麦吊云杉林分密度对林下物种多样性及其自然更新能力的影响
2020-11-04刘世杰彭建莫劲雁李晓清辜云杰
刘世杰, 彭建, 莫劲雁, 李晓清, 辜云杰
1. 四川省林业科学研究院,森林和湿地生态恢复与保育四川重点实验室,四川 成都 610081;
2. 长江上游林业生态工程四川省重点实验室,长江上游森林资源保育与生态安全国家林业和草原局重点实验室,华西雨屏区人工林生态系统研究长期科研基地,四川农业大学,生态林业研究所,四川 成都 611130;
3. 四川勿角自然保护区管理处,四川 九寨沟 623400
林分密度是指单位面积林地上的立木个体数量,是森林群落结构的基础指标,也是森林培育过程中能够认为控制的主要因子[1]。林分密度在一定程度上决定了林内个体间的空间生态位,是影响林下植被物种组成和多样性的重要限制因子[2,3]。许多研究表明,随着林分密度的增大,林下灌木和草本植物的丰富度指数、多样性指数表现出减小趋势[4]。高密度桉树林下灌木层和草本层物种多样性明显低于低密度桉树林[5]。杉木人工林中,当林分密度过高时,对林地伤害巨大,基于植物多样性的主成分分析结果出现“驼峰模式”,不利于植物多样性的发展[6]。林分密度会影响林内环境以及地被物,是林木天然更新的主要障碍因子[7]。张树梓等[8]研究塞罕坝华北落叶松人工林天然更新影响子时发现,高密度的林分都表现出高个体数,低成活率的现象,是造成林分天然更新困难的主要因素。而张希彪等[9]在研究油松人工林群落更新特征时发现,当林分密度从650株·hm−2增加到1 550株·hm−2时,林下更新的幼苗、幼树密度呈增加趋势。可见,不同树种天然更新与林分密度的关系还有很大的不确定性,需要通过更多的研究以揭示其内在规律。
