极濒危物种天台鹅耳枥群落特征及其与环境因子的关系
2020-10-17陈模舜金则新柯世省陈子林潘德月
陈模舜 金则新 柯世省 陈子林 潘德月
(1. 台州学院生命科学学院 浙江省植物进化生态学与保护重点实验室 台州 318000; 2. 浙江省大盘山国家级自然保护区管理局 磐安 322300)
全球环境变化及人类活动对生物群落的影响日益加剧,种群栖息地遭到破坏、生境破碎化及生物入侵等已对植物尤其是珍稀濒危植物产生重要影响(Wilcoxetal., 1985; Loweetal., 2005)。生境片断化导致濒危植物以小种群的形式存在,从而容易遭受遗传侵蚀和近交进而影响其生存(Neietal., 1975; Fingeretal., 2012)。极濒危物种天台鹅耳枥(Carpinustientaiensis)属于桦木科(Betulaceae)鹅耳枥属落叶乔木,其分布区很狭窄,《中国植物志》(中国科学院中国植物志编辑委员会, 1979)曾记载仅浙江省天台县华顶山有分布,《浙江植物志》(王景祥, 1992)记载仅存5株。但近年来在磐安县等地也有发现,是第一批被列入国家二级重点保护的野生植物(汪松等, 2004)。由于受自然干扰和旅游开发、森林采伐等人为活动影响,长期处于生境片断化状态的天台鹅耳枥群落分布区呈进一步减小趋势,已处于极濒危状态。天台鹅耳枥是分类学上地位特殊的物种,在研究植物地理、植物区系等方面均有较高科研价值,对森林群落生物多样性的保持具有重要作用。近年来国内外采用双向指示种分类法(TWINSPAN)和典范对应分析(CCA)研究植物群落和种群特征,分析群落类型及其与环境的关系(许芸等, 2007; 代力民等, 2008; Zereenetal., 2014; Moradietal., 2018)。目前对天台鹅耳枥自然环境群落分类、CCA排序的相关研究未见报道,仅在天台鹅耳枥营养器官解剖结构、光合特性对光强的响应、不同光环境下的叶形变化、组织培养与快速繁殖等方面进行了研究(陈珍等, 2013; 陈模舜等, 2018)。为了更有效地保护天台鹅耳枥,本研究全面调查了天台鹅耳枥群落物种区系特征、种群变化趋势和栖息地环境,运用TWINSPAN 分类和CCA 排序方法研究浙江省天台县和磐安县的天台鹅耳枥天然植物群落特征及其与环境的相关性,以期为科学有效地恢复天台鹅耳枥种群提供依据(Hanski, 1998)。
1 研究区概况
天台鹅耳枥自然群落分布于浙江省天台县和磐安县的4个相互隔离的地方。天台县地处120°41′—121°15′E,28°57′—29°20′N,天台山脉系武夷山仙霞岭中支,由南向北延伸而来,主要为花岗岩侵蚀体构成,主峰华顶山(121°05′E,29°15′N)海拔1 098 m,四季温暖湿润,年均降水量1 700 mm,年均相对空气湿度达85%以上,年均气温17 ℃,≥10 ℃年积温2 858~5 157 ℃,全年无霜期长达234天。华顶山有7株天台鹅耳枥大树,10株中树,幼苗和小树共13株,分布于海拔850~1 000 m的低山山坡疏林中,树高9.5~19 m,生境面积约1.5 km2。
磐安县(120°17′—120°47′E,28°49′—29°19′N)共存天台鹅耳枥遗株5株,其中大盘山3株,高姥山和青明尖各1株,分布于海拔1 000~1 250 m的疏林中,树高5.6~9 m,群落之间相隔20 km之内。磐安县大盘山是天台山、仙霞岭和括苍山的承接处,主峰盘山尖(120°31′E,28°58′N)海拔1 245 m,年均气温16.1 ℃,全年无霜期236天,年均降水量1 573 mm。
