GnRH基因SNPs 与鹅体组成和蛋品质性状的关联分析
2020-10-19霍子东高广亮张克山王启贵赵献芝
霍子东,高广亮,张克山,王启贵,赵献芝*
(1.甘肃省畜牧技术推广总站,甘肃兰州 730030;2.重庆市畜牧科学院,重庆 402460;3.重庆市肉鹅遗传改良工程技术研究中心,重庆 402460)
鹅作为重要水禽之一,具有抗逆性强、适应性好、耐粗饲、抗病性强和耐粗饲等特点,为广大消费者提供肉、蛋、羽绒、肥肝等。体组成和蛋品质性状是重要的经济性状,也是复杂的数量性状,且均受到遗传、营养、饲养方式、外界环境等影响[1-3]。利用生物学等方法进行研究,鸭体尺、鸡蛋壳颜色等分子标记被发现及广泛应用到分子标记辅助选择,极大地推进了禽类的体组成和蛋品质等方面的研究进展[4-6]。
促性腺激素释放激素(Gonadotropin-releasing Hormone,GnRH)是动物下丘脑-垂体-性腺轴中关键的信号分子,下丘脑根据光照变化等外界环境的刺激调节促卵泡素(Follicle-stimulating Hormone,FSH)、黄体生成素(Luteinizing Hormone,LH)、睾酮(Testosterone,T)或雌二醇(Estrogen,E2)等重要激素分泌[7-8]。研究发现,GnRH 广泛分布于各个物种的生殖、神经、内分泌、免疫系统和消化系统,它可以传递信息,从而协调整个系统[9-10]。GnRH 浓度和基因表达量随着繁殖周期的变化而变化[11]。GnRH基因与哺乳动物的繁殖、生长及猪和羊的体组成和生长性状相关[12-14];此外,GnRH基因SNP 位点与禽类的产蛋量、蛋品质等性状显著或极显著相关[15-16],与鹅的产蛋数、蛋重等性状显著或极显著相关[17-18]。
本实验室前期研究发现,鹅GnRH基因包含完整的4 个外显子和3 个内含子区域,92 个氨基酸序列,检测到46 个SNP 位点,并在四川白鹅群体(群体大小为217只)中发现GnRH基因是鹅产蛋性状的重要候选基因[17]。本研究以另外一个四川白鹅群体为实验材料,利用直接测序的方法对该基因的46 个SNP 位点进行基因分型,并进行单位点和单倍型与鹅的体组成和蛋品质性状的关联分析,鉴定GnRH基因显著影响鹅的体组成和蛋品质的分子标记,为鹅的分子选择育种提供分子标记。
