运用稳定同位素技术分析三峡坝前水域的食物网结构
2020-10-31周正黄宇波王斌梁米武娟宋庆洋许元钊毕永红
周正,黄宇波,王斌梁,米武娟,宋庆洋,许元钊,毕永红,*
1. 中国科学院水生生物研究所,武汉 430072
2. 中国科学院大学,北京 100049
0 前言
稳定同位素是天然存在于自然界中且不具有放射性的一类同位素,其细微的质量差别使其显示出理化性质的差别,即同位素效应,该效应被广泛应用于生理生态研究中,稳定同位素分析技术可对研究对象进行定量研究[1]。其中稳定同位素13C、15N被运用得最为广泛,在捕食者和猎物中13C富集极小(约为0.8‰)[2],故δ13C值在捕食者与猎物中的差别不大,常用于食性分析[3];15N 在捕食者中发生一定的富集(3.4‰)[4],故可根据δ15N进行营养级判断。传统胃含物判断生态位只能反映捕食者较短时间内的摄食情况,且有时胃含物会因食物外部形态变化导致不易判断,而稳定同位素技术可准确定量较长时期的摄食情况,传统胃含物分析有逐渐被稳定同位素取代的趋势[5];通过稳定同位素分析河流、湖泊、海洋等水生生态系统已经越来越普遍[6][7],因为稳定同位素的分馏一般较小,所以用所测定物质的稳定同位素与国际公认标准物质稳定同位素的比值来更好表示分馏差异[8]: 如湖泊方面,Post(2002)等对美国30多个湖泊进行分析,探讨了影响食物链长度的因素,认为生态系统尺寸是食物链长度的决定因素[5],Vander Zanden等(1996)对加拿大 14个湖泊中影响消费者13C、15N 的原因进行了分析,并探讨了全球水域生态系统的食物链长度,确定了利用稳定同位素技术对食物链长度分析进行分析的可行性[6],Xu等(2005)利用稳定同位素技术对我国东湖等湖泊进行了食物网分析,分析了淡水湖泊中的食物网结构和营养级关系;在河流方面,Li等(2012)利用稳定同位素技术分析了三峡小江食物网结构并确认了蒙古鲌的食性转变[9],Ba等(2015)调查了长江中游鱼类的食物网结构,并分析了其中颗粒有机物(POM)的时空特征[10];在水库方面,Noboru Okuda等(2017)利用稳定同位素技术对台湾翡翠水库进行了研究[11],得出了水库储水后,消费者会从主要依赖底栖藻类作为食源变为主要依赖浮游藻类作为食源。相比于传统的食物网分析技术,稳定同位素技术在食物网分析时的定量测量优势较为明显,能够构建连续性的营养级,并且能够对小型鱼类或底栖动物等不容易通过胃含物方法进行分析的物种进行营养级和生态位分析。
三峡水库的水域生态情况自三峡大坝建坝以来就广为关注,其生态系统受大坝截留、人类活动、季节气候等因素影响[12],认识坝前和支流鱼类食物网结构差异对于了解大坝对生态系统的影响具有重要意义。坝前水域因为大坝的运行,其食物链长度、食物网冗余程度可能与支流水域存在差异,我们假设坝前水域具有较长的食物链长度和较高的食物网冗余程度。为了验证相关假设,我们选取坝前的茅坪水域和支流的香溪河水域作为代表,通过碳、氮稳定同位素技术,分析三峡坝前干流与香溪河支流的鱼类食物网结构存在的差异。
