基于网络药理学的生脉散用于COVID-19恢复期治疗的物质基础与作用机制研究
2020-11-02郭鹏飞方坚松刘艾林
郭鹏飞 方坚松 刘艾林

摘要 目的:利用網络药理学方法探索“生脉散(SMS)”用于新型冠状病毒肺炎(COVID-19)恢复期治疗的物质基础与作用机制。方法:结合中华人民共和国药典并从TCMSP、在线分析平台(ETCM、TCMID、HIT)以及Chemdb数据库搜索SMS复方中每味中药的归经和成分。药物成分潜在靶标通过PharmMapper、DrugBank、E-TCM和Swiss Target Prediction 4个数据库进行预测。然后,运用String数据库进行SMS药物成分靶标与COVID-19之间靶蛋白相互作用关系的分析,接着利用Cytoscape软件进行网络的构建和分析。最后用DAVID和FunRich数据库对候选靶标进行富集分析。结果:生脉散里的3味中药均归属于肺经,从生脉散复方中共筛选出21个符合选择标准的化学成分,主要包括常见的皂苷类化学成分如人参皂苷Rh4、醇类化学成分如β-谷甾醇,木脂素类化学成分如五味子素,另外还有生物碱类、黄酮类和萜类等化学成分。预测得到中药化学成分相关靶标457个,COVID-19靶标51个。通过String分析发现,化学成分中的70个靶标与COVID-19的29个靶标之间存在着紧密的相互作用;通路富集分析表明,候选靶标富集的通路主要包括癌症相关的多条通路、FoxO信号通路、病毒致癌作用信号通路等。结论:SMS中的有效活性成分可能通过作用于CUL2、NUP62、RAE1等多个靶点进而调节多条信号通路来起到抑制新型冠状病毒肺炎作用,研究结果将为进一步研究提供重要信息依据。
关键词 新型冠状病毒肺炎;生脉散;网络药理学;候选靶标;富集分析
Abstract Objective:To explore the material basis and mechanism of Shengmai San (SMS) in the treatment of COVID-19 during the recovery period using network pharmacology. Methods:The meridian tropism and composition of each herb in SMS were searched from TCMSP, online analysis platform (ETCM, TCMID, HIT), Chemdb database and chinese pharmacopoeia. Then, PharmMapper database, DrugBank, E-TCM and Swiss Target Prediction platforms were used to predict potential targets of components of SMS. The interaction between proteins of SMS drug components and the key targets of COVID-19 were analyzed by using String database, Cytoscape software was used for network construction and analysis. Finally, DAVID platform and FunRich databases were used for enrichment analysis of candidate targets. Results:It was found that the meridian tropism of the three Chinese herbs in SMS are all lung meridian. A total of 21 chemical components that meet the screen standard were selected from the SMS compound, including common saponin compounds such as ginsenoside Rh4, alcohol compounds such as β-sitosterol, lignan compounds such as schisandrin, and other chemical components such as alkaloids, flavonoids and terpenes. In addition, 457 targets related to the chemical compositions from SMS were predicted, and 51 targets related with COVID-19 disease were obtained. String analysis found that there were close interactions between the 70 targets of the chemical components and the 29 targets of COVID-19; The pathway enrichment analysis showed that the pathways enriched by candidate targets mainly included multiple cancer-related pathways, FoxO signaling pathway, viral carcinogenesis signaling pathway, etc. Conclusion:This paper preliminarily explored the material basis and action mechanism of SMS for the treatment of COVID-19 during the recovery period based on network pharmacology. These results will provide important information for further study.