1 材料与方法
1.1 实验动物和数据采集 本研究以重庆市荣昌区安富水禽实验基地的208 只四川白鹅(母鹅)群体为实验材料。为准确记录鹅蛋品质性状,每只鹅从出雏后进行系统的脚号和翅号标记,20 周龄时利用含有肝素钠的真空采血管翅静脉采血2 mL,利用血液基因组提取试剂盒(北京天根公司,DP332)提取血液基因组DNA,并将其溶解在TE 溶液中,在-20℃保存备用。上述四川白鹅孵化出壳后称量记录鹅初生重,饲养至28 周龄,每个个体转移到个体笼(600 mm×800 mm×900 mm)中饲养至65 周(休产期),并自由饮水和饲喂标准饲料。根据组成性状的测量方法[19],利用软尺测量成年鹅的颈长、胸宽、半潜水长、骨盆宽、体斜长、胫长、龙骨长和胫围等;在鹅产蛋高峰期(40 周龄),采集208 只鹅连续产的3 个鹅蛋,利用电子天平称量蛋壳重、蛋黄重;根据《家禽生产性能名词术语和度量统计方法》(NY/T 823-2004)测定,采用NaCl 溶液漂浮法测定蛋比重,设置9 个梯度的浓度(1.068、1.072、1.076、1.086、1.092、1.096、1.100 g/cm3);利用游标卡尺测量蛋壳的钝部、尖部和中间部位,并取三者的平均值计算蛋壳厚度;利用游标卡尺检测蛋长直径和短径,并计算蛋形指数;利用罗氏比色卡进行蛋黄颜色的测定;蛋壳强度利用蛋壳测力计(EFG-0502,Robotmation 公司)进行检测。
1.2 SNP 多态性位点检测和分型 为进一步研究GnRH基因与鹅其他性状的相关性,本研究以四川白鹅的基因组DNA 为模板,利用前期研究中的PCR 引物[17]和直接测序法对GnRH基因的SNP 多态性位点进行检测和基因分型。PCR 扩增体系50 μL:基因组DNA(<1 μg)1 μL,上、下游引物(10 μmol/L)各1 μL,2×Gflex PCR Вuffer 25 μL,Tks Gflex DNA 聚 合 酶1 μL,双 蒸水21 μL。PCR 扩增程序:98℃预变性2 min,30 个循环(98℃变性30 s,退火30 s,72℃延伸60 s),72℃延伸10 min。PCR 产物鉴定正确后送Ιnvitrogen 公司测序,用DNAStar 软件对序列进行比对分析。通过与前期的GnRH基因序列比对确定SNP 基因型[17]。参照命名系统(www.hgvs.org/mutnomen)对多态位点进行统一命名。