麦吊云杉(Picea brachytyla),又名麦吊杉、垂枝云杉,系松科云杉属鱼鳞云杉组植物,我国的三级重点保护树种,其原始分布中心为四川岷江和青衣江流域的山区,现仅局限于我国西部山地的秦岭,大巴山、米仓山、岷山和贡嘎山等地区[10]。麦吊云杉材质致密,韧性大,可供建筑、飞机、机器、家具等用材,具有重要的经济价值和科学价值,同时,它垂直分布在海拔1 800 m~3 000 m之间,最低下限1 300 m(汶川),最高可达3 200 m(岷江、青衣江山区),对水源涵养、保持水土有着不可估量的作用,是选作分布区内森林更新及高山造林的极好树种[11]。目前,我国已有对该树种的地理分布、群落结构、分类特征、林学特征以及引种驯化栽培、抚育管理等方面的研究[11]。多年来由于人为破坏,采伐和保育方式的不合理,使得麦吊云杉天然林群落处于衰退状态。因此,研究麦吊云杉天然林林分密度对林下植被多样性和天然更新能力的影响,有助于揭示林分密度对林下植被多样性的影响和调节规律,评价麦吊云杉天然林是否具有可持续的自然更新能力,可为退化麦吊云杉天然林的恢复和保育提供依据。
1 研究区概况与研究方法
1.1 研究区域概况
研究地区位于四川省阿坝藏族羌族自治州九寨沟县大录乡(103°40′36″E,33°25′58″N)、贡杠岭(103°43′56″E,33°3′22″N)和甲勿池(104°10′4″E,32°59′4″N),海拔高度在2 000~3 000 m之间。研究区域属高原湿润气候,冬长夏短,年均气温12.7 ℃,年均降水量550 mm,昼夜温差大,冬季较寒冷,气温多在0 ℃以下。研究区域属于四川省贡杠岭自然保护区和四川省勿角自然保护区,区内植被保存较为完好。
1.2 标准样地设置和野外调查
根据前期样线调查基础上,选择九寨沟县3个麦吊云杉天然集中分布区内设置标准样地。共设面积 20 m×20 m的标准地30个,其中大录乡10个标准地、贡杠岭10个、甲勿池10个,在每个标准样地的对角线1/4处分别设5 m×5 m的林下植被调查重复样方4个,共计120个小样方。标准地概况见表1。