Keywords COVID-19; Shengmai San (SMS); Network pharmacology; Candidate target; Enrichment analysis
中图分类号:R289.4;R563文献标识码:Adoi:10.3969/j.issn.1673-7202.2020.12.001
目前全球暴发的一种新型冠状病毒肺炎(Coronavirus Disease 2019,COVID-19)是由新型冠状病毒(SARS-CoV-2)引起的以呼吸道感染为主要特征的传染性疾病,临床上主要表现为发热、干咳和乏力等症状,目前尚无治疗特效药物。该病毒具有潜伏期长、传染性强、各类人群普遍易感等特点[1-4]。Wrapp等[5]证明SARS-COV-2的S蛋白和ACE2蛋白的结合强度约为SARS病毒结合ACE2蛋白的10~20倍。因此,如何形成快速有效的救治方案和研发有效的药物来遏制病毒的传播,是当前面临的重大问题。
在湖北确诊病例的临床治疗中,中医药对病毒感染性肺炎表现出来良好的治疗疗效,且参与率极高。同时,临床研究也证明,中药能够明显改善患者的发热、咳嗽、乏力和咽干等症状。中药的治疗对于患者的康复发挥着不可替代的优势。《新型冠状病毒感染的肺炎诊疗方案(试行第五版)》认为患者恢复期为肺脾气虚证,主要表现为气短、倦怠乏力、纳差呕恶、痞满、大便无力、便溏不爽、舌淡胖和苔白腻等症状[6]。基于此,生脉散及其制剂作为治疗气阴两虚证的代表方剂而被广泛推荐。《陕西省新型冠状病毒感染的肺炎中医药治疗方案(试行第二版)》中将生脉制剂作为患者恢复期辅助治疗的主要推荐用药[7]。《华中科技大学同济医学院附属同济医院关于新型冠状病毒肺炎中医诊疗方案及预防方案》报道,恢复期患者可采用生脉散合补中益气丸的联合治疗,以提高预防的效果[8]。
生脉散又名生脉饮,由人参、麦冬、五味子3味中药组成,方中人参补肺益脾以生津;麦冬养阴、消热以生津;五味子固表止汗而生津。张元素在《医学启源》中首载此方,朱震亨在《丹溪心法》中确立其治法。《医方考》谓之“一补,一清,一敛,养气之道备也,名曰“生脉”。《医学启源》卷下曰:“补肺中元气不足”。以上都说明此方在治疗肺病方面有足够的理论支持和依据[9]。
近年来,基于网络药理学进行药物重定位、利用人工智能(AI)助力药物研发以及应用深度学习方法设计疾病蛋白抑制剂等新技术新方法在药物研发的领域越来越扮演着重要的作用。Zhou等[10]利用系统药理学和网络医学方法系统的预测了2 000多个FDA批准的药物,筛选出了16种人类冠状病毒(HCoVs)的候选药物和3种潜在的药物组合,对于防治COVID-19老药新用的临床前研究和药物组发现有一定的指导价值和意义。Richardson等[11]运用机器学习模型,预测巴瑞替尼(Baricitinib)可能作为COVID-19急性呼吸系统疾病的潜在治疗药物。徐波研究团队研发了一个利用CT图像辅助诊断新型冠状病毒肺炎的人工智能模型。该模型鉴别新冠肺炎与其他病毒性肺炎的总准确率可达83%,且快速高效[12]。
我们利用网络药理学的方法从分子水平探索SMS用于治療COVID-19恢复期的机制。首先,通过多个数据库的挖掘并筛选SMS包含有效化学成分和它们对应的靶标,基于文献来总结COVID-19靶标的最新研究成果。然后,构建和分析药物-靶标网络,确定网络中的候选靶标并对关键靶标的分子功能、生物学过程以及通路等进行富集分析。最后,结合已有的药理学文献来探讨其中的机制,以期对生脉散用于治疗SARS-CoV-2感染患者恢复期的康复有所帮助和参考。
1 资料与方法
1.1 资料
从ETCM[13](TCM-MESH[14]、HIT[15]、TCM-ID[16])、TCMSP[17]和Chemdb数据库(http://www.organchem.csdb.cn.[1978-2020]Shanghai Institute of Organic Chemistry of CAS)中获取复方SMS所含3味中药的归经及包含的化学成分。
1.2 方法
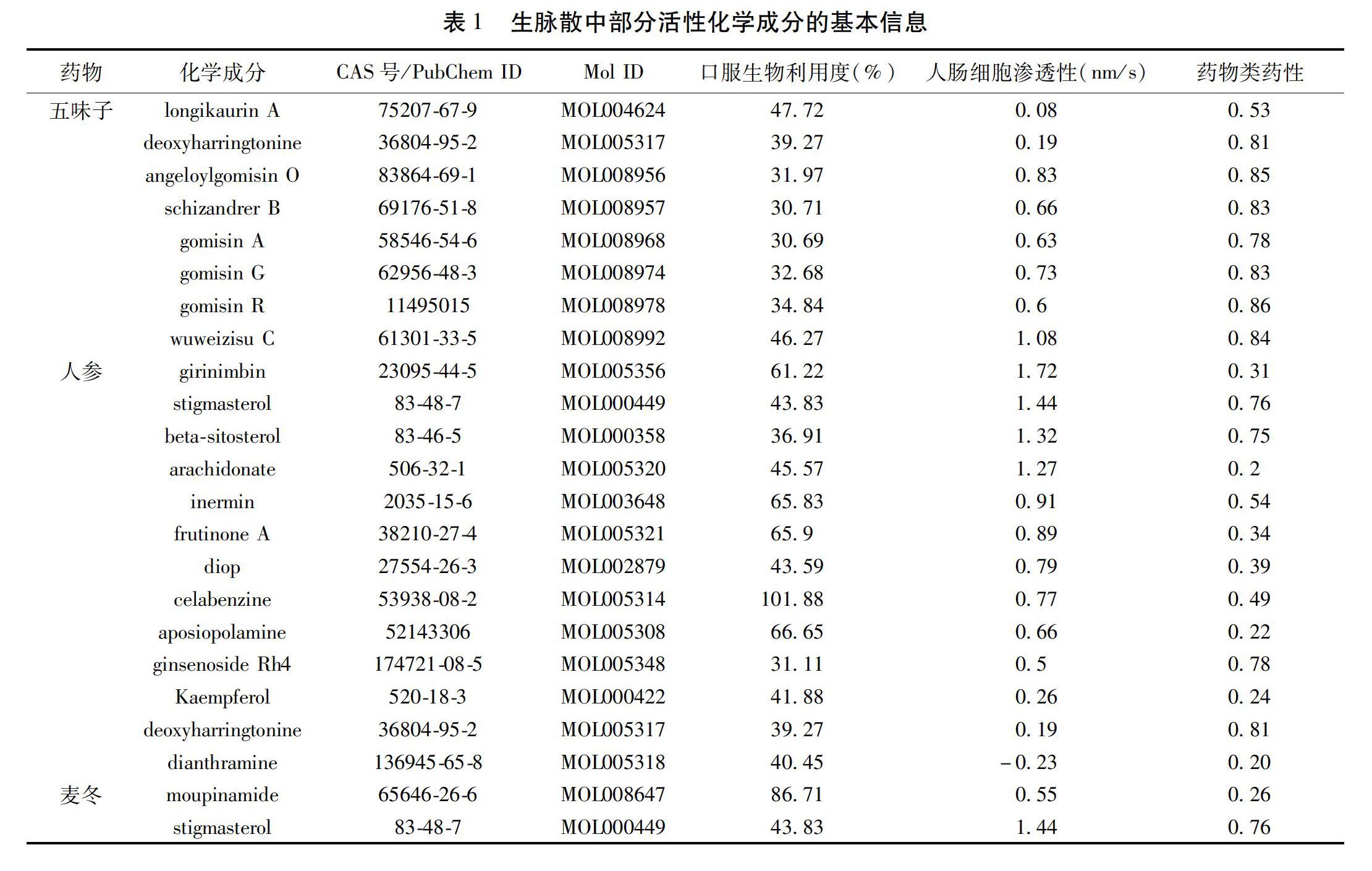
1.2.1 生脉散中药化学成分的收集 从ETCM[13](TCM-MESH[14]、HIT[15]、TCM-ID[16])、TCMSP[17]和Chemdb数据库(http://www.organchem.csdb.cn.[1978-2020]Shanghai Institute of Organic Chemistry of CAS)中获取复方SMS所含3味中药的归经及包含的化学成分,把所有的化学成分输入到TCMSP数据库进行口服生物利用度(OB),药物分子的类药性(DL)和人肠细胞株渗透性(Caco-2)的筛选。其中,OB表示口服药物有效成分或活性基被吸收到达体循环并被吸收的速度与程度。DL说明药物生物活性分子的类药性。Caco-2用作研究药物在肠道上皮中被动扩散的有效体外模型,我们使用Caco-2单层细胞中成分的转运速率(nm/s)来表示肠上皮渗透性。接着把收集到的所有化学成分在PubChem(https://pubchem.ncbi.nlm.nih.gov/)查找化学成分对应的CAS号(PubMed ID)。
1.2.2 生脉散中药化学成分靶标的预测 根据化学成分的名称从ETCM、PharmMapper[18]、TCMSP和Swiss Target Prediction[19]数据库进行靶标的预测。其中,ETCM里的靶标是用化合物设计与结构分析软件(MedChem Studio)预测且只保留可靠性数值大于0.8的靶标;PharmMapper预测的靶标选择(Norm_Fit)>0.7作为预测的靶标;TCMSP选择符合随机森林算法(Random Forest algorithm:http://www.stat.berkeley.edu/users/breiman)阈值≥0.8和支持向量机阈值(Support Vector Machine Score)≥0.7作为候选靶标;Swiss Target Prediction预测的靶标选择置信度(Probability)>0作为候选靶标.
