滇中亚高山森林植物叶-凋落叶-土壤生态化学计量特征
2020-11-24张雨鉴王克勤宋娅丽苏孟白李晓龙李学峰
张雨鉴,王克勤,宋娅丽,*,苏孟白,李晓龙,李学峰
1 西南林业大学生态与环境学院,昆明 650224 2 北京林业大学水土保持学院,北京 100083
生态化学计量学作为研究生态系统中元素稳定循环机理和能量平衡状态的理论[1- 2],为研究碳(C)、氮(N)、磷(P)元素在陆地生态系统中的相互耦合关系提供了新的思路和方法[3- 5]。森林作为陆地生态系统中结构相对完整、组成相对复杂的生态系统,其各组分由于复杂多样的生态学过程而密切联系[6]。森林生态系统中,植物、凋落物和土壤作为森林生态系统中三个重要的组成部分,彼此相互影响形成一个连续的体系[7]。植物通过叶片光合作用积累有机质,并以微生物分解凋落物的形式释放植物中的养分补偿土壤[8- 9];凋落物作为植物与土壤间物质能量交换的枢纽,同时也是森林生态系统有机碳和养分的重要储存库[10- 11];土壤则作为植物的生存基础,主要为植物的生长发育提供充足的养分和水分。由于植物对养分的需求量和凋落物分解过程中养分的归还量以及土壤养分的供应量之间存在相互影响的关系,使得植物-凋落物-土壤体系养分含量具有显著的时空差异[12]。因此,研究C、N、P在植物叶-凋落叶-土壤体系中的化学计量特征及各组分间的相互作用关系,对于揭示森林生态系统中的养分循环规律及不同生态系统间养分平衡特征具有重要的科学意义。
目前,有关森林生态系统各组分化学计量特征已存在较多研究,McGroddy等[13]通过研究全球尺度下森林(温带针叶林、温带阔叶林和热带森林)C、N、P化学计量特征发现乔木叶和凋落叶的C、N、P比值变异性较大;Han等[14]以我国753种乔木、灌木或草本叶片为研究对象,研究发现植物叶片N和P含量随纬度的增加呈增大趋势,而N/P比值则无明显变化;谌贤等[15]研究川西亚高山森林凋落叶发现,林分类型及凋落物分解阶段对凋落物碳氮磷含量的影响较为显著;张泰东等[16]分析了帽儿山5种林型土壤碳氮磷化学计量关系,结果表明不同林型间土壤C-N耦合关系有趋同现象。由此可见,目前针对森林生态系统生态化学计量特征的研究多以单一组分为主(植物器官或凋落物或土壤等),将植物叶、凋落叶和土壤作为一个体系进行研究的报道则相对较少,且多集中在低中海拔区域,如有关子午岭林区[12]、辽东山区[17]、黄土丘陵区[7]等区域的研究,而针对云南低纬度高海拔森林区域的研究则鲜有报道。
滇中亚高山区域位于我国云贵高原、青藏高原和横断山地三大自然地理区域相结合的特殊部位,是一个复合性生态过渡带,气候由南亚热带过渡到北亚热带,植被以半湿性常绿阔叶林为主,物种丰富[18]。针对该区域森林生态系统的化学计量特征进行研究对于认识该特殊气候带下森林生态系统养分分配特征和限制状况具有重要意义。因此,本文以地处该气候区域林龄相近、林相整齐、覆盖度高的5种滇中亚高山典型森林生态系统(云南松林(Pinusyunnanensisforest)、华山松林(Pinusarmandiiforest)、常绿阔叶林(Evergreen broad-leaf forest)、高山栎林(Quercussemecarpifoliaforest)以及滇油杉林(Keteleeriaevelynianaforest))为研究对象,通过测定各森林生态系统植物叶、凋落叶及土壤C、N、P含量,探究不同森林类型各组分(植物叶、凋落叶和土壤)C、N、P之间的相互关系,试图回答以下两个问题:(1)不同森林类型间“植物叶-凋落叶-土壤”C、N、P生态化学计量有何特征?这些特征对林分类型是如何响应的?(2)各森林生态系统“植物叶-凋落叶-土壤”连续体C、N、P化学计量特征有何内在关联?通过以上研究,以期为探明区域尺度森林生态系统养分元素循环规律和系统结构稳定平衡机理提供理论依据。