天台县华顶山和磐安县两地相距85 km,最高海拔相近,均在1 000 m以上,同属亚热带季风气候,土壤均为红壤和黄壤,呈酸性,森林植被覆盖率较高,植物年生长期长,地带性植被类型为亚热带常绿阔叶林(洪仲棉, 1988; 陈远志等, 2011)。
2 研究方法
2.1 天台鹅耳枥群落学调查
基于多年对天台鹅耳枥野外群体与生境的全面调查,在天台县华顶山和磐安县4个天台鹅耳枥分布区的天然林设置固定样地进行定位观察。以天台鹅耳枥分布点为中心设置样地,取样地点为天台县华顶山,磬安县的大盘山、高姥山和青明尖,共设置8块样地,每块样地大小20 m×20 m,调查各样地中有天台鹅耳枥分布的24个10 m×10 m样方,记录每个样方内乔木的株数、胸径、盖度和高度; 在样方的东南、西北两角部位分别设置1个5 m×5 m的灌木小样方; 在样方的东南、西北两角及中心部位分别设置1个1 m×1 m的草本小样方,调查灌木和草本植物的种类、株(丛)数、盖度和高度; 共调查24个乔木样方,48个灌木样方,72个草本样方。同时记录样方所在地的海拔、坡向、坡度、土壤质地和干扰等生态因子。样地概况见表1。
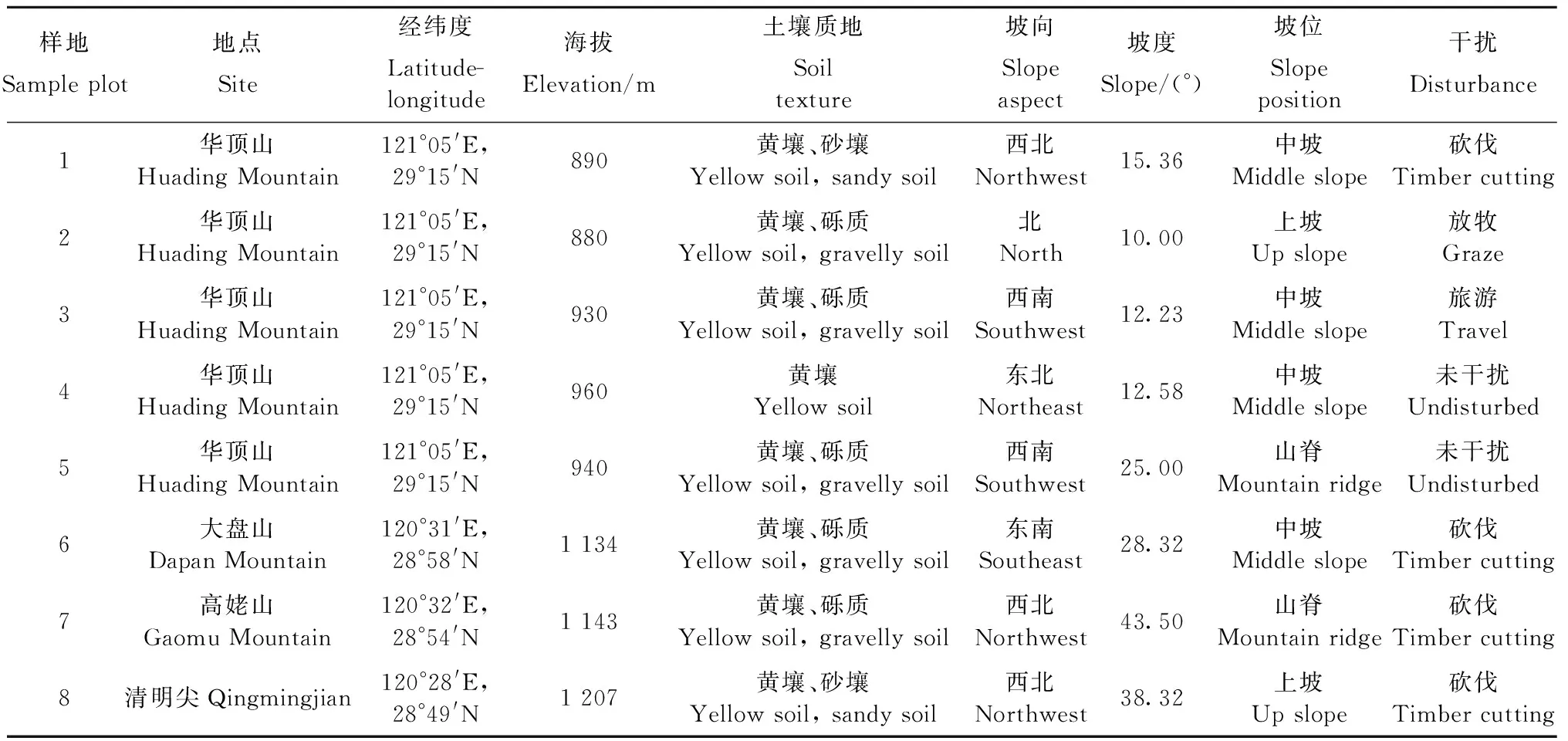
表1 天台鹅耳枥群落样地概况Tab.1 Survey of sample plots of C. tientaiensis community
用多功能坡度测量仪JZC-B2测量坡度、用地质罗盘仪DQY-1测量坡向、用Garmin Monterra GPS测量海拔、经纬度。土壤质地依据中国土壤质地分类系统(吕贻忠, 2008)确定。
2.2 物种重要值计算
物种重要值在一定程度上反映某一物种在群落中的作用和价值,常用于比较某一物种在不同群落中的重要性。依据调查数据,选用下列公式计算各样方内植物种类的重要值:
IV1=(RD + RP + RF)3;
(1)
IV2=(RD + RC + RF)3;
(2)
(3)
(4)
(5)
(6)
式中: IV1为乔木树种的重要值; IV2为灌木或草本的重要值; RD 为相对密度; RP 为相对显著度; RF为相对频度; RC 为相对盖度;Di为i物种的株数与所有种的总株数之比;Pi为i物种胸径断面积和与所有种个体的胸径断面积和之比;Fi为i物种的频度与全部物种的频度之比;Ci为i物种所有个体的盖度和与所有种个体的盖度面积和之比(方全等, 2015)。
2.3 植物群落分析
植被的数量分类以PC-ORD5.0中的TWINSPAN分类双向指示种分析来完成,排序用CANOCO for Windows 5.0软件完成(Peck, 2010;milaueretal., 2014)。其中CCA分析法是对研究区植被数据矩阵和环境数据矩阵进行排序,研究群落分布格局与环境因子的关系。在天台鹅耳枥种群野外收集数据的基础上,原始资料由Excel整理成原始矩阵,对调查样地植物的重要值进行群落分析,记录植物48科74属92种,剔除样方中重要值小于1.50的稀有种,群落主要的维管植物有46种,从而形成24×46(24个样方)的重要值矩阵及24×6(6个环境因子)的环境数据矩阵。