GF-3全极化SAR数据极化分解估算人工林冠层生物量*
2020-10-17魏晶昱范文义毛学刚
魏晶昱 范文义 于 颖 毛学刚
(东北林业大学林学院 森林生态系统可持续经营教育部重点实验室 哈尔滨 150040)
森林生物量指单位面积森林群落在一定时间内积累的有机质总量(尹惠妍等, 2014),其可直接反映森林植被固碳现状,是评价森林生态系统结构、功能和生产力的重要指标之一,也是进行森林资源清查、规划和管理的依据。树冠是树木光合作用、呼吸作用、蒸腾作用的主要发生场所,影响树木生长和变化(沈浩等, 2017; Ozanneetal., 2003),冠层生物量是森林生物量研究的重要内容,其对环境变化十分敏感,在一定程度上影响着森林生态系统生产力(李凤日, 2004; 刘兆刚等, 2005; Wangetal., 1990; Kramer, 1996; Tahvanainenetal., 2008)。
传统的冠层生物量估算通常采用平均枝法或标准枝法(Ibrahim, 1995),存在测定困难、耗时耗力等问题,且因叶片水分损失较快也会带来难以估计的误差(刘琪璟, 2009; Sonetal., 2001)。随着遥感技术不断发展,快速、准确、大范围地估算冠层生物量成为可能,传统光学遥感估算冠层生物量的研究已十分成熟(Curranetal., 1992; 马泽清等, 2008; 闵志强等, 2010),但光学遥感只与叶生物量发生反应,且在生物量较高时具有局限性(Sinhaetal., 2015),无法实现对森林冠层生物量的准确测定。
与传统光学遥感相比,合成孔径雷达(synthetic aperture radar,SAR)波长较长,对树冠具有一定穿透能力,尤其是高频率SAR(X波段或C波段)能够获得丰富的冠层信息(主要为冠层中枝、叶的散射信息)(Santoroetal., 2007); 同时,SAR还可以穿透云层和一定程度的雨区,具有全天时全天候观测能力(陈尔学, 1999)。目前,SAR估算森林生物量主要利用后向散射和极化干涉技术(Toanetal., 1992; Dobsonetal., 1992; Metteetal., 2004; 宋茜等, 2011; 李兰等, 2017),但后向散射存在饱和点问题,极化干涉技术需要多基线或多频段的全相干极化SAR数据,对数据质量要求较高(Cloudeetal., 1999),而SAR估算冠层生物量的研究多集中于后向散射(Beaudoinetal., 1994; Kasischkeetal., 1995; Harrelletal., 1997)。极化目标分解是从极化SAR数据中提取地物极化特征的主要方法(Jawaketal., 2015),目前已有许多提取地物信息的极化分解方法(Krogager, 1990; Cloudeetal., 1996; Freemanetal., 1998; Yamaguchietal., 2005),且一些分解方法已用于生物量估算(Goncalvesetal., 2011; Kobayashietal., 2012; Chowdhuryetal., 2013)。
综上可以看出,利用SAR数据提取极化分解参数在森林冠层生物量估算中具有一定潜力和优势。本研究基于内蒙古赤峰市旺业甸林场2017年GF-3全极化SAR数据,采用Freeman三分量分解、Freeman二分量分解、Yamaguchi三分量分解提取SAR参数,并结合同一时期旺业甸林场外业调查数据,确定极化分解分量与森林冠层生物量的相关关系,构建冠层-地面散射比参数,应用多元逐步回归方法建立森林冠层生物量与SAR提取参数回归模型,旨在探讨GF-3全极化SAR数据在人工林冠层生物量估算中的潜力,提出一种准确估算森林冠层生物量的方法。
1 研究区概况、数据来源与研究方法
1.1 研究区概况 内蒙古赤峰市喀喇沁旗旺业甸林场(41°20′—41°40′N,118°10′—118°29′E)地处燕山山脉北麓七老图山支脉,平均海拔1 120 m。土壤以典型棕壤为主。属温带季风气候,年均降水量522.6 mm,年均气温3.9 ℃,年日照时间2 700 h以上。林场土地总面积25 958 hm2,有林地面积23 118 hm2,以人工林和通过封山育林形成的次生林为主,主要树种包括油松(Pinustabulaeformis)、华北落叶松(Larixprincipis-rupprechtii)、白桦(Betulaplatyphylla)、黑桦(Betuladahurica)等。
1.2 数据来源 1) 样地数据 采用2017年10月外业调查数据。选择林分整齐、受干扰小、具有代表性的区域设置样地,样地大小25 m×25 m。使用RTK对样地中心点和4个角点进行定位,共22块样地分布在SAR数据范围内,记录样地内胸径大于5 cm的树木胸径、树高和冠幅,其中1—10号样地为油松纯林,11—22号样地为华北落叶松纯林(表1)。