1.2.3 疾病靶标的确定 本研究主要通过文献进行检索和汇总来确定疾病的靶标集。主要的方法包括药理学实验验证、快速筛查B谱系β冠状病毒受体结合域的方法、亲和纯化质谱(AP-MS)[20-24]。
1.2.4 靶标蛋白基因名的标准化及化学成分-靶点网络的构建 将预测到的化学成分和COVID-19的靶标运用UniProt(https://www.uniprot.org/)和GeneCards[25]数据库进行基因名的标准化,选择属性为人源蛋白。所有标准化后的靶标删除重复项之后分别作为作为药物靶标集和COVID-19的靶标集。然后,运用Cytoscape 3.2.1[26]软件构建和分析药物-靶点网络。
1.2.5 PPI网络的构建和候选靶点的筛选 为进一步明确潜在药物成分靶标和COVID-19靶点之间的相互作用关系,将筛选出来的化学成分靶点和COVID-19潜在的靶标导入String网络平台[27],选择最高的置信度(Highest_confidence)≥0.9作为条件进行PPI筛选,最终获得与COVID-19潜在蛋白的互作关系,并保存为TSV格式文件。将结果“#node1,node2”信息导入Cytoscape 3.2.1软件中,构建药物活性成分靶标和疾病靶标之间的PPI网络。
1.2.6 候选靶点的通路分析 为了进一步探讨PPI网络筛选出来候选靶标的分子功能、生物学过程、信号通路等机制,将筛选得到靶点导入DAVID数据库[28]进行KEGG富集分析[29],输入靶基因名称列表限定物种为“Homo Sapiens”,所有靶基因名称校正为“Official Gene Symbol”。并同时运用FunRich数据库进行信号通路、分子功能、和靶标归经等富集分析[30],运用在线绘图网站OmiShare Tools将其结果可视化[31]。
2 结果
2.1 SMS复方中有效成分的筛选结果 通过多个在线数据库的检索和收集,最后获得SMS复方中麦冬、人参和五味子中的化学成分共711个,其中80个来自麦冬,375个来自人参,256个来自五味子。以OB≥30%,DL≥0.18,Caco-2>-0.4为标准筛选出化学成分21个,其中2个来自麦冬,13个来自人参,8个来自五味子,其中人参和麦冬共有一个活性成分stigmasterol,人参和五味子共有一个成分 deoxyharringtonine。SMS中筛选的活性化学成分的基本信息见表1。
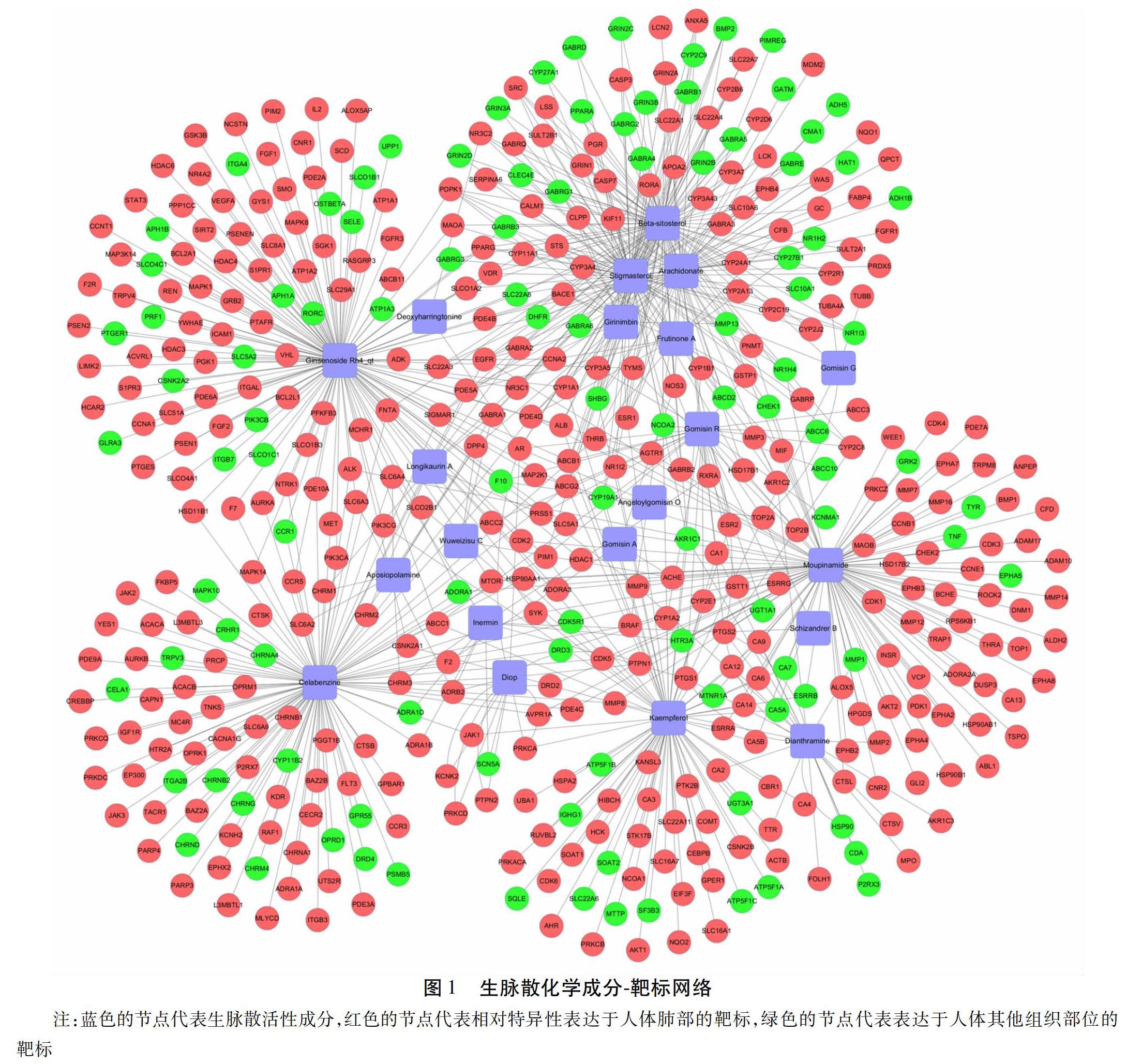
2.2 化合物成分-靶点网络图的构建 运用FunRich数据库对预测到的457个化学成分靶标进行人体表达部位的富集分析(site of expression),结果表明,所有预测的靶标中有430个靶标在肺部相对特异性的表达,表达部位包括肺、肺泡巨噬细胞、肺泡、支气管上皮细胞、支气管肺泡、胎肺、肺成纤维细胞和支气管,其他的靶标表达于T细胞、肝脏、直肠以及膀胱等部位。化合物-靶点网络见图1,图中共479个节点,808条边,红色的节点代表相对特异性表达于人体肺部的靶标,绿色的节点表示表达在人体其他组织部位的靶标,蓝色的节点代表化学成分。以中心度值(Betweenness Centrality)和等级值(Degree)对化学成分和靶点进行排序,排名前十的化学成分中心度值从大到小的化合物是 ginsenoside Rh4、dianthramine、gomisin R、arachidonate、kaempferol、beta-sitosterol、stigmasterol、moupinamide、celabenzine和frutinone A。Degree大于5的靶点有HSP90AB1、EGFR、ESR1、ABCB1、AR、CYP3A4和PTGS2等。在网络图中对应中心度值的变化与此节点的边所连接的Degree值呈正相关,Degree值越大,表明中心度值越高。
2.3 PPI网络的构建和候选靶标的筛选 PPI网络图的构建见图2,图中共涉及99个节点,137条边。红色的节点代表COVID-19靶标,蓝色的节点代表与COVID-19靶标直接相关的化学成分靶标。最后筛选出来99个核心靶标,见表3。其中,COVID-19靶標29个,药物活性成分的靶标有70个。COVID-19靶标和药物成分靶标相互作用打分前10的靶标
对分别是RAE1-NUP214、NUP62-NUPL1、NUP98-NUP54、RAE1-NUP98、RAE1-NUP54、CUL2-VHL、NUP62-NUP214、NUP62-NUP88、NUP62-NUP54和NUP88-NUP214。化学成分靶标和COVID-19靶标之间的PPI网络的分值(combined_score)见表2。本研究选择以上99个候选靶标作为SMS预防和治疗COVID-19恢复期的关键靶点。