环境数据包括海拔、坡向、坡度、坡位、土壤质地和干扰6个因子,其中海拔以高度值表示; 坡位分为斜坡(0°~15°)、陡坡(15°~25°)和峭坡(35°~55°)3个水平; 坡向赋值方法为东南坡和西南坡取3分、东北坡和西北坡取2分、北坡取1分; 坡位的赋值方法为山脊取最高分4分、上坡位取3分、中坡位取2分、中下坡位取1分,分值越大,接受的光照、热量越多; 土壤质地赋值方法为黄壤、砾质取最高分3分,黄壤、砂壤取2分,黄壤取1分; 干扰赋值方法为未干扰取最高分4分,旅游取3分,放牧取2分,砍伐取1分(邱扬等, 2000)。
2.4 年龄结构分析
年龄结构是种群的重要特征,对乔木种群,许多学者用径级结构代替年龄结构研究树种的结构和动态特征。野外样方调查时对天台鹅耳枥植株进行每木检尺,记录树高、胸径等,将天台鹅耳枥的龄级结构按以下标准分为7级: Ⅰ级幼苗,树高<0.3 m; Ⅱ级幼树,树高0.3~1.2 m,胸径(DBH)<2.5 cm; Ⅲ级小树,树高> 1.2 m,DBH为2.5~5 cm; Ⅳ级中树,DBH为5~10 cm; V级大树,DBH为10~25 cm; Ⅵ级大树,DBH为25~45 cm; Ⅶ级大树,DBH为45~60 cm(陈子林等, 2007)。
3 结果与分析
3.1 天台鹅耳枥群落植物属的地理分布
天台鹅耳枥群落主要的维管植物有46种,隶属29科40属。参照吴征镒等(2011)关于中国种子植物属的分布区类型的划分,对组成天台鹅耳枥群落的种子植物区系地理成分进行分析(表2),结果表明,天台鹅耳枥群落中种子植物属的地理分布类型有10种,隶属于37个属中。其中世界分布6属,有鼠李属(Rhamnus)、鬼针草属(Bidens)、珍珠菜属(Lysimachia)、毛茛属(Ranunculus)、堇菜属(Viola)和苔草属(Carex); 北温带分布型以及变型所占比例最大,共有11属,占属总数的45.48%,有鹅耳枥属、李属(Prunus)、槭树属(Acer)、栎属(Quercus)、忍冬属(Lonicera)、杜鹃花属(Rhododendron)、紫菀属(Aster)、风轮菜属(Clinopodium)、玄参属(Scrophularia)、委陵菜属(Potentilla)和龙牙草属(Agrimonia); 东亚和北美洲间断分布有6属,占属总数的19.35%,有柳杉属(Cryptomeria)、栗属(Castanea)、石楠属(Photinia)、金线草属(Antenoron)、獐耳细辛属(Hepatica)和绣球属(Hydrangea); 东亚分布5属,占属总数的16.13%,有松属(Pinus)、蕺菜属(Houttuynia)、博落回属(Macleaya)、野鸦椿属(Euscaphis)和兔儿伞属(Syneilesis); 泛热带分布2属,占属总数的6.45%,有山矾属(Symplocos)和菝葜属(Smilax); 热带亚洲和热带美洲间断分布1属,占属总数的3.23%,为泡花树属(Meliosma); 热带亚洲和热带大洋洲分布1属,占属总数的3.23%,为淡竹叶属(Lophatherum); 热带亚洲至热带非洲分布1属,占属总数的3.23%,为类芦属(Neyraudia); 热带亚洲分布3属,占属总数的9.68%,为山茶属(Camellia)、山胡椒属(Lindera)和箬竹属(Indocalamus); 中国特有分布1属,占属总数的3.23%,为杉木属(Cunninghamia)。在天台鹅耳枥群落中,有许多第三纪或更古老的残遗属,如松属、杉木属、鹅耳枥属、菝葜属、栎属和槭属等。这些区系成分的存在,说明天台鹅耳枥群落的古老性和残遗性。天台鹅耳枥群落植物区系属于泛北极区,属的分布类型以北温带分布、东亚和北美洲间断、世界分布和东亚分布类型为主,其次是热带亚洲、泛热带分布和热带美洲间断分布; 天台山植物区系的暖温带特征显著,热带、亚热带成分也有相当大的比重。