表1 样地数据统计Tab.1 Statistics data of plots
2) 全极化SAR数据 采用GF-3合成孔径雷达获取的C波段全极化单视复数数据,数据为L1级辐射定标产品,SAR数据获取于2017年8月5日,记录方式为16 bit复数形式,中心入射角29.7°,中心点坐标41°42′N、118°12′E,方位向分辨率5.3 m,距离向分辨率4.5 m,幅宽30 km×30 km(图1)。
1.3 研究方法
1.3.1 样地生物量计算 采用本研究区域油松单木生物量模型(马钦彦, 1989)和华北落叶松单木生物量模型(罗云建等, 2009)(表2),将样地每木检尺数据代入模型,分别计算单木枝、叶生物量。油松枝、叶生物量模型为二元模型,落叶松枝、叶生物量模型为一元模型,将枝、叶生物量之和作为单木冠层生物量,计算得到样地内所有单木冠层生物量,求和后除以样地面积(0.062 5 hm2),得到各样地冠层生物量(表3)。

图1 研究区域及样地分布Fig.1 Study area and sample plots distribution
1.3.2 SAR数据预处理 单视复数数据存在辐射误差,为了精确反映地物回波特性,对SAR数据进行辐射定标处理(Shimadaetal., 2009),定标系数根据数据头文件设置,获得SAR的后向散射矩阵[S],并随之生成极化相干矩阵[T]。