1 研究地点与方法
1.1 研究地点概况
三峡位于长江中上游,长约 660余千米,主要由山地和低山丘陵组成,属中亚热带湿润季风气候,年平均气温17—19oC,年降水1000—1200 mm。水库水质以Ⅲ类水为主,与国内其他水体相比,受挥发性有机物的污染较轻[37]。本次研究分别选取茅坪水域和香溪河水域分别作为坝前水域和支流水域的代表,茅坪水域位于三峡坝前,水域面积约 88 km2,其水位的变化受大坝运行的调控,水生生态系统结构受大坝影响明显,本研究在该水域雇用渔民通过设置网具采集相关样品用于分析;香溪河是三峡库区坝前的一级支流,系三峡水库湖北辖区内的最大支流,发源于神农架,其支流有南阳河、古夫河、高岚河三条支流,长度约 94 km,流域面积 3095 km2,随着坝前水位的抬升,距离香溪河河口32 km范围内形成回水区,河流由北向南,在秭归香溪汇入长江干流[13],该支流距离三峡大坝 33 km,本研究选取其作为坝前支流代表,并在该支流的峡口、吴家湾两地采集样品用于分析。其中茅坪镇为秭归县城所在地,人类活动较为频繁,茅坪镇人口8万;香溪河采样地点峡口镇人口4.3万。

图1 长江三峡干流及和香溪河支流采样区域示意图Figure 1 Sampling area of mainstream of Three Gorges Dam and Xiangxi river
1.2 样品采集与处理
全部样品采集于2013年。鱼类样品通过设置渔网等方式捕捞,坝前水域采样地点在茅坪附近,香溪河水域的样品于峡口、吴家湾两地采集。在称体重、鲜肉重、干重后,取其背部肌肉(体积较小的鱼类取三鱼混合碾磨),用冷冻干燥机干燥后用锡纸包裹,并用密封袋密封后避光保存;POM采用100 μm的浮游生物采集网收集并干燥。在需要测量时,将干燥后的样品取出部分,置于2 mL离心管中用高通量碾磨仪碾磨成粉末,取碾磨后的肌肉样品用锡杯包裹并称重,随后用稳定同位素质谱仪测定样品中的13C、15N,所用仪器为中科院水生生物研究所分析测试中心的稳定同位素质谱仪(Thermo Fisher Delta V Advantage)。采集鱼类样品的信息如下: 茅坪采集鱼类144尾,共19种;峡口采集鱼类208尾,共19种;吴家湾采集鱼类99尾,共15种; POM样品3个,采集地点分别为茅坪和峡口。
1.3 数据处理
1.3.1 稳定同位素数据处理
后期数据处理通过SPSS17.0,Excel2016等软件完成,绘图工具为 Originpro9.1,使用 Excel2016计算每种鱼类样品稳定同位素值的平均值和标准差及同位素量化指标,通过SPSS17.0分析两地不同食性鱼类的稳定同位素值得差异显著性情况。
碳氮稳定同位素分析分别采用国际标准物质PDB(Pee Dee Belnite)和标准大气氮对比。分析结果用δ13C、δ15N来表示,计算公式如下:
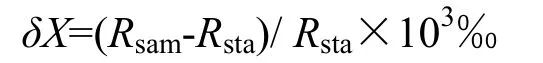
其中X代表13C或15N,R代表13C/12C或15N/14N。
1.3.2 鱼类营养位置关系及食物链长度计算