表2 天台鹅耳枥群落中植物属的地理分布类型统计①Tab.2 Geographical distribution types of plant genera in C. tientaiensis community
3.2 天台鹅耳枥种群结构
物种组成是群落最基本的特征。在调查的24个样方中,主要的46种植物属于29科40属,其中蕨类植物3科3属3种,种子植物26科37属43种。乔木共11种,灌木共13种,草本共22种。乔、灌、草层植物重要值见表3—5。
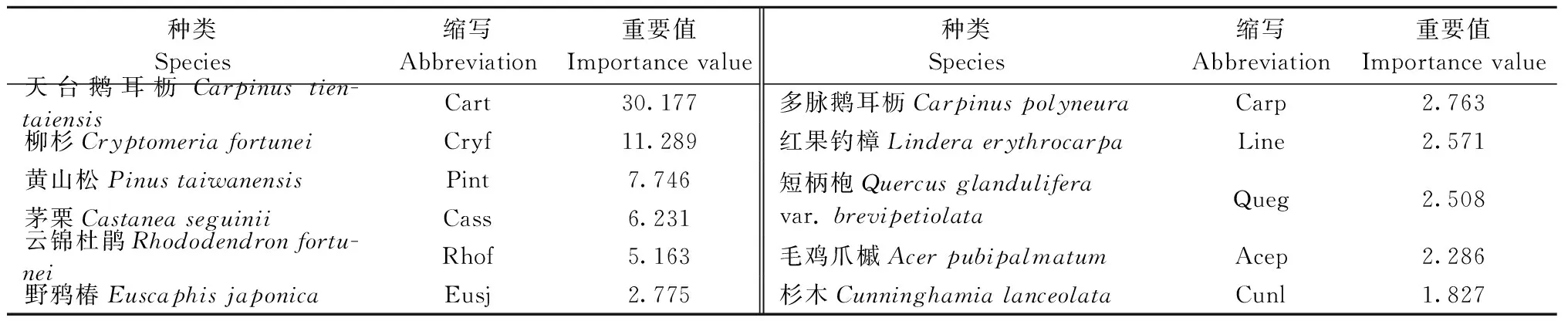
表3 天台鹅耳枥群落乔木层主要物种重要值①Tab.3 Importance value of the main species in tree layer of C. tientaiensis community
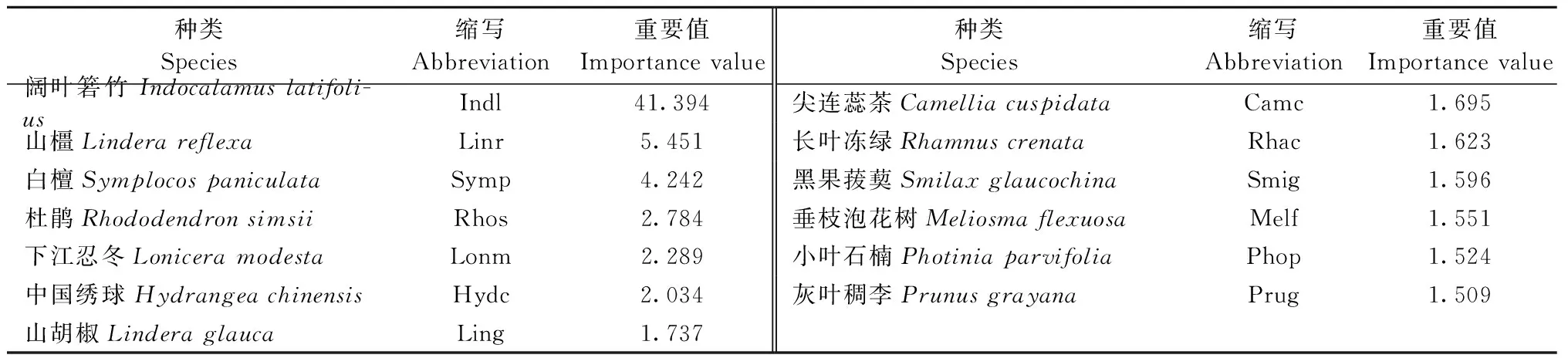
表4 天台鹅耳枥群落灌木层主要物种重要值Tab.4 Importance value of the main species in shrub layer of C. tientaiensis community

表5 天台鹅耳枥群落草本层主要物种重要值Tab.5 Importance value of the main species in herb layer of C. tientaiensis community
3.3 天台鹅耳枥群落类型与分布
利用物种重要值对天台鹅耳枥群落24个样方以及主要的46种植物进行TWINSPAN聚类,将天台鹅耳枥的群落类型归纳为 7个群丛(图1)。其中乔木层以天台鹅耳枥重要值最大,灌木层以阔叶箬竹重要值最大。群落中禾本科(Gramineae)和蔷薇科(Rosaceae)物种丰富度最高,各为4种,其次为菊科(Compositae)3种、壳斗科(Fagaceae)2种。24个样方中,出现次数较高的伴生植物分别为阔叶箬竹(17/24)、山橿(9/24)、白檀(9/24)、茅栗(8/24)和柳杉(7/24)等。根据TWINSPAN 物种的聚类结果,46种植物被分为2类:第一类有1种,为阔叶箬竹; 第二类有45种,分6个亚类,其中亚类又分为13个基本类(图1)。综合样方和物种的聚类结果,结合天台鹅耳枥为优势种的群落调查结果,同时对TWINSPAN分类时可能出现基本类的错分类现象,进行必要的人为调整(张金屯, 1995),依据植被的群落命名原则,天台鹅耳枥群落类型分为Ⅰ—Ⅶ类群丛。
Ⅰ.天台鹅耳枥-云锦杜鹃-多脉鹅耳枥+阔叶箬竹-黑果菝葜+三脉紫菀群丛 包括样方1、3、4、7、11、12、14、17、18和24,海拔为880~960和1 207 m。乔木层优势种为天台鹅耳枥,主要伴生种为云锦杜鹃、多脉鹅耳枥、茅栗、黄山松、野鸦椿、柳杉和毛鸡爪槭等; 灌木层优势种为阔叶箬竹,主要伴生种为黑果菝葜、小叶石楠、下江忍冬、白檀和灰叶稠李; 草本层优势种为三脉紫菀,主要伴生种为玄参、堇菜等。