表2 枝、叶生物量模型①Tab.2 Biomass model of branches and leaves
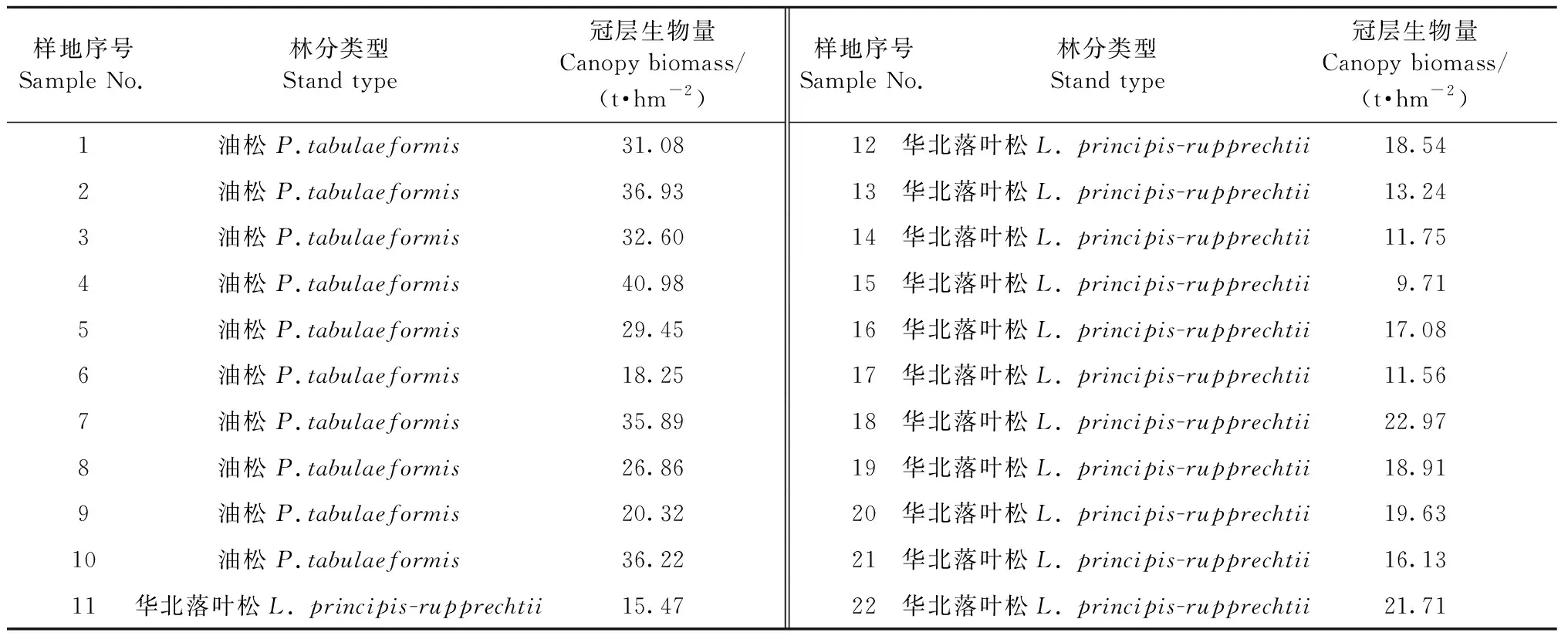
表3 样地冠层生物量Tab.3 Canopy biomass of sample plots
通常采用多视处理和滤波压制SAR数据固有的斑点噪声(Leeetal., 2009)。在生成极化相干矩阵[T]的过程中,方位向和距离向上分别设置视数为2,对SAR数据进行多视处理,该视数可保持处理后像元接近方形且尽量与样地大小相匹配,之后采用Refined Lee方法进行滤波,滤波窗口设置为7(Leeetal., 2002)。
为了消除地形对SAR数据的影响,应用AW3D 30(ALOS World 3D)数据对SAR数据进行正射校正。AW3D是日本宇宙航空研究开发机构(http:∥www.eorc.jaxa.jp)和日本遥感技术中心联合研制的全新一代高分辨率DEM,数据集水平分辨率约30 m,高程精度优于5 m,正射校正后SAR数据空间分辨率10 m,采用WGS84(world geodetic system-1984)地理坐标系和UTM(universal transverse mercator projection)的50 N带投影坐标系。
1.3.3 目标极化分解 极化分解可对目标的散射机制和特性进行合理物理解释。Cloude等(1996)根据目标散射特性将极化分解方法分为相干分解和非相干分解,本研究区域为森林,地物基本为非相干目标,适合采用非相干分解方法。非相干分解通常以二阶统计的极化协方差矩阵[C]和极化相干矩阵[T]为基础,一般对图像上每个像素邻域内协方差或相干矩阵进行平均来估算,得到空间统计平均后的极化协方差矩阵〈[C]〉和极化相干矩阵〈[T]〉,且二者可通过酉矩阵相互转换。极化相干矩阵更易于解释散射的物理过程,本研究应用极化相干矩阵进行极化分解,由于互易性的限制(Cloude, 2010),极化相干矩阵最终简化为三维〈[T3]〉。
采用Freeman三分量分解、Freeman二分量分解、Yamaguchi三分量分解3种极化分解方法,其中后二者对Freeman三分量分解模型进行了不同改进。
1) Freeman三分量分解 Freeman三分量分解(Freemanetal., 1998)是一种基于物理实际的三分量散射机制模型,为非相干极化分解方法,对森林植被后向散射具有良好解译能力,将森林植被后向散射分解为体散射、二次散射和表面散射3个在统计上独立不相关的分量,体散射代表冠层的直接散射,二次散射代表地面与树干间的二面角反射,表面散射代表地面的直接单次后向散射。本研究将3个分解分量功率分别表示为PV、PD、PS:
〈[T3]〉=fS〈[T3]〉S+fD〈[T3]〉D+fV〈[T3]〉V。
(1)
式中:fS、fD、fV为展开系数; 〈[T3]〉S、〈[T3]〉D、〈[T3]〉V分别为表面散射、二次散射、体散射的散射模型。
2) Freeman二分量分解 Freeman(2007)以Freeman三分量分解为基础,提出应用于森林的二分量分解模型,该模型的2种散射机制分别来自于冠层和地面,体散射分量代表来自冠层的直接散射,地面散射分量代表地面与树干间的二次散射或地面的直接单次后向散射。相比Freeman三分量分解固定的体散射模型,Freeman二分量分解在模型中加入了可变动参数,对体散射模型的限制更小。本研究将体散射、地面散射分量功率分别表示为PC、PG:
〈[T3]〉=fG〈[T3]〉G+fC〈[T3]〉C。
(2)
式中:fG、fC为展开系数; 〈[T3]〉G、〈[T3]〉C分别为地面散射、体散射的散射模型。
3) Yamaguchi三分量分解 Cui等(2012)在Freeman三分量分解的基础上提出了Yamaguchi三分量分解,该模型与Freeman二分量分解类似,同样在体散射模型中加入了可变动系数,但对系数范围进行了限制,且该分解方法首先对〈[T3]〉进行正交变换和酉变换(Anetal., 2010; Singhetal., 2011; Yamaguchietal., 2011),之后应用自适应体散射模型进行极化分解,得到体散射、二次散射、表面散射3个分量,功率分别表示为PV(θ)、PD(θ)、PS(θ):
〈[T3(θ)]〉=fS(θ)〈[T3(θ)]〉S+fD(θ)〈[T3(θ)]〉D+
fV(θ)〈[T3(θ)]〉V。
(3)
式中:fS(θ)、fD(θ)、fV(θ)为展开系数;〈[T3(θ)]〉S、〈[T3(θ)]〉D、〈[T3(θ)]〉V分别为表面散射、二次散射、体散射的散射模型。
1.3.4 参数构建 极化分解中,体散射分量代表森林冠层的后向散射信息,二次散射分量、表面散射分量代表SAR信号透过冠层后所获得的树干、地表后向散射信息。冠层散射与地面散射比值对森林冠层结构具有一定敏感性(Freeman, 2007),因此本研究分别将Freeman三分量分解、Yamaguchi三分量分解的体散射分量与二次散射分量和表面散射分量之积进行比值运算,将Freeman二分量分解体散射分量与地面散射分量进行比值运算,从而获得3个新的参数,各分量均在对数变换后参与运算,新参数可以看作各分解方法下的冠-地散射比。参数构建方法如下:

(4)

(5)
(6)
式中:PV、PD、PS为Freeman三分量分解的体散射、二次散射、表面散射功率;PC、PG为Freeman二分量分解的体散射、地面散射功率;PV(θ)、PD(θ)、PS(θ)为Yamaguchi三分量分解的体散射、二次散射、表面散射功率。
1.3.5 像元值提取 在提取固定样地对应的遥感信息时,根据样地中心点坐标将样地冠层生物量数据与SAR数据一一对应。由于样地面积为0.062 5 hm2,而SAR数据像元大小为10 m,所以在提取样地对应的SAR数据信息时,采用八邻域平均法提取,即样地中心点对应像元,相邻8个像元的像元值求平均,即为该样地对应的SAR数据。
1.3.6 相关性分析 相关性分析是通过相关系数来表示2个变量之间相关关系密切程度的统计方法。分别对冠层生物量、各极化分解分量进行以10为底的对数变换(Austinetal., 2003; Gamaetal., 2010),将变换后的冠层生物量数据与分解分量参数进行相关性分析,绘制散点图进行直观评价,并计算相关系数,从而确定变换的冠层生物量与分解分量之间的线性关系。
1.3.7 模型构建与检验 应用多元逐步回归方法构建模型,其基本思想是将自变量逐个引入模型中,对每个引入的解释变量进行显著性检验,保留显著变量,当引入的新变量使得原变量变得不再显著时剔除原变量,逐步回归获得最优模型。
为了评价模型的预测能力,需对模型进行评价检验,通常将数据分为建模样本和检验样本。本研究样地为22块,若保留部分样地数据用于验证将导致用于建模的样地数量过少,因此运用留一法交叉验证(leave-one-outcross-validation,LOOCV)对模型进行评价(Geisser, 1974; Stone, 1974; Woldetal., 1984),即每次将1块样地作为验证样本,其余21块样地作为训练样本进行建模,不断重复该过程并记录所有交叉验证结果,最终获得22组实测值和预测值对原始模型进行评价。该方法能够提供模型真实拟合能力的无偏估计(Cawleyetal., 2004),不会造成数据浪费,且检验过程中所得模型与使用全部数据所得模型基本一致。采用决定系数(R2)、均方根误差(RMSE)、预测误差平方和(PRESS)、平均偏差(ME)、平均绝对偏差(MAE)、平均相对偏差(MRE)、平均相对偏差绝对值(AMRE)和预测精度(P)对模型拟合结果和检验结果进行评价(李凤日, 2004; 董利虎, 2015),计算公式如下:
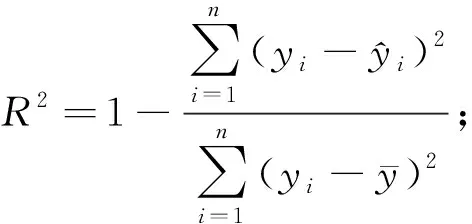
(7)