1 研究材料与方法
1.1 研究区概况
研究区设于云南省玉溪市新平县磨盘山森林生态系统国家定位观测研究站(23°46′18″—23°54′34″N,101°16′06″—101°16′12″E),属于中亚热带气候,是云南亚热带北部气候与亚热带南部气候的过渡地区,具有典型山地气候特征。该区海拔1260.0—2614.4 m,年均气温15℃,年降水量为1050 mm,降水主要集中在5—10月。土壤以山地红壤和玄武岩红壤为主,高海拔地区有黄棕壤。研究区作为中国亚热带地区以半湿性常绿阔叶林为主的重要原始森林区,分布有高等植物楠木(Phoebechinensis)、梭罗树(Reevesiapubescens)、树蕨(Arthropterispalisotii)等98科137属324种。该区植被类型随海拔的升高呈明显的垂直分布特征,主要分布的森林类型为亚热带常绿阔叶林、亚热带中山针阔混交林、针叶林和高山矮林(主要为高山栎林)等,其中常绿阔叶林树种组成为米槠(Castanopsiscarlesii)、光叶柯(Lithocarpusmairei)、糙皮桦(Betulautilis)、马缨杜鹃(Rhododendrondelavayi)、牛筋条(Dichotomanthestristaniicarpa)等,其中优势树种为米槠、光叶柯和糙皮桦,且约60%为米槠。
1.2 样地设计
本研究对象选取了磨盘山森林生态系统国家定位观测研究站区域内5种典型的森林群落,包括云南松林、华山松林、常绿阔叶林、高山栎林和滇油杉林。于每种森林类型选取3个立地条件相近的重复样地,每个样地设置大小为20 m×20 m,共计15个,情况如表1。

表1 5种森林类型样地基本概况
1.3 样品采集及室内分析
针对样地内的立木以林分中胸径(DBH)≥ 5 cm的林木作为起测径阶,对标准样地内所有胸径(DBH)≥ 5 cm的林木进行每木检尺,记录物种名、胸径和树高。每个样地均以平均胸径和平均树高为标准,选择3株生长良好、大小一致的林木个体作为标准样株[18](常绿阔叶林以树种比例和样方内树种分布情况为准,标准样株包括米槠、光叶柯和糙皮桦,米槠占主要部分)。采集各样地标准木健康成熟叶片样品300 g(叶按照东南西北四个方向在冠层中分布高度分为上、中、下三层分别取样,然后将样品进行混合)装入信封带回实验室备用。于每个样地中四角的点及对角线的交点选取100 cm×100 cm的凋落物小样方,收集适量凋落叶装入信封(仅收集样方内表层未分解的凋落叶,其中米槠凋落叶占主要部分,其次为光叶柯和糙皮桦),并在对应的样方中用土钻取0—10 cm土壤约500 g。以上采集的样品均带回实验室,植物叶和凋落叶样品经65℃烘干至恒重、粉碎并过100目筛后供元素测定,土壤样品风干并过100目筛后供元素测定。
植物叶和凋落叶样品用H2SO4-H2O2消煮后,分别采用半微量凯氏定氮法(LY/T 1269—1999)测定氮含量和钼锑抗比色法(LY/T 1270—1999)测定磷含量;土壤样品采用半微量凯氏定氮法(LY/T 1228—1999)测定氮含量和钼锑抗比色法(LY/T 1232—1999)测定磷含量;所有样品均采用重铬酸钾-外加热硫酸氧化法(LY/T 1237—1999)测定碳含量。