2.4 候选靶标的KEGG富集分析 通过DAVID数据库对99个候选靶点进行KEGG功能富集分析,最后富集得到77条信号通路(P<0.05),涉及传染病的通路有9条:Tuberculosis(hsa05152:P=1.5×10-3)、Epithelial cell signaling in Helicobacter pylori infection(hsa05120:P=9.2×10-3)、Toxoplasmosis(hsa05145:P=0.046)、Influenza A(hsa05164:P=1.24×10-6)、Hepatitis B(hsa05161:P=3.93×10-4)、Hepatitis C(hsa05160:P=1.2×10-3)、Epstein-Barr virus infection(hsa05169:P=3.8×10-3)、Measles(hsa05162:P=5.9×10-3)和Herpes simplex infection(hsa05169:P=0.025)。涉及的基因有MAPK1、CEBPB、EP300、ATP6AP1、MAPK14、CREBBP、RAF1、JAK2、SRC等。将上述的通路输入到KEGG数据库进行通路的系统分类,以上9条通路都属传染病系统,其他通路的归属见表4。
2.5 候选靶标的FunRich富集分析 本研究选择富集结果最显著的前20个条目进行分析,分析结果集中于生物学过程,涉及信号转导、抗凋亡、酶活性的负调节、能量途径、炎性反应、蛋白水解和肽水解、蛋白质修饰等;分子功能集中于蛋白质丝氨酸/苏氨酸激酶活性、转运活动、跨膜受体蛋白酪氨酸激酶活性、激酶结合、半乳糖苷酶活性、羧肽酶活性、氧化还原酶活性、配体依赖性核受体活性等多种蛋白酶的活性;细胞组分中所涉及细胞质、胞质溶胶、核孔、线粒体、溶酶体、核包膜、Sin3复合体、组蛋白脱乙酰酶复合物等组分;通路富集分析主要集中于HDAC I类介导的信号传导、雄激素介导的信号传导、整合素相关激酶信号转导、AP-1转录因子网络、IFN-γ通路、视网膜母细胞瘤蛋白的调节、HDAC II类介导的信号传导、NEP/NS2与细胞出口机制的相互作用等。见图3。
2.6 候选靶标归经和临床表型的分析 归经和临床表型富集结果表明,候选的99个靶标中,在肺部相对特异性表达的靶标有AAK1、ABCC1、ACE2、ATP6AP1、AVPR1A、BRD2、BRD4、COMT、CUL2、ERC1等,共88个,占候选靶标总数的88.9%,说明SMS主要活性成分的靶标主要在肺部表达。同时,候选靶标还在人体的子宫颈、前列腺、子宫内膜、肝、直肠、胎盘、宫颈癌、膀胱和睾丸等组织表达。临床表型富集分析结果见图4。富集结果提示,SMS作用的候选靶标可能参与毛细血管瘤、红细胞增多症、血细胞比容增加、血红蛋白增加、红细胞量增加、排尿困难、肺泡细胞癌、肺癌、急性坏死性脑病和脑干病变等临床表型来预防COVID-19和其并发症的作用。
3 讨论
在此次疫情防治工作中,中医药显示了独特的优势,也取得了重要的成果。生脉散是治疗气阴两虚证的经典方剂,现在临床上仍然开发成多种试剂(包括散剂、颗粒剂、口服液、注射剂等)被广泛应用。《素问·至真要大论》曰:“肺苦气上逆,急食苦以泄之”“其泻以辛,其补以酸”“肺欲收,急食酸以收之”。方中人参味甘而微苦,补脾不腻之良品为君药。人参并非直接补肺,而是培土以生金,健母以养子耳。而麦冬甘寒,滋养肺阴,生津解渴用以为臣;五味子酸温,敛肺止汗,生津止渴,为佐药。三药合用,一补一润一敛,益气养阴,生津止渴,故名“生脉”。现代研究也证明,生脉散有治疗肺部疾病的作用。张心月等[32]通过对博来霉素诱导的大鼠肺纤维化模型的作用研究,发现人参、麦冬等药材可能参与机体炎性反应与免疫应答,通过减少促炎性反应因子的表达,最后显著降低大鼠肺纤维化程度。田野等[33]通过对生脉散的临床应用及其抗肺纤维化和血管内皮细胞损伤的药理作用进行综述,从而证明生脉散用于新型冠状病毒肺炎恢复期治疗的可行性。单卓程等[34]认为新型冠状病毒肺炎危重症期属内闭外脱者,治疗方可选生脉散加减。国家中医药管理局新冠肺炎防治方案(第5版)所载治疗方药当中,SMS的3味中药人参、麦冬和五味子都是治疗肺病使用频次最高的首选药物,其中,人参、五味子常用于补肺法,麦冬常用于清肺法[35]。
