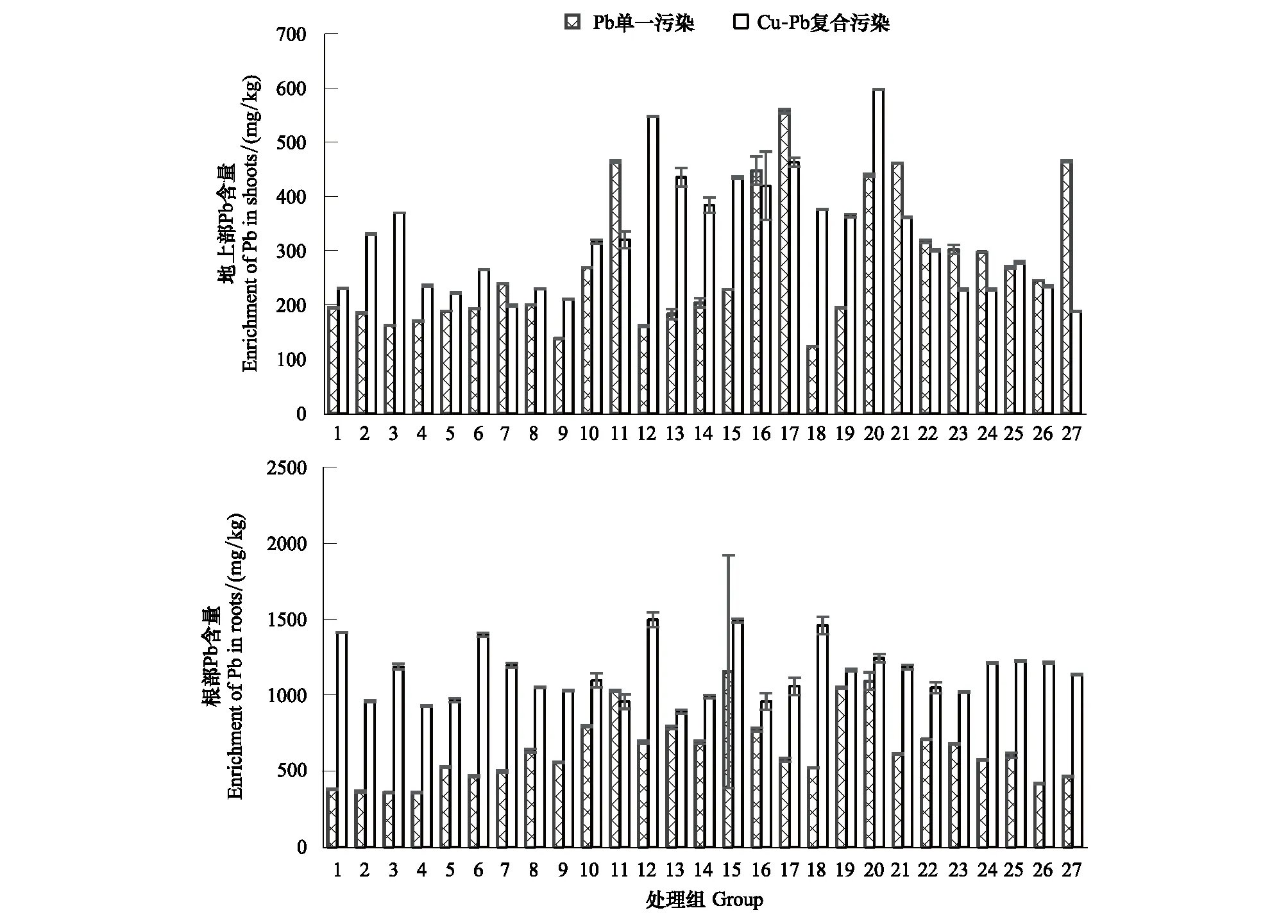
图3 Cu、Pb单一及复合胁迫下不同环境因子对黑麦草Pb富集量的影响Fig.3 The Pb enrichment in shoots and roots of Lolium perenne L. at (three levels of water,temperature,fertilizer) (different environmental factors) under single and compound stresses of Cu and Pb
表3—4为不同环境因子处理对植物富集Cu、Pb含量的影响所进行的方差分析,结果显示:在Cu、Pb单一胁迫条件下,黑麦草的根部Cu富集量及地上部Pb富集量受到水分、温度、肥料以及其二因素、三因素交互作用的极显著影响(P<0.01);而黑麦草地上部对Cu的富集量仅表现出受水分、温度的极显著影响,同时,水分和温度以及温度和肥料之间也表现出极显著的交互作用;黑麦草根部对Pb的富集则受水分和温度交互以及三因素交互的显著影响(P<0.05),其他因素影响极显著。在Cu、Pb复合胁迫条件下,除肥料对地上部Cu富集量、水分和温度的交互作用对根部Cu富集量无显著影响外,其他各因素及其交互作用均表现出对黑麦草富集Cu、Pb含量的极显著影响。

表3 单一胁迫下不同环境因子处理及其组合对黑麦草富集Cu、Pb含量影响的方差分析

表4 复合胁迫下不同环境因子处理及其组合对黑麦草富集Cu、Pb含量影响的方差分析
2.3 Cu、Pb胁迫下环境因子交互对黑麦草Cu、Pb富集及转运系数的影响
由表5可以看出,在不同环境因子组合下,黑麦草的地上部和根部对Cu的富集系数有较大差异,基本体现为地上部BCF(Cu)<1,根部BCF(Cu)>1。分析Cu富集系数可知,在25℃、施NP时,黑麦草地上部在Cu单一污染下富集能力较强,而在Cu-Pb复合污染时,地上部在相同温度施NPK时BCF(Cu)最高;在20℃及30℃施N、NP条件下,复合污染处理在不同程度上提升了地上部BCF(Cu)。在90%WHC、25℃及施NPK时,黑麦草根部在Cu单一污染下取得BCF(Cu)max,而在Cu-Pb复合污染时,根部在20℃的相同水分和养分条件下富集系数最高;在不同温度和水分,施NP、NPK肥条件下,复合污染处理在不同程度上提升了根部BCF(Cu)。

表5 Cu、Pb单一及复合胁迫下不同环境因子组合时黑麦草对Cu、Pb的富集系数
在不同环境因子组合下,黑麦草的地上部和根部对Pb的富集系数高于Cu,且在相对适宜的环境条件下,存在地上部BCF(Pb)>1。分析Pb富集系数可知,在90%WHC、30℃及施NPK时,黑麦草地上部在Pb单一污染下富集系数最大,而在Cu-Pb复合污染时,地上部则在相同温度,80%WHC及施N时具有最大值且高于单一污染时系数;在20℃及25℃施N、NP条件下,复合污染处理在不同程度上提升了地上部对BCF(Pb)。在90%WHC、25℃及施NP时,黑麦草根部在Pb单一污染下取得BCF(Pb)max,而在Cu-Pb复合污染时,根部相同温度和水分,施NPK条件下富集系数最高;此外,复合污染处理均在不同程度上提升了根部BCF(Pb)。