1.4 数据处理与分析
采用单因素方差分析法(one-way ANOVA)分析相同森林类型不同组分间(植物叶、凋落叶和土壤)、相同组分不同森林类型间N、P再吸收率、C、N、P含量及化学计量比的差异性,各组分C/N、C/P及N/P均以质量比表示,表中数据为平均值±标准误差(n=3),显著性水平为α=0.05;采用Pearson相关分析法分析各森林类型中不同组分间C、N、P化学计量特征的相关性;采用一元线性回归分析法分析该研究区域各组分(植物叶、凋落叶和土壤)中N、P含量相关性[19]。养分(N、P)再吸收率参照曾昭霞等[9]的研究方法。计算公式如下:

数据的计算处理和图表绘制在Microsoft Office Excel 2010中完成、统计分析在SPSS 22.0中完成。
2 结果与分析
2.1 植物叶N、P元素再吸收率
各森林类型间N、P再吸收率存在显著差异(P<0.05)(图1)。5种森林类型中,各林型N再吸收率变化范围为12.71%—61.17%,均值为36.48%,5种林型N再吸收率表现为华山松林>云南松林>常绿阔叶林>高山栎林>滇油杉林;各林型P再吸收率变化范围为44.86%—69.87%,均值为61.20%,5种林型P再吸收率表现为华山松林>云南松林>常绿阔叶林>高山栎林>滇油杉林。各林型P再吸收率均高于N再吸收率。

图1 5种森林类型植物N、P养分再吸收率(平均值±标准误差) Fig.1 The N and P resorption rates of plant in five forest types(Mean±SE)Y:云南松林Pinus yunnanensis forest;H:华山松林Pinus armandii forest;C:常绿阔叶林Evergreen broad-leaf forest;G:高山栎林Quercus semecarpifolia forest;D:滇油杉林Keteleeria evelyniana forest. 不同大写字母表示同一林型N、P再吸收率间差异显著,不同小写字母表示不同林型N(或P)再吸收率间差异显著(P﹤0.05)
2.2 植物叶、凋落叶、土壤C、N、P含量
5种森林类型植物叶、凋落叶和土壤C、N、P含量间存在显著差异(P<0.05)(表2)。5种林型植物叶C、N、P平均含量分别为518.56、18.40、1.52 mg/g;凋落叶C、N、P平均含量分别为491.62、10.43、0.55 mg/g;而土壤C、N、P平均含量分别为9.92、0.76、0.40 mg/g。

表2 5种森林类型植物叶、凋落叶与土壤C、N、P含量(平均值±标准误差)
不同林型间,植物叶、凋落叶和土壤C含量均以常绿阔叶林最高,分别为576.03、517.49、15.07 mg/g。其中植物叶C含量在云南松和华山松林中显著较低(P<0.05),凋落叶C含量除常绿阔叶林外差异不明显(P>0.05),土壤C含量在云南松林和滇油杉林中差异不明显,但显著低于其余三种林型。不同林型N含量在植物叶中表现为云南松林(25.42 mg/g)和华山松林(24.08 mg/g)差异不明显,但显著高于其余三种林型,而在凋落叶和土壤中则表现为常绿阔叶林和高山栎林显著较高(P<0.05)。不同林型P含量在植物叶、凋落叶和土壤中均表现为云南松林最高,其中植物叶P含量在各林型间存在显著差异(P<0.05),而凋落叶和土壤P含量则无明显差异(P>0.05)。
