青海云杉育种资源遗传评价
2020-11-17陈雅丽赵祜吕东李伟张宏斌
陈雅丽,赵祜,吕东,李伟,张宏斌
(1.北京林业大学生物科学与技术学院,北京 100083;2.甘肃省祁连山水源涵养林研究院,甘肃 张掖 734000;3.祁连山特有植物繁育及推广国家地方联合工程研究中心,甘肃 张掖 734000;4.甘肃省祁连山特有珍稀树种保护繁育工程研究中心,甘肃 张掖 734000)
青海云杉(PiceacrassifoliaKom.)为松科云杉属乔木,是中国特有树种,分布于我国青海、甘肃、宁夏、内蒙古等省(区),其适应性强,具有耐低温、耐旱、耐瘠薄的特性,是高山区森林更新和荒山造林的重要树种[1-2].青海云杉木材材质优良,可作建筑、桥梁、家具及木纤维工业原料等用材,也是重要的用材树种[3-4].国内开展青海云杉的遗传改良工作是从20世纪70年代开始,相继在各分布区内建立了母树林及种子园[5],目前对于青海云杉种子园育种资源的研究多集中在无性系的开花散粉规律[6-8]、木材和种实等表型性状的变异[9-11]和半同胞子代测定[12-14]等的研究上,基于分子水平分析种子园遗传多样性的研究较少.利用DNA分子标记技术研究物种的遗传多样性可以直接从分子层面反映遗传多态性,除却了环境因素的干扰,而且DNA分子标记可以提供大量的遗传变异信息,是群体遗传关系研究的有力工具[15].胡杨[16]利用ISSR分子标记研究了青海云杉无性系种子园的遗传多样性,结果表明种子园的遗传多样性水平较高,袁行栋[17]利用SSR分子标记对青海云杉无性系的遗传多样性的研究也得出相同结论,但二者对于无性系间的亲缘关系的分析不全面,没有反映不同种源无性系的亲缘关系.育种资源亲缘关系的明确至关重要,对开展杂交实验、种子园无性系配置等具有重要指导意义,能最大限度地避免近交衰退,保证高世代种子园的品质稳固提高,是种子园可持续多世代改良的前提[18].
本研究以张掖龙渠青海云杉无性系种子园为研究对象,采用SSR分子标记技术,对青海云杉无性系种子园的育种资源进行了遗传多样性及个体间、种源间的亲缘关系等研究,以期为青海云杉遗传改良策略的制定及高世代种子园的建设奠定理论基础.
1 材料与方法
1.1 试验材料
甘肃省张掖市龙渠青海云杉无性系种子园位于祁连山脚黑河出山口东侧的冲击扇上,N 38°48′41″,E 100°13′42″,海拔1 700 m,年降水量为193.0 mm,年蒸发量为1 653.0 mm,年均气温7.4 ℃,最高气温33.4 ℃,最低气温-26.5 ℃,相对湿度51%,无霜期152 d,日照时间2 435.6 h.1985年建园,建园材料来源于祁连山区的西水、连城、哈溪、古城、东大山、大黄山、大河口、隆畅河、祁连等种源地,种子园采用完全随机排列设计,品字型栽植,株行距5.0 m×5.0 m.
本研究对种子园中108个无性系进行了分析,各种源无性系数量见表1,采集每个无性系单株当年生健康针叶,与硅胶干燥保存于塑料袋中,分别编号,带回实验室,置于-80 ℃冰箱内保存.
1.2 DNA的提取
用天根植物基因组DNA提取试剂盒(北京天根生化科技有限公司)提取青海云杉无性系基因组DNA,用0.8%的琼脂糖凝胶电泳初步检测 DNA质量,用微量分光光度计(Nano Drop ND-1000)检测DNA的浓度、纯度,置于-20 ℃冰箱中保存备用.

表1 试验材料信息
1.3 引物筛选及PCR扩增
从60对来自挪威云杉(PiceaabiesKarst.)的SSR引物[19]中筛选出10对扩增条带清晰、稳定性好、具有多态性的SSR引物用于本研究,各引物详细信息见表2.引物由Sangon公司合成,20 μL SSR-PCR扩增反应体系包括:模板DNA(5~10 ng/μL)1.0 μL、d NTP(10 mmol/L)0.45 μL、引物(10 μmol/L)0.25 μL、Mg2+(25 mmol/L)2 μL、TaqDNA聚合酶(5 U/μL)0.8 μL、10×PCR缓冲液2 μL、HPLC超纯水13.25 μL.PCR扩增程序为:94 ℃预变性3 min;94 ℃变性30 s,50~60 ℃退火30 s,72 ℃延伸1 min,44个循环;72 ℃延伸3 min;4 ℃保存.扩增产物进行电泳,在紫外灯下拍照记录.