对水分、温度和肥料在铜铅胁迫下对黑麦草富集系数的影响进行单因素及多因素方差分析,结果显示:在Cu单一胁迫条件下,黑麦草的地上部BCF(Cu)受到水分、温度、肥料以及水分与温度组合、温度与肥料组合交互作用的极显著影响(P<0.01),而根部BCF(Cu)均受到3种环境因子及其二因素、三因素交互作用的显著(P<0.05)或极显著影响;当受到Cu-Pb复合胁迫时,各因子及其多因素交互作用除温度、肥料因子对地上部BCF(Cu),水分与温度组合对根部BCF(Cu)无显著影响外,均对黑麦草地上部及根部BCF(Cu)有显著或极显著影响。而黑麦草BCF(Pb)除在单一胁迫时受水分与肥料互作不显著外,均受到各因子及其多因素交互作用的显著或极显著影响。
表6为黑麦草在不同环境条件及单一、复合污染处理下对Cu、Pb的转运系数。结果表明,仅在90%WHC、30℃及施NPK时,黑麦草TF(Pb)最大为1.002>1,其余各环境因子组合下TF(Pb)<1。在20℃、施N肥各水分处理,施NP肥90%WHC水分处理,施NPK肥70%WHC水分处理及25℃、施NPK肥70、80%WHC水分处理;30℃、施N肥70、90%WHC水分处理,施NP肥70%WHC水分处理共10组处理组合中,复合污染处理在不同程度上提高了黑麦草的TF(Cu)。此外,复合胁迫也在25℃、施N肥90%WHC水分处理,施NP肥各水分处理,施NPK肥90%WHC水分处理及30℃、施N肥70、80%WHC水分处理共7组处理组合中提高了TF(Pb),其余均表现为降低作用。

表6 Cu、Pb单一及复合胁迫下不同环境因子组合时黑麦草对Cu、Pb的转运系数
通过三种环境因子单因素及多因素方差分析结果可知:在单一Cu胁迫下,黑麦草TF(Cu)受到肥料的显著影响(P<0.05),水分、温度以及各二因素、三因素交互作用的极显著影响(P<0.01),而在复合胁迫下,TF(Cu)仅表现出受水分及三因素两两组合间交互作用的极显著影响。在单一Pb胁迫下,水分、温度及肥料对黑麦草TF(Pb)无显著作用,而水分与温度组合、温度与肥料组合及三因素交互作用对其TF(Pb)的变化起到显著或极显著的作用;在复合Cu-Pb胁迫条件下,黑麦草TF(Pb)受到各环境因子及其交互作用的极显著影响。
3 讨论