不同组分间,5种林型C、N、P含量除云南松林C含量表现为凋落叶>植物叶>土壤外,其余各林型各元素均为植物叶>凋落叶>土壤。其中,云南松林和华山松林C含量在植物叶和凋落叶间差异均不明显(P>0.05),而其余三种林型C含量在各组分间均差异显著(P<0.05)。此外,5种林型N、P含量在各组分间均存在显著差异(P<0.05)。
2.3 植物叶、凋落叶、土壤C、N、P化学计量比
5种森林类型植物叶、凋落叶和土壤C、N、P化学计量比间存在显著差异(P<0.05)(图2)。结果显示,本研究中5种林型C/N、C/P、N/P均表现为凋落叶>植物叶>土壤。5种林型植物叶、凋落叶和土壤C/N均值分别为31.79、47.26、12.65,C/P均值分别为386.41、919.42、25.80。各林型间植物叶C/N、C/P均表现为云南松林<华山松林<常绿阔叶林<高山栎林<滇油杉林,其中C/N在各林型间存在显著差异(P<0.05),而C/P除滇油杉林较高外,其余林型差异不明显(P>0.05);各林型间凋落叶C/N存在显著差异,C/P则表现为云南松林显著低于其余4种林型;各林型间土壤C/N、C/P均表现为云南松林和滇油杉林显著低于其余三种林型(P<0.05)。
5种林型植物叶N/P在10.17—15.31间,均值为12.27,其中除华山松林较高外,其余林型间无显著差异(P>0.05);凋落叶N/P在13.66—22.14间,均值为19.51,其中除云南松林显著最低外,其余林型间无显著差异(P>0.05);土壤N/P在1.42—2.84间,均值为1.94,其中除华山松林(N/P为1.70)和滇油杉林(N/P为1.64)无明显差异外其余各林型间差异显著(P<0.05)。
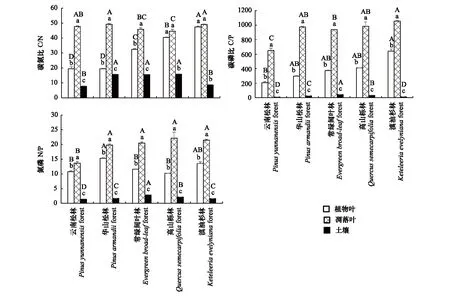
图2 5种森林类型植物叶、凋落叶与土壤化学计量比(平均值±标准误差)Fig.2 The stoichiometric ratio of leaf, litter and soil in five forest types(Mean±SE)不同大写字母表示同一组分不同林型间差异显著,不同小写字母表示同一林型不同组分间差异显著(P<0.05)
2.4 植物叶、凋落叶、土壤C、N、P及其化学计量比相关性
5种森林类型植物叶、凋落叶和土壤各组分间C、N、P含量及其化学计量比Pearson相关分析表明(表3):云南松林植物叶与凋落叶C含量呈极显著正相关(P<0.01),N、P含量均呈显著正相关,凋落叶与土壤N、N/P呈显著正相关(P<0.05);常绿阔叶林植物叶与凋落叶C含量呈极显著正相关(P<0.01),P含量、C/N和C/P呈显著正相关,凋落叶与土壤的C、N含量和N/P呈显著正相关(P<0.05);而华山松林和高山栎林均只有植物叶与凋落叶C含量呈显著正相关(P<0.05);滇油杉林凋落叶与土壤P含量呈极显著正相关(P<0.01)。整体上分析,5种林型植物叶与凋落叶C、N含量均呈极显著正相关(P<0.01),而P含量、C/N和C/P呈显著正相关;凋落叶与土壤的C、N含量和N/P呈显著正相关(P<0.05)。

表3 5种森林类型植物叶、凋落叶与土壤C、N、P含量及化学计量比相关性(Person检验)