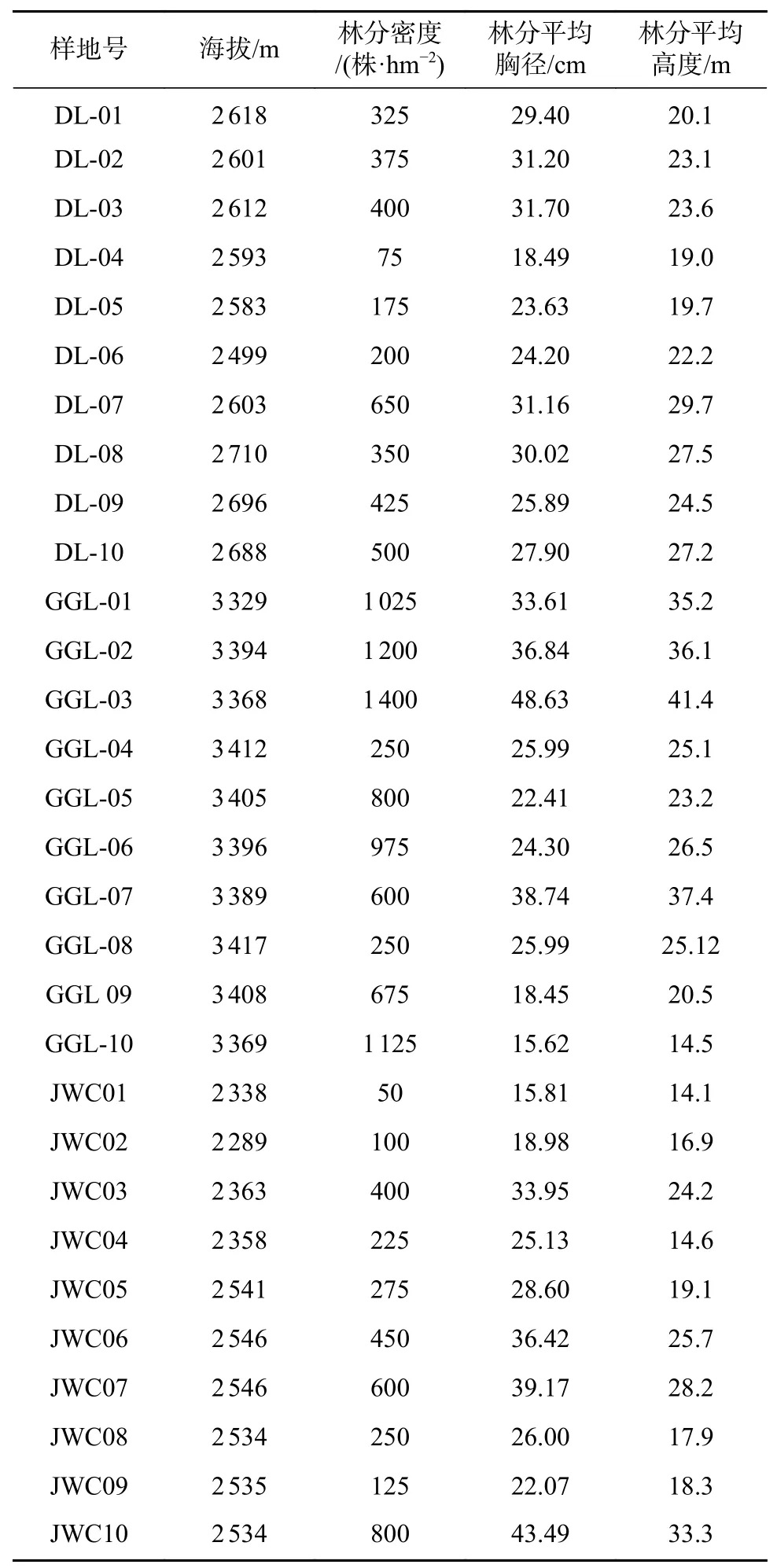
表1 标准地概况Tab.1General information of sampled plots
在设好的标准样地内进行标准地调查,记录标准地的经纬度、海拔等立地因子以及林分密度。在每个样方内进行林下植被群落学调查,记录样方内出现的所有灌木和草本植物种名和数量。同时,记录每个样地内更新的幼苗数量(DBH<4 cm)。调查在 2019 年 7—10 月进行。
1.3 数据分析和指标计算
根据样方内调查统计的各物种的株数计算其密度,参照丁继伟等[1]的方法稍加改动,计算物种重要值,具体公式如下:
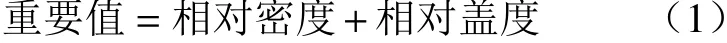
其中,相对密度=(某种的密度/所有种的密度之和)×100%,相对频度=(某种的频度/所有种的频度之和)×100%,相对盖度=(某种的盖度/所有种的盖度之和)×100%。
物种多样性指数相关指标的计算参照以下公式进行:
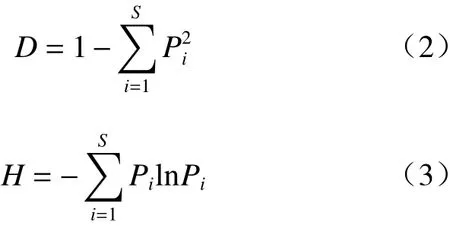
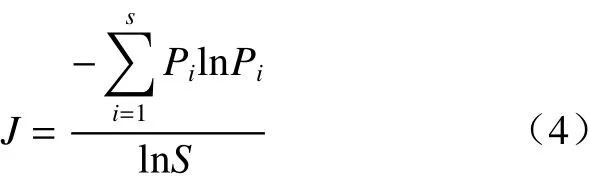
式中,S是群落中的物种总数,Pi是某物种的个体数占群落中所有物种总个数的比值,D是Simpson指数,H是Shannon-Weiner指数,J是Pielou均匀度指数。
2 结果与分析
2.1 不同密度林分伴生植物及林下植被物种组成分析