Ⅱ.天台鹅耳枥+中国绣球-长叶冻绿+山类芦-南方兔儿伞群丛 包括样方20、21和22,海拔为1 134 m。乔木层优势种为天台鹅耳枥,主要伴生种为茅栗、杉木、红果钓樟和黄山松; 灌木层优势种为中国绣球,主要伴生种为长叶冻绿、白檀、山橿和杜鹃; 草本层优势种为山类芦,主要伴生种为南方兔儿伞、大披针苔草、淡竹叶、渐尖毛蕨、獐耳细辛和星宿菜等。
Ⅲ.柳杉-天台鹅耳枥-杉木+阔叶箬竹群丛 包括样方9、13、15和19,海拔930~960 m。乔木层优势种为柳杉和天台鹅耳枥,主要伴生种为杉木、茅栗、多脉鹅耳枥和短柄枹; 灌木层优势种为阔叶箬竹,主要伴生种为长叶冻绿、白檀、小叶石楠、灰叶稠李、山橿和黑果菝葜等。
Ⅳ.黄山松-天台鹅耳枥-短柄枹+阔叶箬竹-山橿群丛 包括样方2,海拔890 m。乔木层优势种为黄山松和天台鹅耳枥,主要伴生种为短柄枹; 灌木层优势种为阔叶箬竹,主要伴生种为山橿。
Ⅴ.天台鹅耳枥+三叶委陵菜-蕺菜-细风轮菜群丛 仅样方6,海拔930 m。乔木层优势种为天台鹅耳枥; 草本层优势种为三叶委陵菜,主要伴生种为蕺菜、细风轮菜、毛茛、珍珠菜、鬼针草、博落回、堇菜和龙牙草。
Ⅵ.天台鹅耳枥+阔叶箬竹-下江忍冬+蕺菜-玄参-巴东过路黄群丛 包括样方8、10、16和23,海拔为930~960和1 143 m。乔木层优势种为天台鹅耳枥,主要伴生种为云锦杜鹃、多脉鹅耳枥和茅栗; 灌木层优势种为下江忍冬,主要伴生种为尖连蕊茶、垂枝泡花树、山胡椒、白檀和杜鹃; 草本层优势种为蕺菜,主要伴生种为玄参、巴东过路黄、三叶委陵菜、堇菜和三脉紫菀等。
Ⅶ.天台鹅耳枥-红果钓樟-毛鸡爪槭+阔叶箬竹+金线草群丛 仅包括样方5,海拔880 m。乔木层优势种为天台鹅耳枥,主要伴生种为红果钓樟和毛鸡爪槭; 灌木层优势种为阔叶箬竹,主要伴生种为下江忍冬、白檀和山橿; 草本层优势种为金线草,主要伴生种为玄参等。

图1 天台鹅耳枥样方与主要物种的双向聚类分析Fig.1 Two-way cluster dendrogram of C. tientaiensis quadrats and main species阴影方格矩阵表示24个样方×46个物种矩阵,树形图表示聚类 The matrix of shaded squares represents 24 quadrats × 46 species matrix, while the dendrograms show the clusteringQ1-Q24:样方序号Quadrate number;T1:天台鹅耳枥Carpinus tientaiensis;T2:柳杉Cryptomeria fortunei;T3:黄山松Pinus taiwanensis;T4:茅栗Castanea seguinii;T5:云锦杜鹃Rhododendron fortunei;T6:野鸦椿Euscaphis japonica;T7:多脉鹅耳枥Carpinus polyneura;T8:红果钓樟Lindera erythrocarpa;T9:短柄枹Quercus glandulifera var. brevipetiolata;T10:毛鸡爪槭Acer pubipalmatum;T11:杉木Cunninghamia lanceolata;S1:阔叶箬竹Indocalamus latifolius;S2:山橿Lindera reflexa;S3:白檀Symplocos