黑麦草作为新型修复型牧草,对重金属镉、铜、铅、锌均有一定的富集能力。黑麦草对Cu、Pb的富集性能也已经开展了多方面的研究。如刘大林等[18]、陈鸣晖等[19]探究了黑麦草的不同品种及植株的不同部位对Cu耐受能力的影响。刘明美等[20]研究发现,不同的Pb2+处理浓度对黑麦草种子萌发和幼苗生长的作用不同,表现为低浓度促进,高浓度抑制(以800 mg/kg为界),且对根的抑制作用强于芽。胡伟等[21]研究表明,黑麦草在Pb胁迫土壤中的生长促进了残渣态铅向可氧化态及可还原态铅的转化,使铅由沉淀状态不断转变为可溶态。可以发现,黑麦草对Cu、Pb的富集能力将受到植物品种、生育期、吸收部位以及重金属化合物形态等多种因素的影响,具有一定的波动范围;同时,本试验结果表明,复合重金属胁迫条件也能对黑麦草对单一Cu、Pb的富集与转移产生影响,这可能与Cu、Pb两者之间所具有的交互作用有关。除此之外,还可通过控制适宜的环境要素使植物在给定条件下将其富集性能发挥至最大限度。而多种环境因子进行交互作用正是植物生长过程中所存在的实际现象。因此,在充分研究单因子作用的基础上再进一步探究各个因子之间的作用具有意义。
水分不仅是植物新陈代谢的溶剂,运输营养物质的载体,还能通过影响土壤pH、Eh、有机质和碳酸钙含量等土壤化学生物特性[22],间接引起重金属在土壤中分配及迁移的改变,并进一步影响植物生物量及重金属含量[23- 25]。Angle等[25]研究得到,遏蓝菜等超富集植物叶片中的重金属含量在土壤80%或100%WHC下会得到提高。而对于伴矿景天来说,茎叶Cd富集量随土壤水分的增大先升高后降低,并在70%WHC时取得最大值[23]。本试验中,黑麦草在80%或90%WHC下易取得较大Cu、Pb富集量,同时主要在90%WHC时具有较高生物量。这表明不同的植物具有不同的最适水分含量,对其进行针对性探究有利于在实际应用中合理提供水分供给。
肥料作为植物营养元素补充的主要来源,在植物生长与生物量积累过程中起到重要作用,但与此同时也将不可避免地引入一些毒害成分,对土壤和植物中的重金属富集造成影响[26- 27]。常见的化肥包括氮、磷、钾肥。黑麦草一年可刈割多次,养分管理粗放,通常施用氮肥。相关研究表明,磷肥、钾肥同样能促进黑麦草对养分的吸收,增加生物量积累[28],且增产效果为N>P>K[17],这可能从植物生长角度影响植物的修复效率。而王美和李书田[29]对施肥影响重金属累积研究进展进行了大量相关文献的系统分析后,总结配施氮、磷、钾肥后土壤各金属含量在一定范围内波动,无明显升降规律,这与本试验中对养分的研究结果相类似,认为肥料中的养分不是影响黑麦草富集能力的主要单因子,如肥料对黑麦草地上部的Cu富集量无显著影响。但也要意识到,植物对不同形态重金属的吸收利用不同,其中可交换态的利用程度最大[30]。一些研究认为,氮、钾肥对土壤重金属累积的影响不大,而磷肥可与重金属形成多种形态的难溶性磷酸盐,降低土壤的污染程度[31];但也有研究表明,磷肥是造成研究所开展的试验区内土壤 Cd、Pb等重金属富集的主要原因[32]。因此,有关各种肥料及施肥方式与土壤重金属含量之间的关系还有待进一步研究。