表2 10对SSR引物信息
1.4 电泳结果统计与分析
参考电泳图谱中的分子量标准,根据 PCR 产物条带大小从小到大依次用大写英文字母A、B、C等表示,确定每个个体的基因型.用POPGENE 32软件计算观测等位基因(Na)、有效等位基因(Ne)、Shannon信息指数(I)、Nei多样性指数(H)、观测杂合度(Ho)、期望杂合度(He)、近交指数(F)、遗传分化系数(Fst)、基因流(Nm)等遗传参数.通过GenAlEx 6.5软件进行分子方差(AMOVA)分析.使用Powermarker V3.25计算基于等位基因频率的Nei’ 1983遗传距离,采用非加权组平均法(UPGMA)对Nei’ 1983遗传距离进行聚类,用MEGV7.0.14查看聚类结果,根据聚类结果分析亲缘关系情况.
2 结果与分析
2.1 青海云杉基因组DNA提取结果
使用天根植物基因组DNA提取试剂盒提取青海云杉无性系的基因组DNA,用0.8%琼脂糖凝胶电泳后,结果如图1所示,可以看出条带的亮度较好,比较集中,用微量分光光度计(Nano Drop ND-1000)检测DNA的浓度为75 ng/μL左右,纯度D值(260/280)约为1.8,符合进行SSR-PCR反应的要求.

图1 青海云杉无性系基因组DNA电泳结果Figure 1 Genomic DNA electrophoresis results of P.crassifolia clones
2.2 SSR扩增结果及SSR位点多态性
利用10对SSR引物对108个青海云杉无性系进行SSR-PCR扩增,经电泳检测后,结果如图2所示,条带清晰,多态性较好,对扩增结果进行统计分析,结果如表3所示,共扩增出50个等位基因,10个位点检测到等位基因(Na)3~8个,平均为5个;有效等位基因(Ne)为1.212 8~5.843 3,平均为2.484 2;观测杂合度(Ho)为0~0.500 0,平均为0.156 6;期望杂合度(He)为0.176 3~0.832 9,平均为0.515 4;Nei多样性指数(H)为0.175 5~0.828 9,平均为0.512 9;Shannon信息指数(I)为0.347 8~1.876 0,平均为1.006 5.其中,EAC1F04位点的多态性水平最高,各项遗传参数值均较高,对所研究的青海云杉无性系遗传多样性贡献较大,相应的,EAC6D01位点的多态性水平则最低.
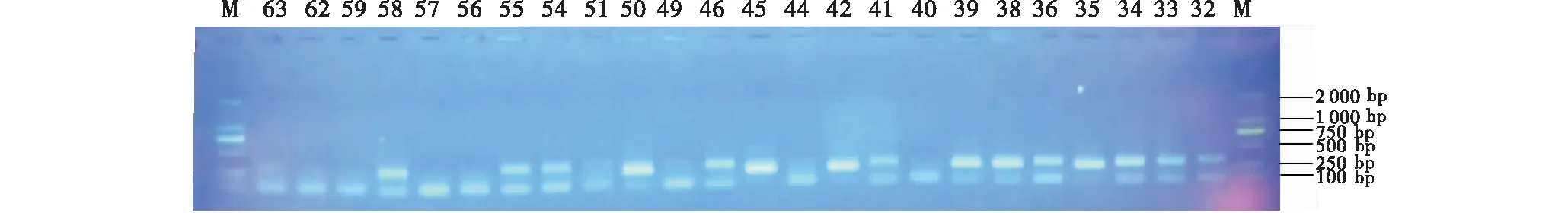
图2 引物EAC1C08对部分青海云杉无性系的扩增图谱Figure 2 The amplification pattern of part of P.crassifolia clones by primer EAC1C08