计算每种鱼类δ13C、δ15N的平均值与标准差,如表1、2所示,括号中的数值代表该种鱼类用于稳定同位素测定的样品量。鱼类相对营养位置关系(Trophic level)及食物链长度是基于POM计算,该试验采集的POM成分主要是初级生产者浮游植物,用如下公式进行计算
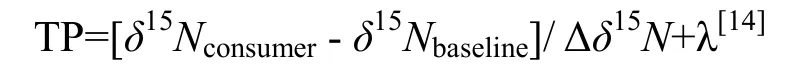
式中,Δδ15N为计算营养级的富集因子,其值为3.4‰,此时 baseline是初级生产者,λ=1[14],结果如图2所示。
一个生态系统的食物链长度(FCL)一般根据最高营养级消费者的δ15N进行计算,取决于其营养级,食物链长度计算方法如下: FCL=(最高营养级的Δδ15N-基准生物的Δδ15N) /3.4‰+a[27],其中a为基准生物的营养级(初级生产者=1,初级消费者=2),本研究采用POM作为基准生物,故a=1。
1.3.3 鱼类群落同位素量化指标计算
对两地水域的都存在的7种鱼类分析其营养结构的差异,通过δ13C-δ15N值双坐标图计算6个同位素量化指标来衡量两地鱼类群落的生态位空间特征[38]。虽然两地都存在的同种鱼类只有 7种,但是包括了植食性鱼类、杂食性鱼类、中上层肉食性鱼类、底栖肉食性鱼类等不同食性和不同生态位的鱼类,故具有一定代表性。六个指标分别为: 氮值范围NR(δ15N Range),碳值范围 CR (δ13C range),总面积 TA (Total area ),平均离心距离 CD (Mean distance to centroid),平均最邻近距离 NND( Mean nearest neighbor distance),最邻近距离标准差 SDNND (Standard deviation of nearest neighbor distance)。计算方法采用Layman等(2007)所使用的方法[38],所有计算基于δ13C-δ15N 值双坐标图,其中氮值范围和碳值范围通过该水域中13C、15N富集程度最高的鱼类的δ13C值和δ15N值减去中富集程度最低的鱼类的δ13C值和δ15N值;总面积通过计算δ13C-δ15N值双坐标图中各水域鱼类围成凸多边形面积得到;平均离心距离则通过计算同一水域中各鱼类距其中心点(即该处水域鱼类的δ13C、δ15N平均值)距离的平均值得到;平均最邻近距离通过计算该处水域每种鱼类和距离自己最近的鱼类距离的平均值得到;而最邻近距离标准差通过计算该处水域每种鱼类和距离自己最近的鱼类距离的标准差得到。
2 结果
2.1 δ13C测定结果
此次采集的 POM 测定其δ13C值为-23.99‰±2.88‰,样本中 POM 的δ13C 范围在-25.93‰—-20.67‰。从表1中可以看出,坝前干流水域鱼类的δ13C 范围是-33.5‰—-19.4‰,支流水域鱼类为-28.3‰—-13.9‰,坝前干流水域和支流水域同种鱼类的δ13C 值差别在(1‰—8.9‰)。