paniculata;S4:杜鹃Rhododendron simsii;S5:下江忍冬Lonicera modesta;S6:中国S7绣球Hydrangea chinensis;S8:山胡椒Lindera glauca;S9:尖连蕊茶Camellia cuspidata;S10:长叶冻绿Rhamnus crenata;S11:黑果菝葜Smilax glaucochina;S12:垂枝泡花树Meliosma flexuosa;S13:小叶石楠Photinia parvifolia;S14:灰叶稠李Prunus grayana;H1:蕺菜Houttuynia cordata;H2:金线草Antenoron filiforme;H3:三叶委陵菜Potentilla freyniana;H4:玄参Scrophularia ningpoensis;H5:细风轮菜Clinopodium gracile;H6:堇菜Viola verecunda;H7:巴东过路黄Lysimachia patungensis;H8:渐尖毛蕨Cyclosorus acuminatus;H9:蕨Pteridium aquilinum var. latiusculum;H10:南方兔儿伞Syneilesis australis;H11:三脉紫菀Aster ageratoides;H12:淡竹叶Lophatherum gracile;H13:大披针苔草Carex lanceolata;H14:珍珠菜Lysimachia clethroides;H15:毛茛Ranunculus japonicus;H16:紫萁Osmunda japonica;H17:博落回Macleaya cordata;H18:獐耳细辛Hepatica nobilis var. asiatica;H19:鬼针草Bidens pilosa;H20:龙牙草Agrimonia pilosa;H21:星宿菜Lysimachia fortunei;H22:山类芦Neyraudia montana.
由于生境片段化分布区狭小,群丛Ⅳ、Ⅴ和Ⅶ仅由单个样方组成,群丛Ⅴ没有灌木层,仅乔木层和草本层,群落组成简单。
3.4 天台鹅耳枥群落与环境因子的CCA 排序
在进行物种-环境排序之前,首先利用DCA检测物种矩阵的排序轴梯度,结果显示排序轴最大值为5.63,超过4,故可选用典范对应分析(CCA)排序探讨物种-环境之间的关系(表6)(milaueretal., 2011)。
运用CCA对应分析自然和干扰因素对天台鹅耳枥群落物种分布的影响,物种的排序与环境信息表示在双序图(biplot)上(图2)。基于排序计算结果(表6),第1排序轴包含了28.30%的生态信息,前2排序轴累计包含了51.07%的生态信息,前3排序轴累计包含了72.66%的生态信息。排序轴显著性检验结果显示CCA第1轴的显著性为P=0.006,模型整体显著性为P=0.016,表明该CCA模型的排序效果及可信度均较高。

表6 天台鹅耳枥群落物种分布与环境因子的CCA排序计算结果Tab.6 CCA calculation of species distributions and environmental factors in C. tientaiensis community
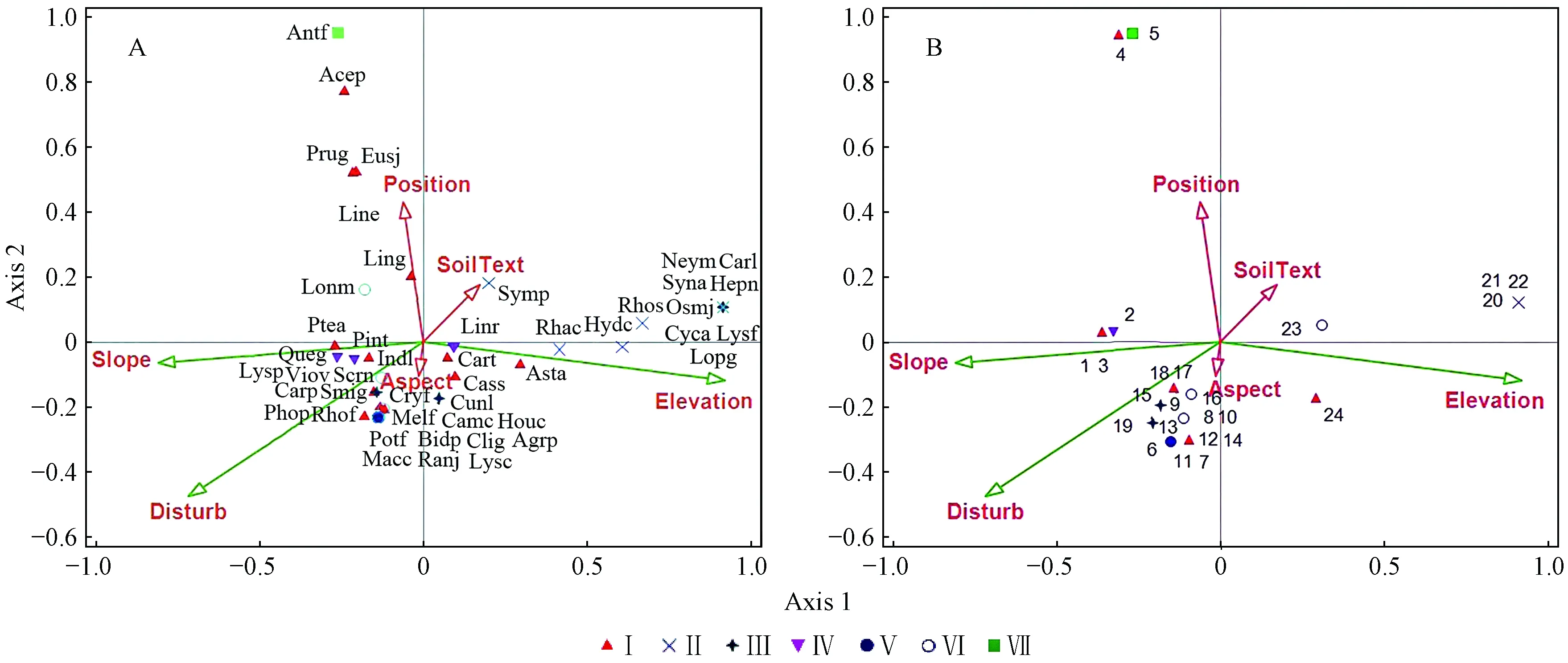