从网络药理学分析结果来看,生脉散的主要活性成分的靶标和SARS-CoV-2潜在的靶标存在紧密的相互作用关系。研究表明,在主要的活性成分中,豆甾醇(stigmasterol)可以调节卵清蛋白诱导的哮喘豚鼠模型中的过敏性气道炎性反应,具有显著的抗哮喘特性[36]。β-谷甾醇(beta-sitosterol)是唯一能在纳摩尔浓度下显着抑制铜绿假单胞菌依赖性IL-8表达的化合物。初步的机制分析表明,β-谷甾醇可以抑制铜绿假单胞菌依赖性蛋白激酶C亚型的活化,已知该活化作用与跨膜信号激活支气管上皮细胞中IL-8基因的表达有关。这些数据表明,β-谷甾醇是控制支气管纤维化患者过度肺部炎性反应的分子[37]。同样,预测的候选靶标在病毒的预防当中发挥着重要的作用。Vanessa Monteil等[38]证明临床级可溶性人ACE2有效抑制SARS-CoV-2在Vero E6、人血管类器官和人肾脏类器官的感染,或将有助于控制SARS-CoV-2感染。Zhao等[39]对715例受试者(415例,300例对照)使用多因素降维和累积遗传风险评分方法分析了基因与基因之间的相互作用,最后研究结果表明,ACE rs4340D/NOS3 rs1799983T的组合有助于中國儿童支原体肺炎性肺炎(MPP)的遗传易感性。
候选靶标的通路富集结果表明,COVID-19的发生发展和特定种类的癌症、多种细胞因子的表达、神经系统的调控、机体免疫反应以及破坏人类多能干细胞密切相关。
3.1 特定种类的癌症 Xiaodong Jia等[40]通过大数据分析发现特定种类癌症更具新冠病毒的易感性,对这些人群作出重点保护将是防疫工作的关键环节。本研究中预测的SMS的主要成分arachidonate、girinimbin、beta-sitosterol和kaempferol都具有减轻肺癌细胞损伤的作用,揭示了它们潜在预防COVID-19并发症的作用[41-45]。另外,DAVID和FunRich富集分析也发现候选靶标富集在多条癌症的信号通路,包括 hsa05200、hsa05203、hsa05205等。见表4。参与的基因有EGFR、PTGER1、AR、HSP90AA1、BRAF、MAP2K1、VHL、CREBBP、PPARG、RAF1、BCL2L1、IGF1R、CUL2等。
3.2 多种细胞因子的表达 Pei等[46]发现(SARS-CoV)、(MERS-CoV)以及其他呼吸道病毒如鼻病毒Rhinovirus和甲型流感病毒H1N1均可以诱导COVID-19细胞受体ACE2的上调。细胞因子如干扰素IFN-beta和IFN-gamma也可以刺激ACE2的表达,这表明SARS-CoV-2感染导致的“细胞因子风暴”不仅可以对人体器官造成损伤还可以促进病毒的进一步感染和传播。并推测病毒感染后激活免疫系统并诱导包括IFN在内的多种细胞因子表达,这些细胞因子通过其受体激活下游信号通路来促进ACE2的转录和表达。本研究通过FunRich富集分析发现,多个关键靶标如ABCC1、ERC1、HDAC2、NDUFAF2、NUP214、PRKACA、PTGES2、RIPK1、TMPRSS2等参与干扰素IFN-gamma pathway(P-Value=9.34×10-10)通路。见图3。
3.3 神经系统的调控 Mao等[47]对214名确诊为冠状病毒SARS-CoV-2感染并诊断出为严重急性呼吸道综合征(SARS)的住院患者进行了研究,其中88位(41.1%)为重度患者,126位(58.9%)为非重度患者。研究发现,与非严重的COVID-19患者相比,严重的患者通常具有神经系统症状,具体表现为急性脑血管疾病,意识障碍和骨骼肌症状。因此,对于COVID-19患者,除了呼吸系统症状外,医生还应密切注意任何神经系统表现;Xu等[48]从一名有明显中枢神经症状的SARS患者的尸体解剖获得的脑组织样本中成功分离出SARS-CoV株;Lau等[49]从一名SARS患者的脑脊液样本中也检测出SARS冠状病毒(SARS-CoV),表明SARS-CoV可能导致了中枢神经系统(CNS)的感染。本研究中,KEGG和FunRich通路富集分析结果均提示,药物作用的关键靶标参与多条神经系统的调控,其中KEGG的调控通路有 hsa00590(Arachidonic acid metabolism)、hsa04720(Long-term potentiation)hsa04726(Serotonergic synapse)、hsa04722(Neurotrophin signaling pathway)、hsa04730(Long-term depression)、hsa04725(Cholinergic synapse),涉及的基因有CBR1、PTGES2、PTGES、HPGDS、MAPK1、EP300、BRAF、MAP2K1、CREBBP、RAF1、PRKACA等。FunRich富集得到的神经相关通路如Neurotrophic factor-mediated Trk receptor signaling,参与的基因有DNM1、PIK3CA、YWHAE、PRKCZ、MAPK1、RAF1、SRC、BRAF、MAP2K1、MAPK14等。