将鱼类按照食性分类进行分析,在坝前干流水域中,植食性鱼类(鳊鱼、草鱼等)与底栖杂食性鱼类(鲤鱼、鲫鱼等)的δ13C 没有显著差异(P>0.05),植食性鱼类与底栖肉食性鱼类(瓦氏黄颡鱼、光泽黄颡鱼等)之间的δ13C 具有显著差异(P<0.05),而底栖杂食性鱼类与底栖偏肉食性鱼类之间的δ13C具有极显著差异(P<0.01),而中上层肉食性鱼类(翘嘴鲌、鳡鱼等)与底栖肉食性鱼类之间的δ13C也具有显著差异(P<0.05);而在香溪河水域中,底栖杂食性鱼类与底栖偏肉食性鱼类之间的δ13C没有显著差异(P>0.05)。
此外,坝前干流水域和香溪河水域两地草鱼δ13C 相差较大(8.9‰),其他鱼类的δ13C 相差 0.77‰—3.77‰,两地的植食性鱼类δ13C、中上层肉食性鱼类的δ13C、底栖肉食性鱼类的δ13C均没有显著差异(P>0.05),两地草鱼δ13C 相差较大(8.9‰),其他鱼类的δ13C相差0.77‰—3.77‰。
2.2 δ15N测定结果
此次试验所采集的POM的δ15N值为4.52‰±1.12‰。表 1中,坝前干流鱼类的δ15N值范围是6.72‰—15.5‰,香溪河水域鱼类的δ15N 值范围是4.8‰—14.4‰。两处水域都采集到的鱼类样品中,坝前干流鱼类的δ15N 平均值(11.1‰)大于支流水域鱼类(10.1‰)。将鱼类按照食性分类进行分析,在坝前水域中,草鱼、鳊等植食性鱼类体内的15N富集程度最低,其次是鲤、鲫等底栖杂食性和鲢等滤食性鱼类,鳡、翘嘴鲌等肉食性鱼类样品的15N富集最多,其中植食性鱼类和底栖杂食性鱼类之间、底栖肉食性鱼类和中上层肉食性鱼类之间的δ15N值差异均不显著(P>0.05),而植食性鱼类和中上层肉食性鱼类的δ15N 差异极显著(P<0.01),底栖杂食性鱼类与底栖肉食性鱼类的δ15N 差异显著(P<0.05);在香溪河水域中,草鱼等植食性鱼类体内的15N 富集程度最低,其次是鲢等滤食性鱼类,斑鳜、鳡等肉食性鱼类样品的15N富集最多,其中植食性鱼类与中上层肉食性鱼类差异显著(P<0.05)。

表1 三峡水库坝前干支流水域鱼类的δ13C和δ15N值Table 1 δ13C and δ15N values of fish in the mainstream and tributary of the area in front of the Three Gorges Dam
2.3 相对营养位置
将鱼类按照食性分类进行分析,两地均为植食性鱼类处于低营养级,香溪河水域为草鱼(1.09),坝前干流水域为鳊鱼(1.65);中上层肉食性鱼类占据最高生态位,坝前干流水域为翘嘴鲌(4.17),香溪河水域为斑鳜(3.92)。经计算得到,香溪河水域鱼类的平均营养级为 2.64,食物链长度为 3.9,其中营养位置在 2—2.5之间的鱼类占总数的 41.7%,占据鱼类种类数的大部分;坝前干流水域鱼类的平均营养级为3.00,食物链长度为4.35。其中营养位置在2.5—3.5之间的占总数的52.6%。从图2关于两地鱼类营养层级的数据可看出,在两处水域中,鲤鱼、瓦氏黄颡鱼等杂食性鱼类的营养级较为接近,而翘嘴鲌、鳡等肉食性鱼类的营养级,茅坪水域则大于香溪河水域。