1.3 统计分析 针对上述GnRH基因的46 个SNP 位点,用JMP14.0 软件(SAS institute Japan,Minato-ku,Japan)通过滑动窗口方法构建单倍型,每3 个SNPs 构成1 个窗口,每次向前滑动1 个SNP。利用多因素方差分析模型分析GnRH基因SNP 多态性位点及其构建的单倍型与性状之间的关联性,模型:Y=μ+Gi+eij,其中Y 为性状观测值,μ 是群体平均值,Gi为个体基因型,eij为随机误差。计算各个性状对应的基因型的最小二乘均数。显著差异水平为P<0.05,极显著差异水平为P<0.01。
2 结果
2.1GnRH基因SNP 多态性位点与体组成性状的关联分析 SNP 多态性位点与体组成性状的相关分析结果(表1)表明,开产体重与g.1507C>T 和g.3099C>T 多态性位点显著相关,体斜长与g.408C>G、g.695T>C、g.1455A>G、g.1507C>T、g.1542A>G、g.1695A>G、g.1795C>T 位点显著或极显著相关;胫长与g.356_352 del 5、g.742G>C、g.793G>A、g.1542A>G、g.2476T>C、g.2510T>C、g.2542T>C 和g.3113A>C 显著或极显著相关;龙骨长与g.356_352 del 5、g.1307G>A、g.1311A>C、g.1695A>G 和g.1715C>A 显著或极显著相关;胫围与g.687A>G、g.1307G>A、g.1311A>C 和g.1836T>C 位点显著或极显著相关(P<0.05 或P<0.01);GnRH基因SNP 位点与半潜水长、骨盆宽和胫围等性状无显著相关。
2.2GnRH基因SNP 多态性位点与鹅蛋品质性状的关联分析 由表2 可见,GnRH基因的9 个SNP 位点(g.218G>A、g.350G>A、g.490A>G、g.1366A>C、g.1832A>G、g.2085C>T、g.2908C>T、g.2961C>G 和g.3113A>C)与蛋壳强度显著或极显著相关(P<0.05 或P<0.01);g.695T>C、g.1143T>C 和g.1246T>C 位 点 与 蛋 壳 重显著或极显著相关(P<0.05 或P<0.01)。GnRH基因SNP 位点与鹅蛋形指数、蛋比重、蛋黄颜色和蛋黄重等蛋品质性状无显著相关。