不同密度林分下植物及伴生植物的物种组成及其重要值见表2。麦吊云杉天然林从45~1 400株·hm−2不同密度林分下植被共出现物种26个,分属15科22属;其中,伴生植物9种,分属5科9属,林下植被17种,分属12科13属。从图1可以看出,林下植被和伴生植物的科、属、种数量大致随着林分密度呈现先升高后下降的趋势。冷杉、糙皮桦等是各密度林分中较为常见的伴生乔木植物。在不同密度林分中,蹄盖蕨、青绿薹草和狗舌草构成了草本植被本底;野草莓、荚蒾类、三颗针等耐阴性很强的矮小灌木是各密度林分中灌木植物的代表种,高山柳、高山杜鹃等较高的灌木也是林下常见的灌木种。
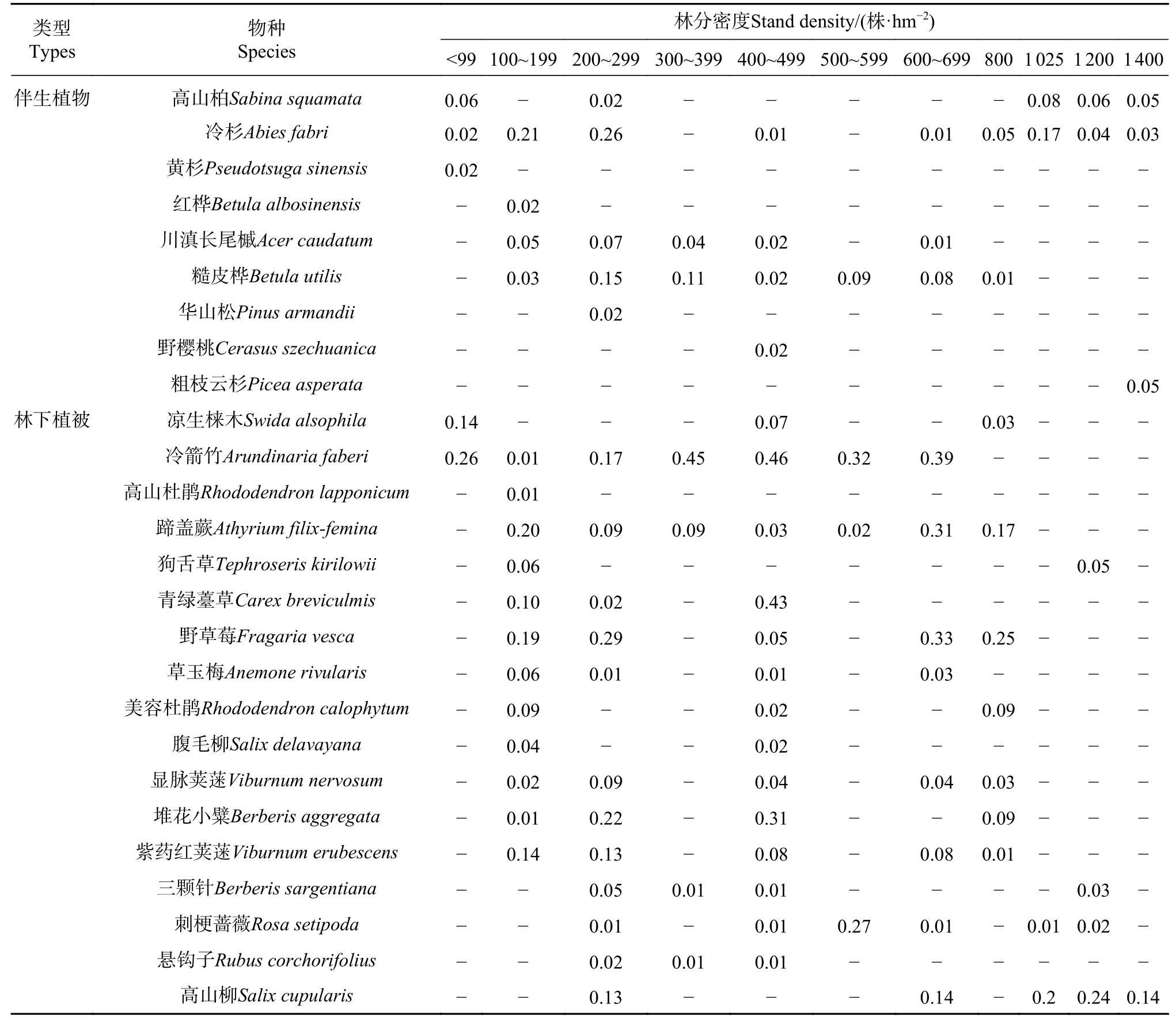
表2 不同密度麦吊云杉伴生植物及林下植被重要值Tab.2Species and importance values of associated plants and understory vegetation in Picea brachytyla natural forest with different stand densities
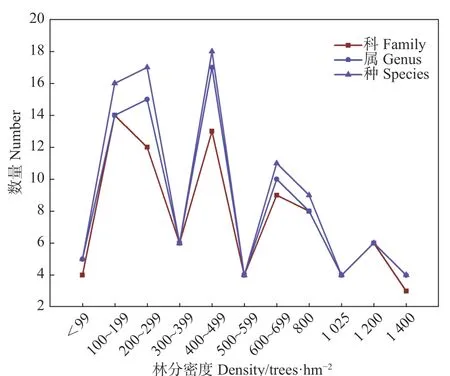
图1 不同林分密度下植物科、属、种数量Fig.1Number of families, genera, and species under different stand densities
2.2 林下植被丰富度指数随林分密度的变化
麦吊云杉天然林不同林分密度的物种丰富度指数随着林分密度的增大表现为双峰响应模式,但整体呈先升高后降低的趋势,在林分密度为200~299 株·hm−2和400~499株·hm−2时林下植被的物种丰富度指数达到峰值(见图2)。林下乔木、灌木和草本植物物种数在不同林分密度之间的差异较大,乔木种数的变化在1~5,灌木植物种数变化在1~9,而草本植物种数变化在0~5。其中,乔木种在林分密度400~499株·hm−2出现峰值,灌木植物种在林分密度度400~499株·hm−2出现峰值,草本植物种在100~199株·hm−2、200~299株·hm−2和400~499株·hm−2等3个林分密度下出现峰值。