图2 两处水域鱼类营养级Figure 2 Trophic levels of fish in the mainstream and tributary of the area in front of the Three Gorges Dam
2.4 鱼类食物网结构及同位素量化指标
图3、图 4分别为坝前干流水域和香溪河水域的鱼类δ13C—δ15N图。由图可知,坝前干流水域生态系统中鱼类的δ13C 较为集中,在该生态系统中,鱼类的主要营养层可分为 3个,草鱼、鳊、鲢等在食物网中营养层级较低,鲫、贝氏餐、瓦氏黄颡鱼等杂食鱼类处于中间营养级,鳡和翘嘴鲌等肉食性鱼类占据上层生态位,其中鳡占据最高营养位置。其中中间层级鱼类竞争压力较大,生态位重叠较多;香溪河水域中,主要鱼类营养层能分为3个,草鱼、鲢等植食性和滤食性鱼类在食物网中营养层级较低,瓦氏黄颡鱼等杂食性鱼类处于中间营养层,而鳡鱼和斑鳜等肉食性鱼类处于最高营养层,其中斑鳜占据最高营养位置。

图3 坝前干流水域食物网结构图Figure 3 Food wed structure of the mainstream area in front of the Three Gorges Reservoir
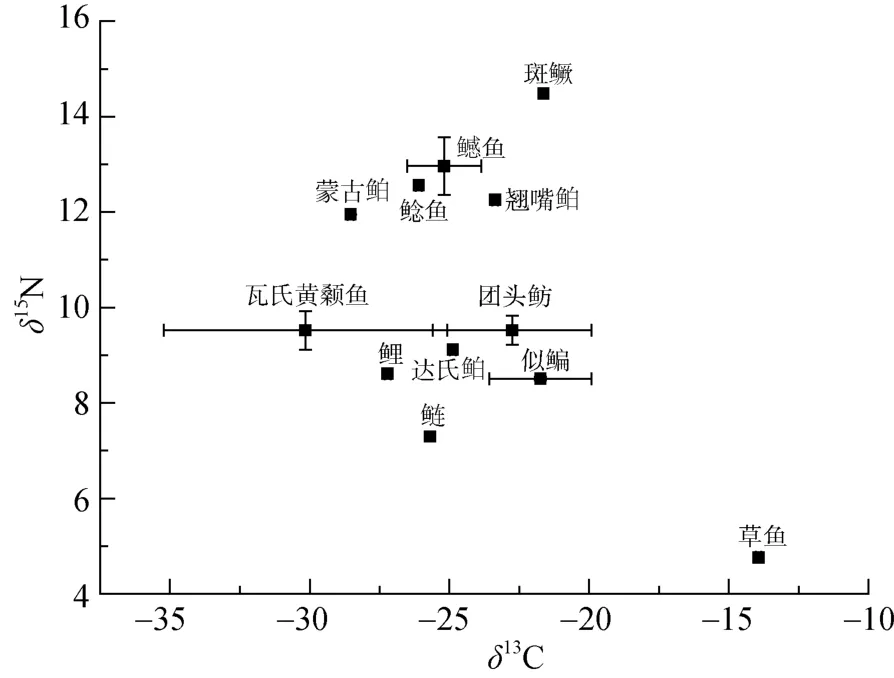
图4 香溪河水域食物网结构图Figure 4 Food wed structure of the Xiangxi River
对两地水域的同种鱼类分析其营养结构的差异,通过δ13C—δ15N值双坐标图计算6个同位素量化指标来衡量两地鱼类群落的生态位空间特征,其结果如表3所示。在计算得到的6个反应鱼类生态位空间特征的指标中,两处水域同种鱼类氮值范围 NR相差不大(0.37),但香溪河水域的碳值范围 CR远大于坝前干流水域(9.76),其中总面积 TA香溪河水域约是坝前干流水域的 3倍,说明香溪河水域鱼类的食源多样性可能大于坝前干流水域。香溪河水域的平均最邻近距离和最邻近距离标准差均大于坝前干流水域,说明香溪河水域的营养冗余程度低于坝前水域,另外两水域的平均离心距离相差不大。
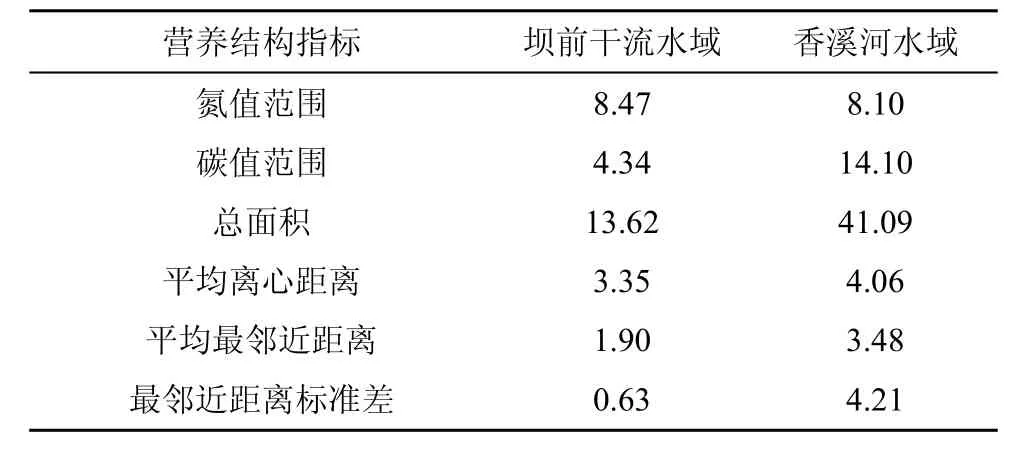
表3 两处水域同种鱼类群落同位素量化指标Table 3 Community-wide quantitative metrics of fish community of two waters

图5 两处水域同种鱼类群落营养结构图Figure 5 Trophic structure of the selected species over two waters
3 讨论