表3 基于SSR标记的青海云杉无性系遗传多样性参数
2.3 不同种源遗传多样性差异
9个青海云杉种源的遗传多样性参数汇总如表4所示,观测等位基因(Na)为2.0~3.4,西水(XS)种源的值最高,祁连(QL)种源的值最低,平均为2.6,平均有效等位基因(Ne)为1.902 3.观测杂合度(Ho)为0.100 0~0.280 0,祁连(QL)种源的值最高,哈溪(HX)种源的值最低,平均为0.170 9;期望杂合度(He)为0.300 7~0.488 9,大河口(DHK)种源的值最高,哈溪(HX)种源的值最低,平均为0.400 3.Shannon 信息指数(I)的变化范围为0.467 2~0.819 6,平均为0.656 1,Nei多样性指数(H)的变化范围为0.278 0~0.476 5,平均为0.376 1,两者皆为古城(GC)种源的值最大,祁连(QL)种源的值最小,种源间数值相差较大,说明种源间遗传多样性水平差异较大.采用Shannon信息指数(I)、Nei多样性指数(N)、期望杂合度(He)这几个参数来度量9个青海云杉种源的遗传多样性水平,由大到小依次为古城(GC)、东大山(DDS)、西水(XS)、大河口(DHK)、连城(LC)、大黄山(DHS)、隆畅河(LCH)、哈溪(HX)、祁连(QL)种源.

表4 9个青海云杉种源的遗传多样性分析
2.4 不同种源间的遗传分化
青海云杉9个种源的遗传分化系数(Fst)和基因流(Nm)(表5)为0.281 6和0.637 8,表明种源间存在较高程度的遗传分化,种源之间的基因交流不频繁,容易发生遗传漂变现象[20].
分子方差(AMOVA)分析结果(表6)表明84%的遗传变异存在于个体间,只有16%来源于种源间.青海云杉个体间变异大于种源间,说明青海云杉个体间变异为主要变异.

表5 遗传分化系数与基因流
由表5所示,10个SSR位点的近交系数(F)在0.013 7~1.000 0之间,平均值为0.681 0,表明种子园内无性系纯合子过量,杂合子较少.

表6 9个青海云杉种源的AMOVA分析
2.5 青海云杉育种资源亲缘关系分析
2.5.1 不同种源间的亲缘关系分析 计算9个青海云杉种源的遗传距离,用UPGMA算法进行聚类,得到种源间的聚类图(图1).根据聚类图,可以看出9个种源大致分为2组,第1组为大黄山(DHS)、古城(GC)和哈溪(HX)种源,第2组为东大山(DDS)、西水(XS)、大河口(DHK)、连城(GC)、祁连(QL)和隆畅河(LCH)种源.由图可知,大黄山(DHS)、古城(GC)和哈溪(HX)种源之间存在一定的亲缘关系,古城(GC)和哈溪(HX)种源亲缘关系更近;东大山(DDS)和西水(XS)种源亲缘关系比较接近,它们和大河口(DHK)种源存在一定的亲缘关系;连城(LC)和祁连(QL)种源的亲缘关系比较接近;第2组中,隆畅河(LCH)种源与其他种源的亲缘关系较远.
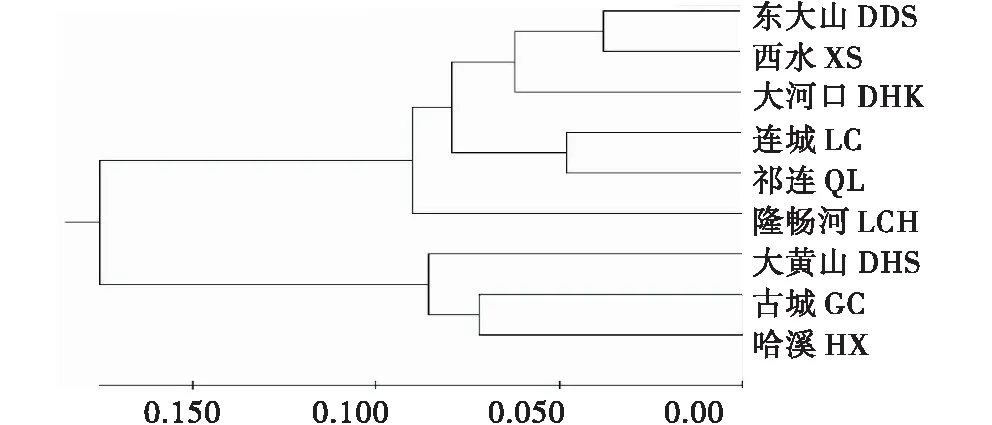
图3 青海云杉9个种源间亲缘关系UPGMA聚类图Figure 3 UPGMA clustering of relationship among the 9 provenances of P.crassifolia
2.5.2 个体间的亲缘关系分析 基于108个青海云杉无性系间的Nei’ 1983遗传距离进行UPGMA聚类分析,得到108个个体的亲缘关系图谱(图4).108个个体大致分为两大组,基本与种源的聚类结果相一致:第1组中有29个无性系,大黄山(DHS)、古城(GC)和哈溪(HX)种源的无性系占75.9%;第2组中有79个无性系,东大山(DDS)、西水(XS)、大河口(DHK)、连城(GC)、祁连(QL)和隆畅河(LCH)种源的无性系占97.5%.聚类结果显示无性系间存在逐步分化的关系,种源相同的无性系优先聚在一起,种源聚类图中某些种源亲缘关系较近,在个体聚类图中这些种源的无性系也大多聚为一类.