2.3 不同密度林分植被多样性的变化
从表3可以看出,Simpson指数和Shannon-Weiner指数的表征作用相近,以Shannon-Weiner指数的区分度更好。林分密度对伴生植物和林下植被物种多样性有显著影响,在0~499株·hm−2范围内,林分植被多样性随林分密度增大而增加,在500~1 400株·hm−2范围内,植被物种多样性指数虽林分密度增大而明显下降,与物种丰富度指数的变化趋势一致,呈双峰模式。林下植被Pielou均匀度指数随着林分密度的增大,大致呈现出逐渐增大的趋势。可见,林分密度对植被个体空间分布格局有一定影响。
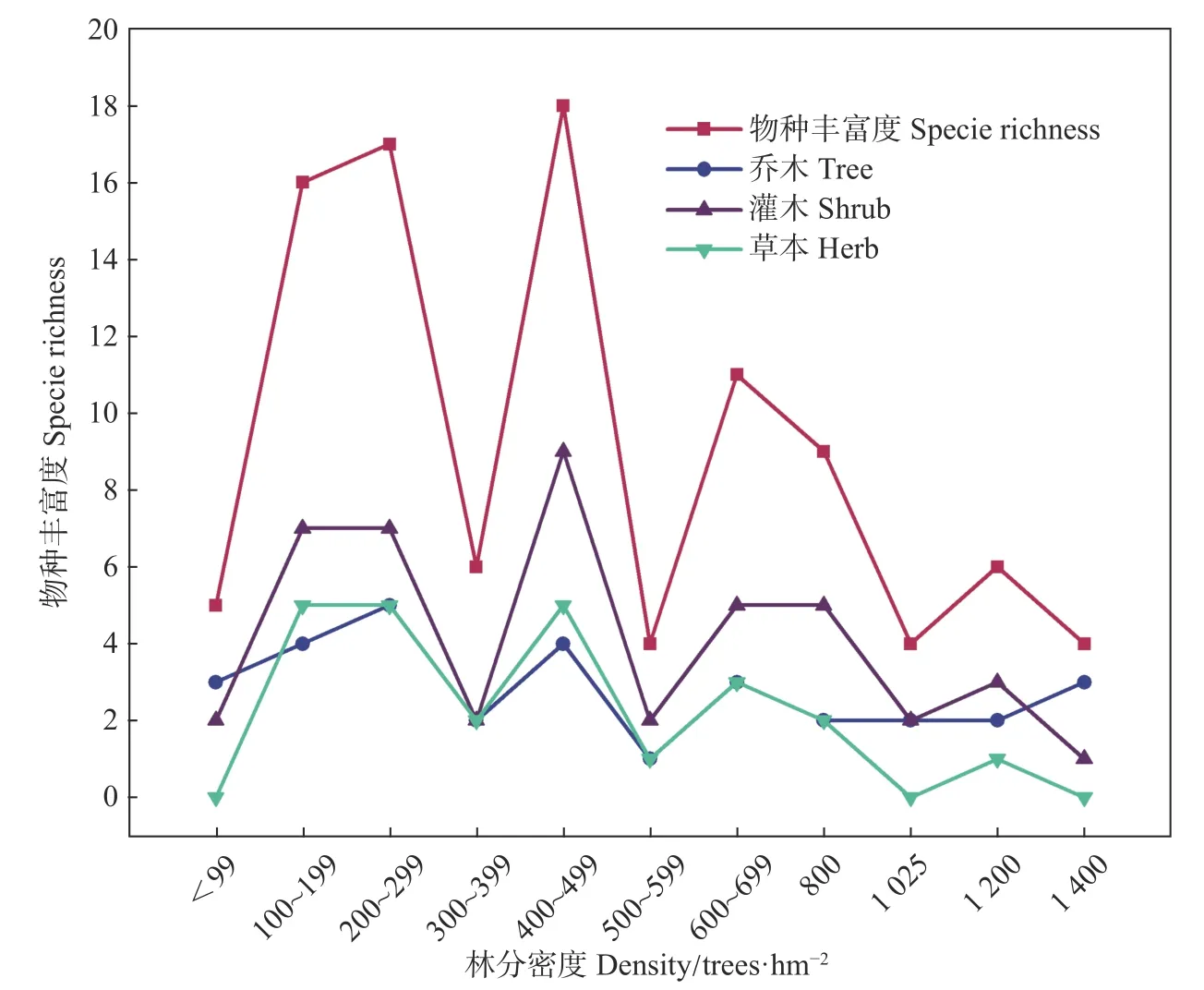
图2 不同密度林分下物种丰富度、乔木、灌木、草本数变化Fig.2Variation of species richness, tree, shrub, and herb under different stand densities
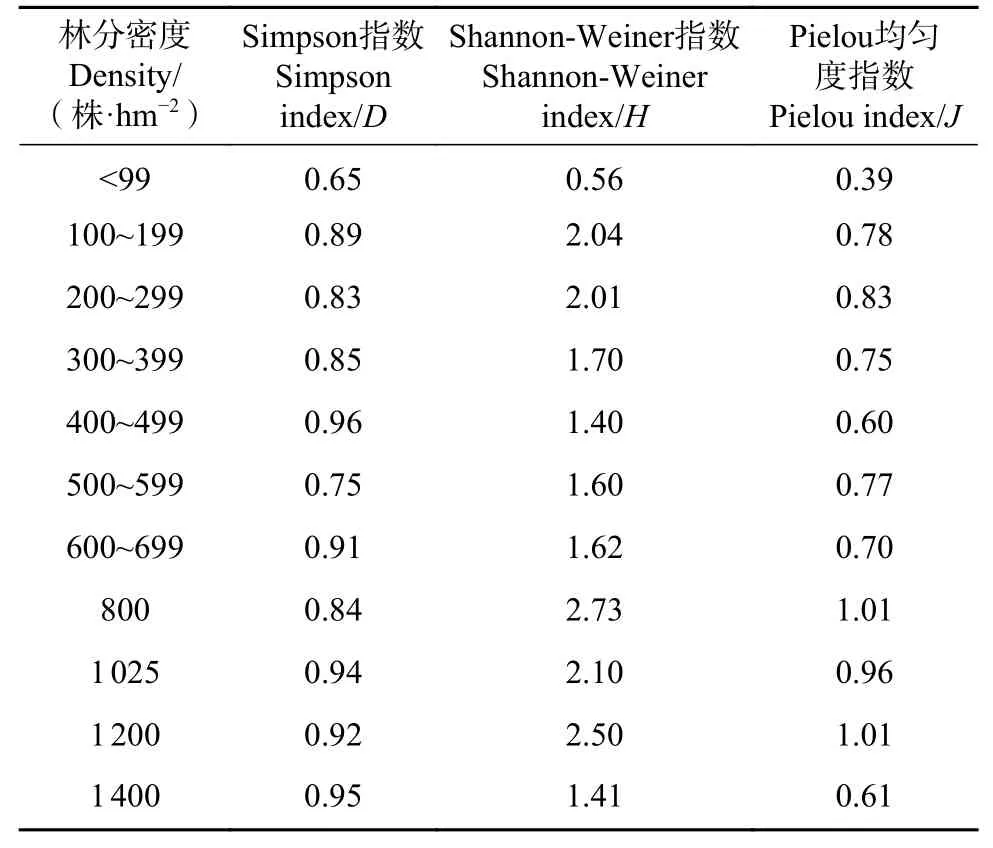
表3 不同林分密度下物种多样性指数Tab.3Species diversity indexes of understory vegetation under different stand densities
2.4 林分密度对自然更新能力的影响