5种林型“植物叶-凋落叶-土壤”连续体主要元素N、P间相关关系分析结果表明(图3):研究区5种林型植物叶中N和P含量呈显著正相关关系(R2=0.7688,P<0.01),而土壤中N和P含量呈显著负相关关系(R2= 0.7546,P<0.01),凋落叶中N和P含量间则无明显相关性(R2=0.0169,P>0.05);5种林型N含量在植物叶、凋落叶和土壤中存在较大变异,P含量在植物叶和凋落叶中变异较大,在土壤中变异则不明显。
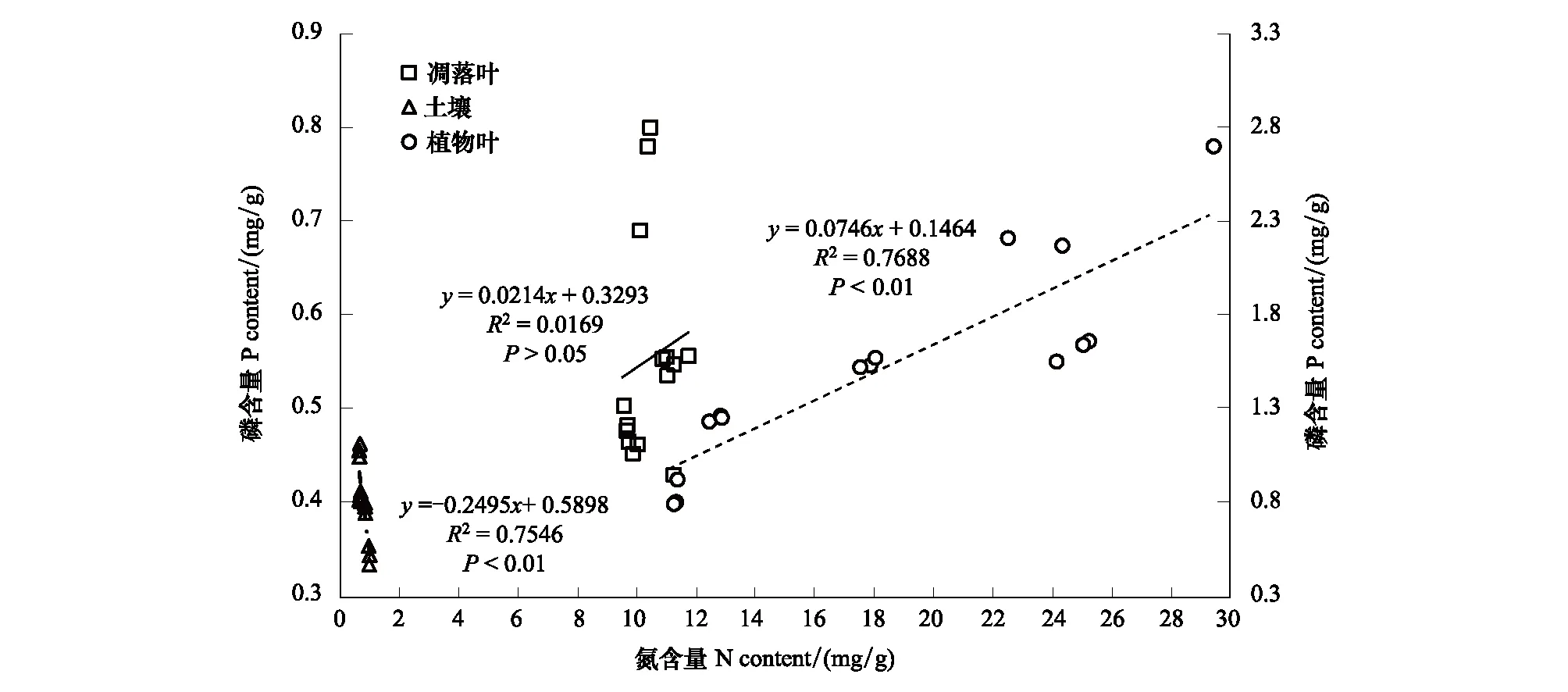
图3 植物叶、凋落叶、土壤N、P含量相关关系Fig.3 The correlation between N and P contents in leaf, litter and soil纵坐标轴中的主坐标轴(左边)表示凋落叶和土壤P含量,次坐标轴(右边)表示植物叶P含量
3 讨论
3.1 植物叶-凋落叶-土壤C、N、P含量及养分再吸收特征
C、N、P作为影响植物生长发育所必需的养分,在植物各种生理生化过程中均发挥着重要作用[2]。本研究中5种滇中亚高山森林类型植物叶C含量平均值为518.56 mg/g,高于全球492种陆生植物叶片C平均含量的464 mg/g[3];同时也高于鄱阳湖湿地优势植物叶C含量的均值(414.3 mg/g)[20]和辽东山区不同森林类型乔木叶C含量的均值(411.08 mg/g)[17],这可能与湿地植物中木质素含量较低及辽东山区林地土壤侵蚀严重有关,一方面,本研究区域多为中龄林,处于快速生长时期,需要大量合成有机物,因此,植物叶片储存C积累有机物的能力相对较强[21],另一方面,该区域除常绿阔叶林外,多为物种单一的林分,易发生病虫害,如云南松林和华山松林可以通过提高单宁、蜡质等富碳次生代谢物的含量增强对病虫害的防御,故植物叶C含量较高[5]。而在该5种林型中尤以常绿阔叶林叶C含量显著较高(576.03 mg/g)。这可能是由于不同植物对环境适应机制不同,该研究区域属于高海拔区(平均海拔约2236 m),常绿阔叶林植物又主要是通过叶片光合作用合成有机物,因此为适应高海拔低温生长环境可能会发生叶片形态的改变,例如增大叶片中栅栏组织的厚度,影响CO2的扩散和固定,进而增加有机物的合成[22]。也有研究认为[23],处于高海拔低温状态下的植物其光合作用中羧化速率较高,从而使得植物C含量较高。此外,也可能是由于不同种植物其元素含量区间不同以及不同种植物生长所依赖的土壤中元素利用有效性不同所导致[9]。研究区5种林型植物叶N含量平均值为18.40 mg/g,低于全球植物叶片的平均水平(20.09 mg/g)[19],而与Han等[14]针对全国753种陆地植物研究所得N含量平均值相近(18.6 mg/g)。本研究5种林型植物叶P含量的平均值为1.52 mg/g,高于全国753种陆地植物叶片的1.21 mg/g[14]、陕西3种主要树种叶片的1.28 mg/g[24]以及桂西北喀斯特6种森林植物叶片的1.2 mg/g[9]。植物叶片营养元素含量与植物自身结构功能特点及生长发育状况有关,不同生长阶段及环境条件的差异会造成较大异质性[25]。淑敏等[11]对不同林龄樟子松(Pinussylvestrisvar.mongolica)人工林的研究发现,樟子松在15年到35年阶段生长速率较快,需要较多的rRNA以增加蛋白质的合成,最终导致针叶N、P含量较高,这与本研究中云南松和华山松两种人工林N、P含量高于其余林型的结果一致,本研究中的云南松林和华山松林正处于生长较快期,需要合成大量干物质,蛋白质等物质的加速合成使rRNA的需求量增大,从而使叶片中的N含量上升,而rRNA又是植物P库的主要贡献者,因此P含量也会相继上升[26-27]。