3.4 机体免疫反应 近日,Grifoni等[50]首次分析了针对新型冠状病毒的机体有效免疫反应的潜在靶点,发现B细胞和T细胞表位在SARS-CoV-2和SARS-CoV之间具有高度保守性,靶向作用这些区域的疫苗策略或许能够帮助机体产生特殊的免疫力,不仅能提供交叉感染的保护能力,还能有效抵御病毒未来的进化。KEGG通路富集分析也表明,新冠病毒与免疫系统密切相关,如 hsa04611(Platelet activation)、hsa04062(Chemokine signaling pathway)、hsa04664(Fc epsilon RI signaling pathway)、hsa04620(Toll-like receptor signaling pathway)、hsa04660(T cell receptor signaling pathway)。涉及的基因有MAPK1、PRKCZ、MAPK14、PIK3CA、PRKACA、NOS3、SRC、ITGA2B等。
3.5 破坏人类多能干细胞 Lin等[51]研究表明,SARS-CoV-2破坏人类多能干细胞并将其转变为成纤维细胞,这为SARS-CoV-2的致病机制提供了新的见识。在本研究中,通过对候选靶标KEGG富集分析发现,基因MAPK1-IGF1R-MAP2K1-MAPK14-RAF1-PIK3CA-JAK2参与调控 hsa04550(Signaling pathways regulating pluripotency of stem cells)信号通路。
通过长期的实践从疗效观察中总结,中医的辨证用药讲究归经,归经指药物作用及药效在人体的脏腑经络的定位。根据中医理论,SMS复方中的3味中药都归属于肺经,为了证明药物中的关键成分作用靶标的归属,FunRich数据库富集说明了候选靶标在人体许多组织中的表达。而目前的临床研究也证明了靶标除了在肺部也可能在人体其他部位表达。例如,Mengyuan等[52]利用生物信息学方法研究了ACE2的组织分布,研究表明ACE2除了在肺組织表达外,还在胃肠道、肝脏、睾丸、肾脏、心脏和甲状腺等多个组织高表达,表明SARS-CoV-2可能会感染肺以外的其他组织。临床研究也表明,ACE2在男性的睾丸和女性的胎盘的表达量非常高。Cai等[53]研究表明,ACE2在睾丸中的表达量几乎是人体内最高的,并由此推测SARS-CoV-2很可能通过与这些ACE2阳性细胞结合,入侵并损伤患者的睾丸组织。Bo等[54]收治85例经实验室确认的COVID-19的患者的估计肾小球滤过率(eGFR)以及其他临床参数进行了回顾性分析,结果发现SARS-CoV-2可诱导COVID-19患者发生急性肾衰竭(ARF),病毒直接感染人肾小管以引起急性肾小管损伤。Dong等[55]研究报道了一名携带有COVID-19的孕妇分娩出新生儿2 h后,抗体IgM水平升高,细胞因子测试结果异常,并出现炎性反应和肝损伤的并发症状。IgM抗体水平升高表明该新生儿在子宫感染了病毒,同时间接支持垂直传播的可能性;Zeng等[56]发现在6名确诊COVID-19的孕妇中,有2例出生的婴儿中检测到IgM(通常不会从母亲转移到胎儿,因为其较大的大分子结构),这表明也存在母婴垂直传播的可能;同时,Kimberlin等[57]系统总结了以上2个研究成果,同时指出支持SARS-CoV-2的垂直传播。以上研究证明,SARS-CoV-2主要造成人体肺部的感染,同时可能引发多种并发症。SMS可能通过调控多个靶标来预防和治疗COVID-19恢复期的复发以及多种并发症的发生发展。
综上所述,本研究运用网络药理学方法,系统地研究了SMS治疗COVID-19恢复期的有效成分、潜在作用靶标及其网络作用机制,其中部分结果与最新文献报道相一致。SMS的有效成分是其发挥COVID-19康复期治疗作用的物质基础,将为药物治疗的研发提供有益的信息,关键靶标之间相互作用以及其相关通路研究对于揭示SMS的药效机制提供新的思路。
参考文献
[1]Wang W,Tang J,Wei F.Updated understanding of the outbreak of 2019 novel coronavirus(2019-nCoV)in Wuhan,China[J].J Med Virol,2020,92(4):441-447.
[2]苗青,丛晓东,王冰,等.新型冠状病毒肺炎的中医认识与思考[J].中医杂志,2020,61(4):286-288.
[3]Wu F,Zhao S,Yu B,et al.A new coronavirus associated with human respiratory disease in China[J].Nature,2020,579(7798):265-269.
[4]Wang C,Horby PW,Hayden FG,et al.A novel coronavirus outbreak of global health concern[J].Lancet,2020,395(10223):470-473.