由 图3可 知,仅200~299株·hm−2、600~699株·hm−2、1 200株·hm−2和1 400株·hm−2林分密度下观测到更新幼苗(见图3)。其中,1 400株·hm−2密度林分下更新幼苗数量最多,为50株幼苗。调查中发现 1 400 株·hm−2林分样方中存在大的林窗,该林窗下灌木和草本较少,且盖度较低,麦吊云杉更新幼苗全部集中在林窗下;而其他如 600~699 株·hm−2、100~199 株·hm−2等更新幼苗极少或无更新幼苗的林分林窗较少,林下灌木和草本丰富,且盖度较大。这可能是造成 1 400 株·hm−2林分密度较大,但更新幼苗较多这种极端差异的原因。
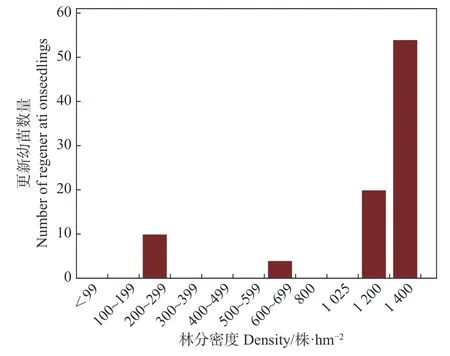
图3 不同密度林分更新幼苗数量Fig.3Number of regeneration seedlings under different stand densities
3 讨论
林分密度通过改变林分郁闭度等林内环境因子影响林下生境,进而对植被物种数量和多样性造成影响[12]。研究地区的麦吊云杉天然林在45~1 400株·hm–2密度范围伴生植物和林下植被共出现物种26种,分属15科22属,其中乔木9种,主要为糙皮桦。王建宏等[13]对白水江自然保护区麦吊云杉群落类型调查研究发现,麦吊云杉群落中乔木有14种,主要为糙皮桦和红桦。与本研究结果具有相似性,但物种数量和种类存在一定的差异,而这种差异的形成可能与地理环境、林分密度大小和研究地区纬度高低有关。丁继伟等[1]认为林下植被多样性对林分密度持续增大的响应表现为非单一变化趋势的规律,随着林分密度的增大,林下植被多样性指数呈现先增大后减小的变化规律。麦吊云杉天然林群体林下灌木和草本物种多样性指数在不同林分密度之间的差异较大,并出现双峰响应模式。其结果表明麦吊云杉天然林分在一定的密度(500株·hm–2)之下,对其林下植被多样性影响小。邹扬等[14]研究发现,高寒区青海云杉人工林林下灌木数量受林分密度的影响要高于草本植物。