凋落物作为森林生态系统中生物地球化学循环的一个主要组成部分,其分解过程是植物养分回归土壤的重要途经[28]。本研究5种林型凋落叶C、N、P平均含量分别为491.62、10.43、0.55 mg/g,与辽东山区不同森林类型植物凋落叶相比,本研究C、N含量高于后者的371.62、9.30 mg/g,而P含量低于后者的0.74 mg/g[17];而与桂西北喀斯特地区6种植物凋落叶相比,则表现为C含量高于后者的396.2 mg/g,N含量低于后者的12.7 mg/g[9],与植物叶呈现出一致性,说明了凋落叶秉承了植物叶的特性[8],本研究相关性分析所得植物叶和凋落叶的C、N含量呈极显著相关关系的结果也表明了这一点(表3)。
土壤是植物生长发育的基础和提供所需养分的主要场所,其供应养分的状况与植物光合作用和矿质代谢等生理生化过程密不可分[29]。本研究中5种林型土壤0—10 cm的C、N、P含量平均值分别为9.92、0.76、0.40 mg/g,低于桂西北喀斯特地区相同土层的92.0、6.4、1.5 mg/g[9]和鄱阳湖湿地土壤0—15 cm的25.1、1.9、0.9 mg/g[20],其中C含量更是显著较低;但高于科尔沁沙地东南部樟子松0—20 cm土层的6.89、0.31、0.20 mg/g[11]和黄土高原0—20 cm土层的3.0、0.3、0.3 mg/g[30]。本研究中土壤C含量显著较低可能是由于该区域海拔较高,吕世丽等[31]通过对牛背梁自然保护区森林土壤的研究发现,在海拔达到约1900 m时,土壤水分适宜但温度较低,土壤微生物活动受抑制,有利于有机质积累,但随着海拔的升高,土壤水分和温度均降低,土壤有机质积累减少,而本研究各林型海拔均在2100—2400 m之间,从而C含量相对较低;此外,该区域针叶林相对较多,而针叶林凋落物富含单宁树脂等较难分解的物质,在分解过程中产生残余酸性物质,从而导致土壤酸化,影响微生物活动,导致凋落物的分解受到抑制,进一步降低了土壤C、N输入[32]。在不同林型间,土壤C、N含量存在差异且均以常绿阔叶林最高,而P含量则无明显差异,这可能与P元素主要来源于岩石的风化和淋溶作用有关,并且P形成的矿物质具沉积性,不易迁移[33]。而C、N则主要来源于凋落物的分解,本研究相关性分析所得凋落叶与土壤的C、N含量呈显著正相关的结果也说明了这一点(表3)。在常绿阔叶林中,由于其生物多样性高,地表微生物种类丰富,有利于凋落物的分解,此外,常绿阔叶林植物叶含难分解物质(如单宁、木质素等)较低,从而凋落物分解速率较其他林型高,养分的释放较快,土壤中C、N含量则较高[34]。
植物养分再吸收率反映植物对养分的储存和再利用及适应贫瘠土壤环境的能力[35]。滇中亚高山5种森林类型中,N再吸收率范围为12.71%—61.17%,P再吸收率变化范围为44.86%—69.87%。本研究中云南松林和华山松林N再吸收率(均值为60.09%)显著高于常绿阔叶林和高山栎林(均值为24.76%),这可能是由于云南松和和华山松的土壤N含量显著低于常绿阔叶林和高山栎所致,表明植物会在土壤缺乏某一元素的情况下,自发的增强对该元素的再吸收作用,进而呈现较高的再吸收率[9]。一般情况下,植物叶片在凋落前会将部分N、P元素转移至植物其他器官,从而使得凋落叶N、P含量低于植物叶[36],本研究各林型凋落叶N、P含量显著低于植物叶也验证了这一点。此外,本研究5种林型P再吸收率均值与全球199种木本植物(均值为60.7%)相近[37],而略高于东华172种木本植物的51.0%[38],这种对养分较强的重吸收作用,可以降低植物对土壤养分的依赖性,这在该区域土壤P含量较低的情况下也是一种重要的应对机制[5]。