[5]Wrapp D,Wang N,Corbett KS,et al.Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation[J].Science,2020,367(6483):1260-1263.
[6]新型冠状病毒感染的肺炎诊疗方案(试行第五版)[J].江苏中医药,2020,52(2):封4,封3.
[7]马战平,阴智敏,魏耕树,等.陕西省新型冠状病毒感染的肺炎中医药治疗方案(试行第二版)[J].陕西中医,2020,41(3):275-277.
[8]华中科技大学同济医学院附属同济医院中醫科,华中科技大学同济医学院中西医结合研究所.华中科技大学同济医学院附属同济医院新型冠状病毒肺炎中医诊疗方案及预防方案[J].医药导报,2020,39(3):308-309.
[9]张群智.古方生脉散现代应用探讨[J].中成药,2001,23(4):286-288.
[10]Zhou Y,Hou Y,Shen J,et al.Network-based drug repurposing for novel coronavirus 2019-nCoV/SARS-CoV-2[J].Cell Discov,2020,6:14.
[11]Richardson P,Griffin I,Tucker C,et al.Baricitinib as potential treatment for 2019-nCoV acute respiratory disease[J].Lancet,2020,395(10223):e30-30e31.
[12]Shuai W,Bo K,Jin lM,et al.A deep learning algorithm using CT images to screen for Corona Virus Disease(COVID-19)[J/OL].medRxiv(BMJ Yale),[2020-04-24].https://www.medrxiv.org/content/10.1101/2020.02.14.20023028v5.
[13]Xu HY,Zhang YQ,Liu ZM,et al.ETCM:an encyclopaedia of traditional Chinese medicine[J].Nucleic Acids Res,2019,47(D1):D976-976D982.
[14]Zhang RZ,Yu SJ,Bai H,et al.TCM-Mesh:The database and analytical system for network pharmacology analysis for TCM preparations[J].Sci Rep,2017,7(1):2821.
[15]Ye H,Ye L,Kang H,et al.HIT:linking herbal active ingredients to targets[J].Nucleic Acids Res,2011,39(Database issue):D1055-1059.
[16]Huang L,Xie D,Yu Y,et al.TCMID 2.0:a comprehensive resource for TCM[J].Nucleic Acids Res,2018,46(D1):D1117-1117D1120.
[17]Ru J,Li P,Wang J,et al.TCMSP:a database of systems pharmacology for drug discovery from herbal medicines[J].J Cheminform,2014,6:13.
[18]Wang X,Shen YH,Wang SW,et al.PharmMapper 2017 update:a web server for potential drug target identification with a comprehensive target pharmacophore database[J].Nucleic Acids Res,45(W1):W356-W360.
[19]Yuan Y,Pei J,Lai L.Binding site detection and druggability prediction of protein targets for structure-based drug design[J].Curr Pharm Des,2013,19(12):2326-2333.
[20]Zhou P,Yang XL,Wang XG,et al.A pneumonia outbreak associated with a new coronavirus of probable bat origin[J].Nature,2020,579(7798):270-273.
[21]Letko M,Marzi A,Munster V.Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B betacoronaviruses[J].Nat Microbiol,2020,5(4):562-569.
[22]Hoffmann M,Kleine-Weber H,Schroeder S,et al.SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor[J].Cell,2020,181(2):271-280.e8.
[23]Hoffmann M,Kleine-Weber H,Krüger N,et al.The novel coronavirus 2019(2019-nCoV)uses the SARS-coronavirus receptor ACE2 and the cellular protease TMPRSS2 for entry into target cells[J/OL].bioRxiv,[2020-O1-31].https://www.biorxiv.org/content/10.1101/2020.01.31.929042v1.
[24]Gordon DE,Jang GM,Bouhaddou M,et al.A SARS-CoV-2-Human Protein-Protein Interaction Map Reveals Drug Targets and Potential DrugRepurposing[J/OL].Nature,[2020-04-30].https://www.nature.com/articles/s41586-020-2286-9?utm_source=other&utm_medium=other&utm_content=null&utm_campaign=JRCN_2_DD01_CN_NatureRJ_article_paid_XMOL.
[25]Stelzer G,Rosen N,Plaschkes I,et al.The GeneCards Suite:From Gene Data Mining to Disease Genome Sequence Analyses[J].Curr Protoc Bioinformatics,2016,54:1.30.1-1.30.33.
[26]Demchak B,Otasek D,Pico AR,et al.The Cytoscape Automation app article collection[J].F1000Res,2018,7:800.
[27]Szklarczyk D,Gable AL,Lyon D,et al.STRING v11:protein-protein association networks with increased coverage,supporting functional discovery in genome-wide experimental datasets[J].Nucleic Acids Res,2019,47(D1):D607-607D613.
[28]Huang dW,Sherman BT,Lempicki RA.Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources[J].Nat Protoc,2009,4(1):44-57.
[29]Rossner P,Vrbova K,Rossnerova A,et al.Gene Expression and Epigenetic Changes in Mice Following Inhalation of Copper(II)Oxide Nanoparticles[J].Nanomaterials(Basel),2020,10(3):E550.
[30]Pathan M,Keerthikumar S,Ang CS,et al.FunRich:An open access standalone functional enrichment and interaction network analysis tool[J].Proteomics,2015,15(15):2597-601.
[31]Tai H,Opitz N,Lithio A,et al.Non-syntenic genes drive RTCS-dependent regulation of the embryo transcriptome during formation of seminal root primordia in maize(Zea mays L.)[J].J Exp Bot,2017,68(3):403-414.
[32]張心月,张丹,李慧,等.丹参、人参、连翘、麦冬对肺纤维化模型大鼠肺组织中BMP-4表达的影响[J].山东中医杂志,2019,38(11):1062-1066,1074.
[33]田野,李瑞明,任紅微,等.生脉散用于新型冠状病毒肺炎恢复期治疗的可行性探讨[J].药物评价研究,2020,43(3):378-383.
[34]单卓程,叶人,袁拯忠,等.温州地区中西医结合治疗新型冠状病毒肺炎体会[J/OL].新中医:1-7.[2020-05-09].http://kns.cnki.net/kcms/detail/44.1231.R.20200317.1519.002.html.