生态系统中捕食者的δ13C取决于其食物,δ13C能够在一定程度上指示鱼类的食物来源[19];13C的富集程度很小,鱼类背部肌肉组织中的δ13C值与其全身匀浆的δ13C近似相等,因此背部肌肉组织中δ13C值可反映鱼类食性[18]。坝前干流水域鱼类的δ13C大部分集中在-21‰—-27‰,占 78.9%,和坝前干流食物来源相对较为单一相关。坝前干流水域中鳊和草鱼的δ13C十分接近,与草鱼和鳊均为植食性鱼类相符[20];香溪河水域鱼类与坝前干流水域不同,底栖杂食性鱼类与底栖肉食性鱼类之间的δ13C不具有显著差异(P>0.05),可能是因为香溪河水域的底栖杂食性鱼类与底栖肉食性鱼类食源差异不大;另外香溪河水域草鱼的δ13C明显不同于其他鱼类,与香溪河水生植物较少[21],草鱼不是主要类群,也不是肉食鱼类的主要食物来源等因素有关。此外,坝前干流水域与香溪河水域中草鱼δ13C 相差很大(8.9‰),说明在坝前干流与支流草鱼具有不同的食物来源。综上所述,坝前干支流水域部分鱼类的食物来源存在一定的差异,这种食物来源的差异可能是导致干支流鱼类资源量差异的一个重要原因。
消费者和食物间的δ15N差异就是营养分馏(trophic fractionation)值,随着鱼类营养级增加 其δ15N不断增加,不同营养级间的15N富集值的平均值为 3. 4‰,因此可通过分析δ15N分析鱼类相对营养位置关系[19][24]。营养级关系是群落内各生物间重要的联系,分析营养级关系及食物链长度等是了解生态系统能量流动的核心[25-26]。已有研究认为污水排入等会增大水体中基础食源的δ15N[20][27]。对两地鱼类的δ15N值的如表1所示,坝前干流鱼类的δ15N平均值(11.1)大于支流水域的鱼类的δ15N 平均值(10.1),且就同种鱼类比较,坝前干流鱼类的δ15N均大于香溪河水域,可能是因为坝前水域所在地靠近县城,人类活动较为频繁会增大水体中基础食源的δ15N,进而导致该水域中鱼类的δ15N值偏大。在香溪河水域中斑鳜处于最高营养位置,已有研究表明,斑鳜较为凶猛,甚至存在同类相食的情况[28],斑鳜的这种食性会导致体内15N 的富集程度较高;在坝前干流水域中鳡鱼处于最高营养位置。
香溪河水域鱼类的平均营养级为 2.64,营养位置在 2—2.5之间的鱼类占总数的 41.7%,中间营养级的鱼类占总数的大部分;坝前干流水域鱼类的平均营养级为 3.00,营养位置在 2.5—3.5之间的占总数的 52.6%,与巴家文等对长江中游干流的分析情况相似[10],长江中游干流江段主要生物资源种类的营养级处于 2.42—4.88,集中在 2.83—3.61 之间,这些研究结果共同说明长江中上游的生态位较为拥挤。从图3和图4可见,两地均存在中间生态位较为拥挤的现象,而坝前水域较香溪河水域更为拥挤,与其所处水域位置中食物来源较少有关[29]。从关于两地鱼类营养层级的数据可看出,在两处水域中,鲤鱼、瓦氏黄颡鱼等杂食性鱼类的营养级较为接近,而翘嘴鲌、鳡等肉食性鱼类的营养级,坝前干流水域则大于香溪河水域,可能是由于坝前干流水域拥有更为稳定的食物网结构,从而使该处肉食性鱼类有足够的其他鱼类作为食物,进而拥有更高的营养级。在计算得到的六个生态学指标中,前四个指标(NR、CR、TA、CD)反映物种空间范围,代表群落营养多样性,后两个指标(NND、SDNND)反映物种相对位置,代表营养冗余[38]。