3.2 植物叶-凋落叶-土壤C、N、P计量比特征
植物C/N、C/P通常用于表征植物吸收N、P并同化C的效率[27],同时也反映了土壤N、P的供应状况,由于土壤养分的供应与植物的光合作用和矿质代谢等生理过程密切相关,所以植物在营养元素供应不足的情况下具有较高的养分利用效率,反之则较低[39]。本研究5种林型植物叶C/N和C/P平均值分别为31.79和386.41,低于赵一聘等[6]研究黄土丘陵沟壑区森林的36.69和438.78;但高于全球平均水平的22.5和232[3]和长白山温带针阔混交林的24.7和321[40]。较高的C/N和C/P反映了植物的N和P利用率高[41],相关研究表明植物在土壤养分缺乏时提高养分利用率是植物适应养分贫瘠状况时的生存策略[42]。而在不同林型间,植物叶C/N、C/P均一致表现为云南松林<华山松林<常绿阔叶林<高山栎林<滇油杉林的规律,这是由于植物在生长过程中合成的核糖体分别由蛋白质和rRNA组成,二者分别为重要的N库和P库,密不可分,相互作用,从而使得植物体中N素和P素保持一致性[43]。本研究5种林型凋落叶C/N和C/P平均值分别为47.26和919.42,低于全球亚热带针叶林凋落物的51.7和1593.3[13],王宁等[44]研究山西省油松(Pinustabuliformis)林认为凋落物的C/N、C/P与其分解速率呈正相关,因此本研究区凋落叶与全球亚热带针叶林相比分解速率较低。在不同林型间,常绿阔叶林和高山栎林的C/N均低于其余三种针叶林。相关研究认为[17],较低C/N的凋落物具有较高的N利用率及较快的凋落物分解速率,也就说明本研究中两种阔叶林凋落物分解较其余林型快,这是由于微生物在凋落物分解初期需要积累足够的N、P养分,较低的C/N能提高微生物活性,进而促进分解[45]。本研究5种林型土壤C/N和C/P分别为12.65和25.80,C/N高于辽东山区的12.60[17]和黄土丘陵区的10.86[7]。研究表明,C/N与土壤有机质的矿化呈负相关[46],因此本研究较以上区域土壤有机质矿化慢。C/P则表征土壤有机质被微生物矿化释放P的潜力,本研究土壤C/P低于辽东山区的53.64[17]而高于黄土丘陵区的10.5[7],说明森林类型、土壤质地等不同会使土壤微生物对有机质的矿化及P的释放存在差异性。此外,本研究5种林型C/N、C/P均表现为凋落叶>植物叶>土壤,这与曾昭霞等[9]研究结果一致,这可能是本研究中N、P再吸收率相对较高所致。植物成熟叶在凋落前,将部分养分转移到其他器官以防止养分供给不足,从而导致凋落叶中养分含量相对较低,这是植物应对极端环境的一种适应机制,本研究中植物均生长在高海拔低温区域,因而对N、P的再吸收更为强烈。