李伟伟等[15]认为灌木与草本植物对林分密度变化引起林内生境改变的敏感度不同,灌木受林分密度变化的影响较草本少。而本研究结果显示,麦吊云杉天然林林下灌木和草本随林分密度变化的趋势保持一致,其原因可能为麦吊云杉的林下灌木和草本较少,且对生境的要求均较高,且基本保持一致,因而对林分密度变化引起的生境改变相对一致。物种均匀度不受物种丰富度影响,只与物种个体树木在群落或环境中分布的均匀程度有关,是对不同物种在个体数上接近程度的衡量[14]。研究还发现,麦吊云杉天然林林下植被均匀度指数随林分密度的增大呈现增加的趋势,但不规律,说明林分密度对林下植被的分布状况有影响,但不明显。吕婧娴等[16]研究发现,油松人工林草本层的均匀度指数与林分密度呈显著负相关关系。而周树平等[17]对不同密度柚木人工林林下植被的研究结果显示,随着林分密度增加,柚木人工林林下植被均匀度呈先增加后减少的趋势。可见,林分密度对林下植被多样性的影响具有一定的复杂性,要想得出全面规律还需调整林分密度范围、结合林龄及生境生态等因子进行综合研究分析。
植被演替的主要特征是物种组成和群落结构的变化,而更新层物种的演替是森林群落结构发生变化的主要驱动因子[18,19]。木本植物的更新对植物群落结构的优化以及群落的稳定性具有重要作用[20]。本研究地区麦吊云杉天然林中,仅4个林分密度下调查发现有自然更新幼苗,且数量较少,表明麦吊云杉自然更新能力极差。但在调查中发现 1 400 株·hm–2林分样方中存在大的林窗,该林窗下灌木和草本较少,且盖度较低,麦吊云杉种子掉落后,能够接触到土壤,增加其发芽、生长的机会,故而该林分虽然密度较大,但自然更新幼苗却最多。综合分析麦吊云杉天然林自然更新能力,为提高麦吊云杉更新能力,在实施保育措施时,应及时辅以人工辅助技术以提供其更新能力,提高遗传保育效果。