香溪河水域的平均离心距离(CD)大于坝前水域,碳值范围(CR)大于坝前干流水域(9.76),但两处水域同种鱼类氮值范围(NR)相差不大(0.37),CR在一定程度上反应鱼类群落的食源的碳源范围,NR反映该水域的营养长度(食物网垂直结构),该结果说明两处水域的营养长度相近,但香溪河水域的基础食物源多样性大于坝前水域;另外,在反应物种相对位置的指标上,香溪河水域的平均最邻近距离值(NND)和最邻近距离标准差值(SDNND)均大于坝前干流水域,说明坝前干流水域的鱼类营养结构更为冗余,且分布较香溪河流域更为均匀,可认为坝前干流水域中不同鱼类之间的竞争相对于香溪河水域更为激烈[38]。Shahid等研究发现,一般情况下,较高的冗余程度有助于维持生态系统的稳定性[39],而林会洁等对拓林湾海洋牧区不同功能区的食物网结构分析中,也得出了人工鱼礁区的营养冗余程度较大故其生态系统稳定性更佳的结论[40],所以坝前水域的生态系统可能会因为其更高的营养冗余程度而更加稳定。
Vander Zanden等对全球219个水域生态系统食物链长度的文献进行分析研究,得出河流生态系统食物链长度平均为3.5的结论[27],在本研究中,坝前干流水域和香溪河水域的食物链长度分别为4.35与3.9,均大于河流生态系统食物链的平均长度,可能是由于两地均处于长江流域生态系统中,并且拥有相对其他河流水域生态系统较大的生态系统空间所导致。根据生态系统大小假说,大的生态系统可直接增加生态系统资源可获得性,通过物种定居与消失的可能性、物种多样性、栖息地可利用性与异质性、消费者杂食性、捕食者被捕食者之间的相互作用稳定性来影响捕食者猎物个体大小比例等依赖资源可获得性的种群特征,进而决定食物链长度[29]。另外,Hoeinghaus等对巴西UpperPara河的食物链长度的研究时,发现在水库中食物链长度最长,变幅在4.0—4.35之间,高比降河流中食物链长度最短[30],坝前干流水域鱼类的食物链长度(4.35)高于香溪河水域(3.9),与 Hoeinghaus的研究结果相符,这是由于坝前干流水域位于长江干流,生态环境受大坝蓄水、支流汇入等诸多因素的影响,生境多样性高,进而导致食物链长度大于香溪河水域[31]。基于动态稳定性假说,有研究认为一定程度的干扰会增加生态多样性,进而增长食物链长度[32-33],也有研究表明河流扰动会限制鱼类食物中次级消费者的丰度而缩短食物链长度[34-35],且鱼类在水文环境变化较大的环境中摄取到中间捕食者的可能性小于稳定环境,目前关于影响食物链长度的原因尚无定论[36][42][43],但在本研究中,坝前干流水域食物链长度大于香溪河水域,且鱼类营养级结构更为复杂,可认为坝前干流水域生态系统更为稳定,从而其有足够的时间发展出更长的食物链,并且形成稳定的鱼类食物网结构。
4 结论
结果表明坝前干流水域的食物链长度为4.35营养级,香溪河水域食物链长度为3.9营养级;坝前干流水域食物网结构较香溪河水域更为稳定;坝前干流水域不同鱼种之间的竞争较支流水域更为激烈,且坝前干流水域的鱼类营养结构较支流水域更为冗余,坝前干流和支流都存在中间生态位较为拥挤的现象。
致谢: 感谢中国科学院水生生物研究所分析测试中心的左艳霞老师、王敏老师在仪器使用方面提供的便利!