图2 主要物种(A)、样方(B)和环境因子CCA 排序Fig.2 CCA biplot of main species and quadrats-environment factors物种缩写为属名前3个字母加种加词的第1个字母(如表3-5)The species are abbreviated with the first three letters of their genus name and the first letters of species name (as the table 3-5 showed).1-24: 样方编号Quadrate codes; Ⅰ-Ⅶ: 群丛类型Association types; Elevation: 海拔高度; SoilText: 土壤质地Soil texture; Aspect: 坡向Slope aspect; Slope: 坡度Slope; Position: 坡位Slope position; Disturb: 干扰Disturbance.
运用变量分解方法,Monte Carlo置换检验,以海拔、坡度和干扰为主变量,土壤质地、坡向和坡位为协同变量,海拔、坡度和干扰与CCA排序轴显著相关(P<0.01),而土壤质地、坡向和坡位与CCA排序轴均无显著相关性(表7)。与第一轴相关性系数最高的环境因子是海拔,呈正相关(相关系数为0.905 6,P<0.01) 与干扰呈负相关(相关系数为-0.705 9,P<0.01)。第二轴与坡位的相关系数最高(0.625 6), 与坡度呈负相关(相关系数为-0.898 1,P<0.01)。由图2可知:第一轴反映了植物群落所在生境的海拔和干扰情况,即沿着CCA 第一轴从左往右,海拔逐渐升高,光照条件逐渐变好,海拔和干扰为主要影响因子; 第二轴基本上表现了植物群落所在生境坡位和坡度的变化趋势,即沿着CCA 第二轴从下至上,坡位从中坡到上坡,植物群落接近于山顶,坡度由平缓变陡峭,坡度为主要影响因子。根据显著性检验结果,海拔、干扰和坡度是对植物群落分布起决定作用的因子。

表7 限制性排序分析中单个解释变量的显著性检验①Tab.7 Significance test of individual interpretation variables in the restriction sorting analysis
3.5 天台鹅耳枥径级结构
种群径级结构反映种群内不同年龄个体的数量分布状况,是判断植物种群发展趋势的重要指标。根据天台鹅耳枥群落野外调查数据,以立木径级结构代替种群年龄结构,初步分析天台鹅耳枥种群动态。
图3表明,天台鹅耳枥种群的径级结构呈负增长型,以Ⅳ径级中龄树为主,缺少Ⅰ、Ⅱ和Ⅲ级的幼苗、幼树和小树,种群的年龄结构属于衰退类型。在实际调查中发现,仅天台县华顶山分布点存在幼龄级苗木。 通过对磐安县3个分布点的考察和当地调研,多年未见种苗萌发,可见天台鹅耳枥种群的年龄结构表现为衰退型,自我更新能力差。

图3 天台鹅耳枥群落的径级结构Fig.3 Diameter class structure of C. tientaiensis community
4 讨论
采用物种重要值进行TWINSPAN分类排序,以群落的优势种重组命名,将天台鹅耳枥群落样方划分为7个群丛类型,群落物种组成反映环境特征。通过物种、样方与环境CCA二维排序可知,影响天台鹅耳枥群落及物种分布格局的主要环境因子是海拔、土壤质地、地形要素和干扰等,其中海拔、干扰和坡度作用最大(杨国栋等, 2018)。