此外,温度对植物生长和重金属转运的影响也不可忽视,不仅能通过植物酶促反应、蒸腾作用等各项生理活动直接影响植物生长发育过程[13],也将引起土壤重金属存在形态的改变。研究发现,高温下黑麦草的生长速率减慢,但植物体内游离脯氨酸等渗透调节物质显著增加,表现出一定的抵抗逆境能力;低温下相应指标则得到一定积累[33]。本试验反映出黑麦草在几个相近普适温度下的富集能力差异,未进行极端高温和低温的温度设置。但可以发现,黑麦草在25℃或30℃下对Cu、Pb的总体修复性能较强,20℃不利于黑麦草的生长与金属富集。
综合分析黑麦草不同部位的生长与富集情况,认为在Cu单一污染下,选择90%WHC、施NPK,30℃及25℃因子组合;在Pb单一污染下,选择90%WHC、施N或NPK,30℃因子组合;受Cu-Pb复合污染时,黑麦草的生长受到抑制作用,在90%WHC、30℃及施NPK时对Cu、Pb的综合修复作用较佳。黑麦草对Cu、Pb的富集及转运系数表明,黑麦草对Cu、Pb具有一定的富集能力,并会随着水分、温度、肥料等环境因子的改变而发生变化,但对Cu、Pb的转运能力较弱。方差分析结果显示,在水分、温度、肥料三者中,水分和温度对黑麦草Cu、Pb富集能力具有显著影响,且三因子之间存在二因素及三因素的交互作用。其中在Cu、Pb单一污染胁迫下,受到其二因素交互的显著影响;在单一及复合污染中,均在不同程度上受到三因素交互作用的显著影响。
水分、温度和肥料是植物生长过程中的重要环境因子,在重金属污染土壤条件下,它们自身以及组合所引起的变化均会直接影响黑麦草的生长发育。相关研究表明,水分肥料互作能显著影响植物对水分的利用率及产量[34],其机理主要是通过促进植物的根系发育[35]。温度与土壤温度、湿度分别呈正相关、负相关关系,根系的土壤温度影响植株对水分的吸收。肖自添等[36]研究表明,植株根系的吸收养分和水分能力将在低温环境下随温度的降低而下降,此时增加施肥量会对植株造成盐胁迫。对于黑麦草富集能力的影响,通过总结前人研究成果及本试验结果,认为可能通过:(1)对黑麦草生长发育过程的影响间接影响富集能力。修复植物的生物量是衡量其富集性能的重要指标。各环境因子间存在相互影响制约的关系,温度影响水分,水分通过影响植株体内物质的转运及根系的生长,而影响整体生长情况[37]。根系吸收水分和养分,通过光合作用产生物质积累,促进植株各器官转化[38]。因此适宜的水肥及温度组合将促进植株的生长,促进生物量累积;而过高或过低的水肥及温度组合反而会导致相应指标的下降,不利于植株的正常生长。(2)对黑麦草富集能力的直接影响。在本试验研究条件下,几种环境因子之间存在的二因素及三因素交互作用,对土壤pH、粘结性等具有一定影响,从而使Cu、Pb在土壤中的存在形态发生变化,这可能引起了黑麦草Cu、Pb富集性能的变化;此外,黑麦草体内脯氨酸、可溶性糖蛋白以及丙二醛等抗性物质的含量对于水分、温度及肥料交互作用的响应,也将反映于黑麦草对Cu、Pb富集性能的改变中。目前环境因子及其交互对于黑麦草生长的影响已进行了较多研究,但关于其对黑麦草在污染土地中的生长与富集的影响的文献论述仍较少,对于黑麦草在其他不同重金属及浓度土壤中,三因子交互对其生长与富集能力的作用还需进一步研究总结。