2.3 单倍型与体组成性状的关联分析 由表3 可知,window1、window4、window7、window10、window12、window19、window20、window23、window29、window32、window37、window38、window43 和window44,共14 个单倍型与初生重、体斜长、胫长、龙骨长和胫围等性状呈显著或极显著相关(P<0.05 或P<0.01)。这些单倍型包括单位点显著的21 个SNP 位点(表1),说明这21 个SNP 可能是影响鹅的初生体重、体斜长、胫长、龙骨长和胫围性状的重要变异位点。
2.4 单倍型与蛋品质性状的关联分析 由表3 可知,window1、window6、window7、window9、window10、window12、window16、window43 和window44,共9个单倍型与蛋壳强度、蛋壳重和蛋壳厚度等性状呈显著或极显著相关(P<0.05 或P<0.01)。这些单倍型包括单位点显著的19 个SNP 位点(表2),表明这19 个SNP 位点可能是影响鹅的蛋壳强度、蛋壳重和蛋壳厚度等性状的重要变异位点。
3 讨论
前期研究结果表明,GnRH基因在畜禽繁殖和体组成性状等方面发挥重要功能[17,20-23]。因此本研究将GnRH基因的46 个SNP 位点与鹅的体组成和产蛋性状的相关性进行分析,为进一步探究该基因影响上述性状的遗传机理提供依据并提供相应的潜在分子标记。
人和哺乳动物中研究表明,GnRH基因调控GnRH激素合成,影响与体组成相关的性腺分泌的激素,而且GnRH基因与青少年的体组成成分显著相关[24];哺乳动物研究中发现,去势猪的体组成性状和血液中的GnRH激素浓度与正常猪差异显著[13];与此类似,去势羊血液体组成性状和血液中的GnRH 激素浓度与正常组差异显著[14],推断GnRH 激素浓度影响猪体组成性状。综上所述,本研究推测GnRH 的SNP 多态性位点,影响GnRH 分泌,直接或者间接影响鹅的体组成性状,具体机制需要进一步验证。