海拔决定天台鹅耳枥分布范围,群落集中分布于海拔850~1 250 m的中低山亚热带常绿阔叶林中,在整个群落中热带、亚热带成分突出。天台鹅耳枥与柳杉、黄山松、短柄枹、云锦杜鹃、多脉鹅耳枥和红果钓樟等强阳性树种形成混合群落,各群丛灌木层和草本层的组成由于地域的差异而有很大变化,灌木优势种为阔叶箬竹,也包括山橿、下江忍冬、中国绣球、山胡椒和长叶冻绿等落叶种类; 草本为蕺菜、金线草、玄参、细风轮菜、巴东过路黄、南方兔儿伞、三脉紫菀和大披针苔草等1年或多年生草本植物,大多为冬季地上部分枯死的种类。天台鹅耳枥群落优势种中不少是属于中亚热带的固有成分,表明了天台鹅耳枥群落有着与当地气候相适应的植被构成(陈灵芝, 2014)。
环境作用和物种退化是导致物种濒危的原因,生境的丧失和破碎化使植物物种失去赖以生存的栖息地,是物种受威胁的主要原因(蒋志刚等, 2014; 董鸣, 2017)。由于分布区域狭小、生境呈岛屿化,天台鹅耳枥部分群丛结构简单,群落稳定性不高(Saundersetal., 1991; Liuetal., 2014)。随着分布区显著缩小,有些群落天台鹅耳枥仅剩少量植株(Heinkenetal., 2013),如黄山松-天台鹅耳枥-短柄枹+阔叶箬竹-山橿群丛和天台鹅耳枥-红果钓樟-毛鸡爪槭+阔叶箬竹+金线草群丛等。天台鹅耳枥栖息地随着隔离时间的增加及人为干扰的加剧,片断化的群落结构变得不完整(吴倩倩等, 2017),如天台鹅耳枥+三叶委陵菜-蕺菜-细风轮菜群丛,没有灌木层,仅乔木层和草本层,群落组成简单,物种盖度较低。天台鹅耳枥生长在山坡疏林和溪谷两旁,对太阳辐射量和湿度要求较高。天台鹅耳枥+中国绣球-长叶冻绿+山类芦-南方兔儿伞群丛分布在坡度较大地带,由于重力作用以及地表径流,水分不断向下渗透,导致其表层土壤的水分条件低于坡度小的生境,出现干旱的生境,自然植被退化,物种组成相对简单(闫东锋等, 2010; 廉凯敏等, 2015)。天台县华顶山是最大的野生天台鹅耳枥群落,其群落为天台鹅耳枥-云锦杜鹃-多脉鹅耳枥+阔叶箬竹-黑果菝葜+三脉紫菀群丛,分布在坡度较缓地带,土壤湿度较好,有少量野生天台鹅耳枥幼树及幼苗成活,这可能是由于面积较大的栖息地内部环境较稳定,受到外界气候等不利因素的干扰较少(蒋志刚等, 2014; Lenoreetal., 2019)。
5 结论
天台山脉长期受相对稳定的亚热带气候影响,保留了较多的我国特有种,并保存了一定面积较为完好的森林植被。虽然天台鹅耳枥能够适应较多群落类型,但其所处的群落稳定性不高,容易受到自然和人为干扰影响。天台鹅耳枥群落分布受海拔、坡向、坡度、土壤质地和干扰等生态因子的影响,其中海拔、干扰和坡度是对植物群落分布起决定作用的因子,但土壤质地、坡向与坡位等要素在一定程度上也影响着天台鹅耳枥群落分布。在制定天台鹅耳枥多样性保护对策时,应该对那些面积较大、植物物种丰富度高的栖息地给予重点关注。生境片断化是濒危植物天台鹅耳枥减少的主要原因,林下幼苗少见,种群呈衰退趋势,有必要把人工繁殖苗木回归原生地保护或迁到人工形成的适宜生境,切实保护好原生地环境和群落伴生植物,扩大种群数量。由于生境的严重片断化,本研究涉及的天台鹅耳枥群落样方数量有限,未来相关研究要考虑其特有的生境及生物学和生理生态学特性,深入探索天台鹅耳枥种群天然更新困难的相关机制。