[35]周铭心,王苗.新型冠状病毒肺炎中医防治策略与方药筛选[J].中医学报,2020,35(3):458-463.
[36]Antwi AO,Obiri DD,Osafo N,et al.Stigmasterol Modulates Allergic Airway Inflammation in Guinea Pig Model of Ovalbumin-Induced Asthma[J].Mediators Inflamm,2017,2017:2953930.
[37]Lampronti I,Dechecchi MC,Rimessi A,et al.β-Sitosterol Reduces the Expression of Chemotactic Cytokine Genes in Cystic Fibrosis Bronchial Epithelial Cells[J].Front Pharmacol,2017,8:236.
[38]Monteil V,Kwon H,Prado P,et al.Inhibition of SARS-CoV-2 Infections in Engineered Human Tissues Using Clinical-Grade Soluble Human ACE2[J].Cell,2020,S0092-8674(20)30399-8.
[39]Zhao J,Zhang W,Shen L,et al.Association of the ACE,GSTM1,IL-6,NOS3,and CYP1A1 polymorphisms with susceptibility of mycoplasma pneumoniae pneumonia in Chinese children[J].Medicine(Baltimore),2017,96(15):e6642.
[40]Jia X,Yin C,Lu S,et al.Two Things about COVID-19 Might Need Attention.Preprints[J/OL].Preprint,[2020-02-23].https://www.preprints.org/manuscript/202002.0315/v1.
[41]Chen J,Tong W,Liao MX,et al.Inhibition of arachidonate lipoxygenase 12 targets lung cancer through inhibiting EMT and suppressing RhoA and NF-κB activity[J].Biochem Biophys Res Commun,2020,524(4):803-809.
[42]Rajavel T,Banu PG,Suryanarayanan V,et al.Daucosterol disturbs redox homeostasis and elicits oxidative-stress mediated apoptosis in A549 cells via targeting thioredoxin reductase by a p53 dependent mechanism[J].Eur J Pharmacol,2019,855:112-123.
[43]Mohan S,Abdelwahab SI,Cheah SC,et al.Apoptosis Effect of Girinimbine Isolated from Murraya koenigii on Lung Cancer Cells In Vitro[J].Evid Based Complement Alternat Med,2013,2013(10):689865.
[44]Vundru SS,Kale RK,Singh RP,et al.β-Sitosterol induces G1 arrest and causes depolarization of mitochondrial membrane potential in breast carcinoma MDA-MB-231 cells[J].BMC Complement Altern Med,2013,13:280.
[45]Govindaraju S,Roshini A,Lee MH,et al.Kaempferol conjugated gold nanoclusters enabled efficient for anticancer therapeutics to A549 lung cancer cells[J].Int J Nanomedicine,2019,14:5147-5157.
[46]Pei HW,Yun C.Increasing Host Cellular Receptor-Angiotensin-Converting Enzyme 2(ACE2)Expression by Coronavirus may Facilitate 2019-nCoV Infection[J/OL].bioRxiv,[2020-02-27].https://www.biorxiv.org/content/10.1101/2020.02.24.963348v1.
[47]Mao L,Jin H,Wang M,et al.Neurologic Manifestations of Hospitalized Patients With Coronavirus Disease 2019 in Wuhan,China[J].JAMA Neurol,2020 Apr 10;e201127.
[48]Xu J,Zhong S,Liu J,et al.Detection of severe acute respiratory syndrome coronavirus in the brain:potential role of the chemokine mig in pathogenesis[J].Clin Infect Dis,2005,41(8):1089-1096.
[49]Lau KK,Yu WC,Chu CM,et al.Possible central nervous system infection by SARS coronavirus[J].Emerg Infect Dis,2004,10(2):342-344.
[50]Grifoni A,Sidney J,Zhang Y,et al.A Sequence Homology and Bioinformatic Approach Can Predict Candidate Targets for Immune Responses to SARS-CoV-2[J].Cell Host Microbe,2020,27(4):671-680.e2.
[51]Lin Z,Wu Z,Mai J,et al.The Nucleocapsid Protein of SARS-CoV-2 Abolished Pluripotency in Human Induced Pluripotent Stem Cells[J/OL].bioRxiv,[2020-05-01].https://www.biorxiv.org/content/10.1101/2020.03.26.010694v3.full.
[52]Mengyuan L,Lin L,Yue Z,et al.Expression of the SARS-CoV-2 cell receptor gene ACE2 in a wide variety of human tissues[J].Infect Dis Poverty,2020,9(1):45.
[53]Cai BF,Kai L,Yan HD,et al.ACE2 Expression in Kidney and Testis May Cause Kidney and Testis Damage After 2019-nCoV Infection[J/OL].medRxiv(BMJ Yale),[ 2020-02-13].https://www.medrxiv.org/content/10.1101/2020.02.12.20022418v1.
[54]Bo D,Chenhui W,Rong SW,et al.Human Kidney is a Target for Novel Severe Acute Respiratory Syndrome Coronavirus 2(SARS-CoV-2)Infection[J/OL].medRxiv(BMJ Yale),[ 2020-04-10].https://www.medrxiv.org/search/Human%252BKidney%252Bis%252Ba%252BTarget%252Bfor%252BNovel%252BSevere%252BAcute%252BRespiratory%252BSyndrome%252BCorona-virus%252B2%252B%2528SARS-CoV-2%2529%252BInfection.
[55]Dong L,Tian J,He S,et al.Possible Vertical Transmission of SARS-CoV-2 From an Infected Mother to Her Newborn[J].JAMA,2020,323(18):1846-1848.
[56]Zeng H,Xu C,Fan J,et al.Antibodies in Infants Born to Mothers With COVID-19 Pneumonia[J].JAMA,2020,323(18):1848-1849.
[57]Kimberlin DW,Stagno S.Can SARS-CoV-2 Infection Be Acquired In Utero More Definitive Evidence Is Needed[J].JAMA,2020,323(18):1788-1789.
(2020-05-12收稿 責任编辑:杨觉雄)