本研究发现,GnRH基因SNP 位点与四川白鹅蛋壳强度、蛋壳重和蛋壳厚度等指标有显著或极显著的影响。值得注意的是,g.1366A<C 突变位点造成谷氨酸到丙氨酸的突变,与蛋壳强度和蛋壳厚度显著相关,AA 基因型个体的蛋壳强度均大于其他基因型个体。前期研究发现g.1366A<C 突变位点与四川白鹅产蛋数显著相关,且AA 基因型个体产蛋数显著低于CC 基因型个体[17]。研究发现褐壳纯系蛋鸡的蛋壳厚度与产蛋数负相关[25],一定程度上解释了g.1366A<C 突变位点AA 基因型的蛋壳厚度高于其他基因型,而产蛋数显著低于CC 基因型。研究表明鹅的GnRH基因SNP 位点与蛋重、产蛋数等显著相关[17,21],但对本研究中的4 个SNP 位点(g.1795C>T、g.1822A>G、g.2760T>C 和g.3348C>T)均未报道,可能是由于鹅品种、群体大小或者关联分析的公式等不同造成结果差异。
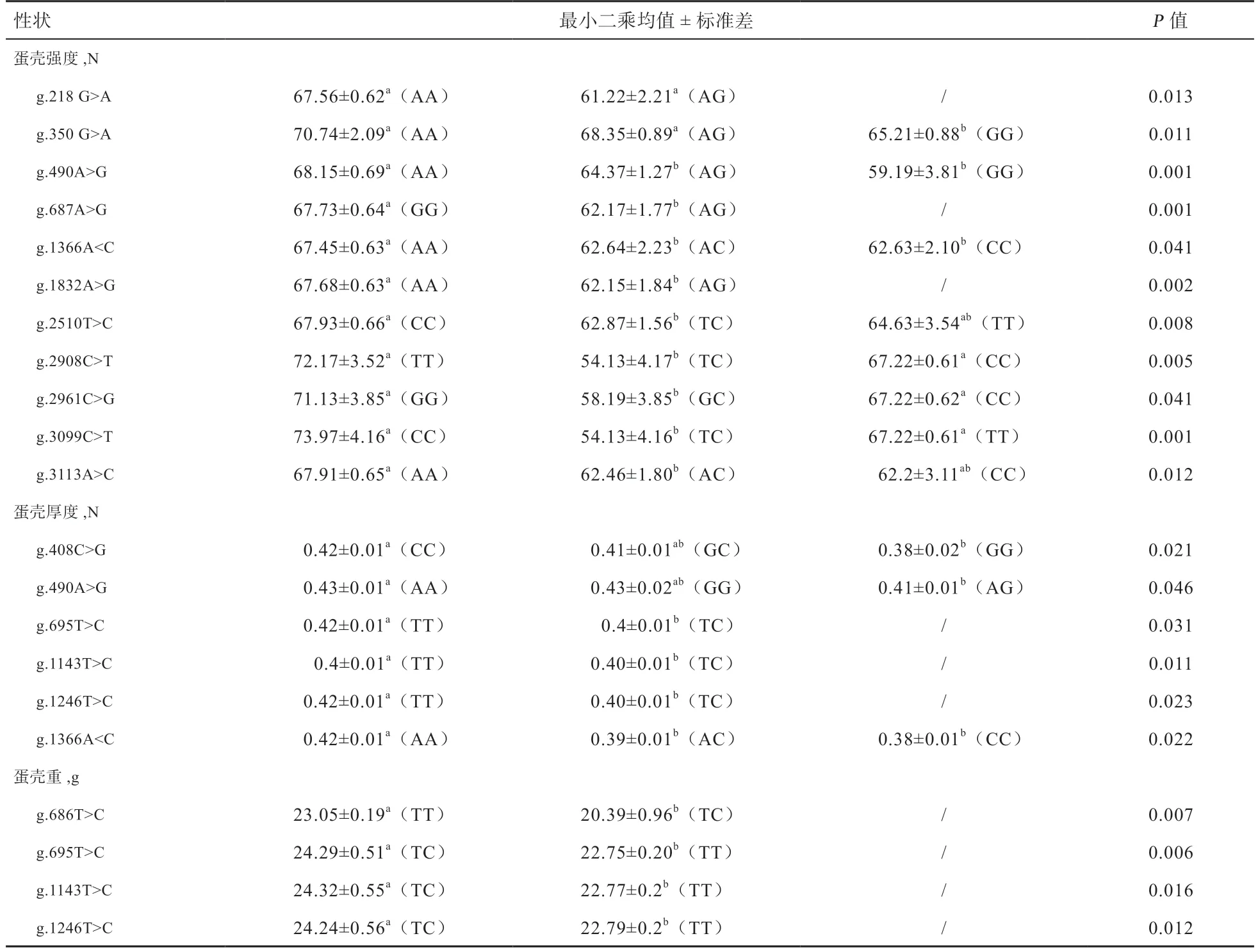
表2 GnRH 不同基因型对鹅蛋品质性状的影响
蛋壳强度、蛋壳硬度和蛋壳重等指标与钙离子吸收相关。本研究发现,GnRH基因的21 个SNP 位点与蛋壳强度、蛋壳硬度和蛋壳重显著或极显著相关。GnRH的电生理特性及活性神经元高度依赖于钠、钾、钙和氯等离子通道活性及离子的浓度[26-27]。钾、钙、氯通道对静息状态的激活及GnRH 神经元的激活,某些离子通道基因和特定离子的表达,离子状态浓度状态影响鹅蛋重量、蛋壳厚度[28]。推测GnRH与上述蛋品质相关的SNP 位点的不同基因型的电生理特性不同,对应的离子通道和离子浓度不同,因此蛋品质在不同基因型也存在显著差异,具体机制需要进一步实验证明。
4 结论
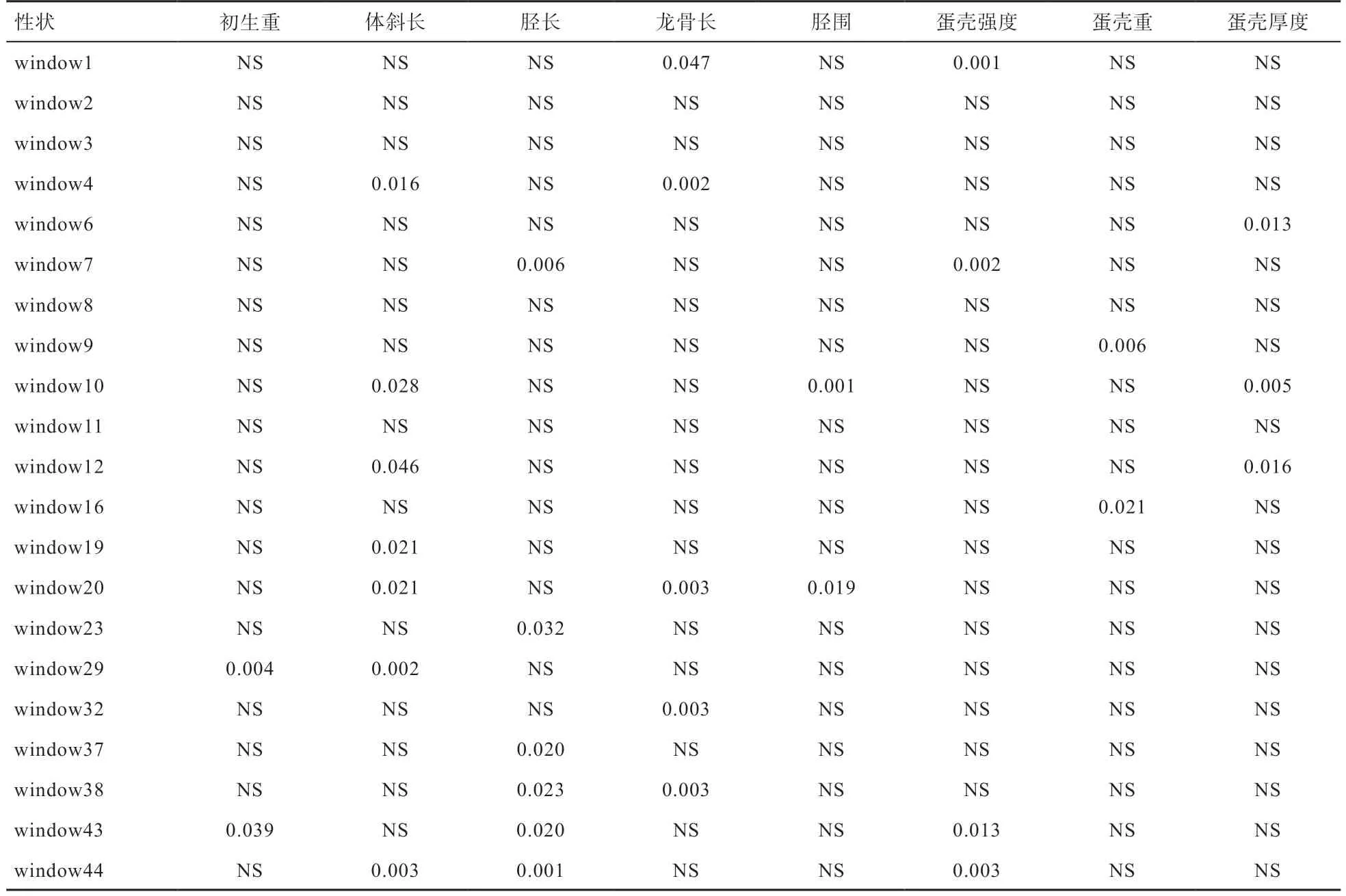
表3 GnRH基因单倍型对鹅体组成和蛋品质的影响(P值)
本实验结果表明,GnRH基因的多态性位点与鹅初生重、体斜长、胫长、龙骨长、胫围、蛋壳强度、蛋壳重和蛋壳厚度等性状显著或极显著相关,是上述体组成和蛋品质性状的潜在分子标记,有望应用于鹅育种实践并有助于上述性状遗传机理的研究。