图4 108个个体亲缘关系UPGMA聚类图Figure 4 UPGMA clustering of 108 individuals
3 讨论与结论
青海云杉是个古老的树种,拥有广泛的分布范围,已有研究表明,青海云杉天然群体具有较高的遗传多样性水平[21-22].本研究采用SSR分子标记技术对张掖龙渠青海云杉无性系种子园进行了遗传多样性研究,结果共扩增出了50个多态位点,平均等位基因数(Na)为5,平均有效等位基因数(Ne)为2.484 2,平均观测杂合度(Ho)为0.156 6,平均期望杂合度(He)为0.515 4,平均Nei多样性指数(H)为0.512 9,平均Shannon信息指数(I)为1.006 5,表明青海云杉育种资源具有较高的遗传多样性水平,遗传结构优良,可为高世代种子园的建设提供丰富的原材料.与其他基于SSR标记的针叶树遗传多样性的研究结果相比,青海云杉的遗传多样性低于同为云杉属的粗枝云杉(PiceaasperataMast.)(Nei多样性指数为0.707)[23]、天山云杉(Piceaschrenkiana),Shannon信息指数为1.281 1[24],高于华北落叶松(Larixprincipis-rupprechtiiMayr)(Shannon信息指数为0.712 7)[25]、白皮松(PinusbungeanaZucc.)(Shannon信息指数为0.401 0)[26]、油松(Pinustabuliformis)(Shannon信息指数为0.599)[27],稍低于长白落叶松(LarixolgensisHenry)(Shannon信息指数为1.020 3)[28]、樟子松(Pinussylvestrisvar.mongolica),Shannon信息指数为1.034[29].
结合Shannon信息指数(I)、Nei多样性指数(H)、期望杂合度(He)这几个遗传参数的值,古城(GC)、东大山(DDS)、西水(XS)种源的遗传多样性水平相对较高,隆畅河(LCH)、哈溪(HX)、祁连(QL)种源遗传多样性水平相对较低,但隆畅河(LCH)和祁连(QL)种源地的样本数量较少(n=5),容易存在“遗传漂变”的影响,这2个种源地的遗传多样性水平还有待于进一步研究.
青海云杉种源间存在较高程度的遗传分化,遗传分化系数(Fst)为0.281 6,基因流(Nm)为0.637 8,说明群体之间基因交流较少[30],这也是导致种源之间遗传分化水平较高的原因.孙雪新等[31]和李萍等[21]利用同工酶标记分子研究结果表明青海云杉天然群体间的遗传分化水平低,群体平均分化系数分别为0.023和0.064 3.王文峰[22]利用ISSR分子标记技术对青海云杉天然群体遗传多样性研究的结果为群体间的遗传分化系数(Gst)为0.289 7、群体间基因流(Nm)为0.613 0,与本研究的结果相似.由此说明了利用不同的标记研究青海云杉群体遗传分化水平的结果有差异,而DNA分子标记可以直接反应植物在分子水平上的遗传变异,因此得出的结果也更准确.分子方差(AMOVA)分析结果显示大部分的遗传变异存在于个体间(84%),王娅丽等[32-33]研究了青海云杉天然群体表型性状和种实性状的变异,结果也发现了群体内变异占主要地位(68.80%、72.82%).青海云杉无性系种子园平均近交系数值为0.681 0,纯合子严重过量,说明种源内部近交程度严重.
群体聚类分析发现,9个种源可分为2组,第1组为大黄山(DHS)、古城(GC)和哈溪(HX)种源,第2组为东大山(DDS)、西水(XS)、大河口(DHK)、连城(LC)、祁连(QL)和隆畅河(LCH)种源,其中有3对种源的亲缘关系较近,他们分别是古城(GC)种源和哈溪(HX)种源、东大山(DDS)种源和西水(XS)种源、连城(LC)种源和祁连(QL)种源.据资料显示,哈溪(HX)种源与古城(GC)种源在地理位置上相近,东大山(DDS)、西水(XS)和大河口(DHK)种源之间的地理位置相近,由此说明种源间亲缘关系与地理距离存在相关性,与孙雪新等[31]、王娅丽等[33]、王文峰[22]的研究结果一致.个体间的聚类基本上也分为两大组,与种源间聚类结果相对应,种源相同的无性系多聚为一类.