N/P作为用于判断N饱和与P缺乏的关键指标,可以反映环境对植物生长的养分供应状况。Tessier和Raynal[47]对湿地植物的研究指出,当植物的N/P<14时,N是限制植物生长的关键元素,而N/P>16时,P则起主要限制作用;Güsewell[48]研究陆地生态系统并综合比较了大量研究得出,当N/P<10时,植物主要受N的限制,而N/P>20时,则主要受P的限制,介于10—20间则受二者共同限制。不同区域植物种类、生长阶段、生长环境存在差异,这些因素对N/P临界值的变化均会产生影响[49]。本研究对象均为陆地森林群落,因此参照Güsewell的研究结果,5种林型植物叶N/P范围为10.17—15.31,均值为12.27,低于全国森林平均值的16[48],但介于10—20间,表明该区域整体植物的生长可能受到N、P的共同限制。5种林型凋落叶N/P平均值为19.51,大于全球木本植物的12[50],表明该区域可能也存在P匮乏现象。此外,不同组分间,5种林型N/P同C/N、C/P表现出一致规律,均为凋落叶>植物叶>土壤,向云西等[26]对赣南马尾松(Pinusmassoniana)的研究也得出相似结果。若以该滇中亚高山区域森林N/P作为养分供应状况的判断指标,其临界值尚且需要通过进一步的施肥实验进行确定。
植物在生长发育过程中,在生理生化上能够逐渐形成较为完善的自我调节能力,从而适应外界环境的变化,植物C、N、P含量随着环境的变化不断改变,但由于植物生理生化过程的内在联系,C、N、P之间存在明显的相关性[49]。本研究结果显示,滇中亚高山区域植物叶N、P含量呈极显著正相关(P<0.01),凋落叶N、P含量无明显相关性,而土壤N、P含量则呈极显著负相关(P<0.01)(图3),与曾昭霞等[9]研究结果一致,说明养分元素在植物中具有明显的耦合性,同时体现了植物养分元素间的共变性和传承性[24],这也映照上文所讨论“植物叶C/N、C/P均一致表现为云南松林<华山松林<常绿阔叶林<高山栎林<滇油杉林”的结论,植物在生长过程中蛋白质和rRNA保持着一致性,二者又分别为重要的N库和P库,密不可分,相互作用,从而使得植物体中N素和P素也保持一致性。至于土壤中N、P含量的负相关关系,其中一个原因可能是土壤N、P元素来源不同导致,此外,外界因素如水热条件、人为干扰等也是重要影响因素,具体有待后续深入研究进行探明。
本研究主要针对滇中亚高山5种森林类型植物叶、凋落叶和土壤养分化学计量特征及其相关性进行了探讨,对于揭示该区域森林生态系统养分状况和养分循环特征具有重要意义,并为后续研究该区域森林生态系统养分限制情况提供理论数据。但本研究未考虑环境因子等对化学计量特征的影响,因此进一步结合海拔、温度、降雨等环境因子等对不同森林类型“植物叶-凋落叶-土壤”连续体化学计量特征进行研究具有重要意义,此外,若以该滇中亚高山区域森林N/P作为养分供应状况的判断指标,开展该区域不同梯度施肥实验对确定其临界值具有必要性。
4 结论
(1)5种森林类型植物N、P再吸收率均为华山松林>云南松林>常绿阔叶林>高山栎林>滇油杉林;各林型P再吸收率均高于N再吸收率。
(2)5种森林类型C、N、P含量除云南松C含量为凋落叶>植物叶>土壤外,其余均表现为植物叶>凋落叶>土壤。
(3)5种森林类型C/N、C/P、N/P均表现为凋落物>植物叶>土壤;各林型间植物叶C/N、C/P均表现为云南松林<华山松林<常绿阔叶林<高山栎林<滇油杉林;各林型生长均受到N、P元素的共同限制。
(4)5种森林类型植物叶与凋落叶C、N含量均呈极显著正相关,而P含量、C/N和C/P呈显著正相关;凋落叶与土壤的C、N含量和N/P呈显著正相关;5种林型植物叶中的N和P含量呈显著正相关关系;土壤中N和P含量呈显著负相关关系。