(8)

(9)
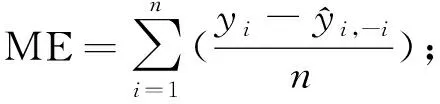
(10)

(11)
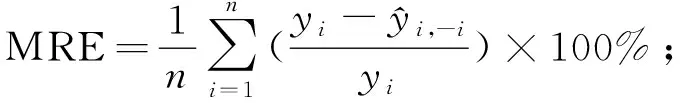
(12)
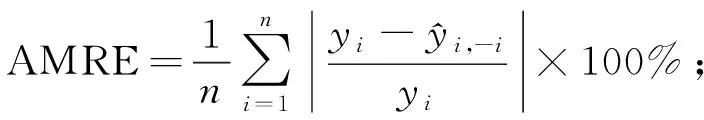
(13)
其中,
(14)

2 结果与分析
2.1 极化分解分量与冠层生物量的相关性 不同极化分解方法所得分量与冠层生物量均有较存在显著的负相关关系,Freeman三分量分解表面散射分量(PS)与冠层生物量在0.05水平上显著相关,其余分量均在0.01水平上与冠层生物量显著相关(表4),各极化分解方法提取分量对冠层生物量的变化均较为敏感。Freeman三分量分解体散射分量(PV)与冠层生物量的相关性高于二次散射分量(PD)和表面散射分量(PS)(图2a),Freeman二分量分解有着相似的特征(图2b),但Freeman二分量分解体散射分量(PC)与冠层生物量的相关性稍好于Freeman三分量分解体散射分量,说明在Freeman三分量和二分量分解中代表冠层散射特征的体散射分量对冠层生物量更为敏感,且Freeman二分量分解比Freeman三分量分解效果更好。Yamaguchi三分量分解体散射分量[PV(θ)]与冠层生物量的相关性和Freeman三分量分解体散射分量(PV)相似,而二次散射分量[PD(θ)]、表面散射分量[PS(θ)]与冠层生物量的相关性均高于体散射分量[PV(θ)]。在所有极化分解分量中,Yamaguchi三分量分解二次散射分量[PD(θ)]与冠层生物量的相关性最高(图2c)。
2.2 模型构建 本研究将对数变换后的冠层生物量(lgW)作为因变量,8个SAR极化分解分量(表4)和3个新构建冠-地散射比参数[R1、R2和R3,见式(4)、(5)、(6)]共11个参数作为自变量,进行逐步回归,最终冠-地散射比参数R2、R3进入回归模型,且具有较强的显著性,共线性容差大于0.5,二者间无明显多重共线性(表5)。模型R2为0.658,RMSE为4.943 t·hm-2。
2.3 模型检验 将应用多元逐步回归方法获得的最优模型进行留一法交叉验证,记录每次验证预测结果,以交叉检验所得冠层生物量预测值为自变量,以冠层生物量实测值为因变量,建立一元线性回归方程,并绘制散点图(图3)和残差图(图4),对模型进行评价(徐小军等, 2011)。结果表明,模型预测误差均较低,平均偏差(ME)为-0.665 t·hm-2,预测模型有轻微高估现象,平均绝对偏差(MAE)为4.845 t·hm-2,平均相对偏差(MRE)和平均相对偏差绝对值(AMRE)分别为3.33%和23.233%,预测精度(P)达91.5%,说明模型存在轻微偏差,但预测精度良好。对冠层生物量实测值与预测值之间的回归方程进行置信椭圆F检验,其F为0.145,小于F0.05(2,20)=3.49,表明实测值与预测值之间差异不明显(胡珍珠等, 2016; 王树文等, 2016),预测值大致分布在1∶1直线接近,模型未出现饱和点。模型标准化残差全部在2倍标准残差内,且呈随机分布,说明模型估算效果受生物量变化影响较小,具有良好的适用性和预测性。

表4 极化分解分量与冠层生物量的相关系数①Tab.4 Correlation coefficient of polarization decomposition component and canopy biomass
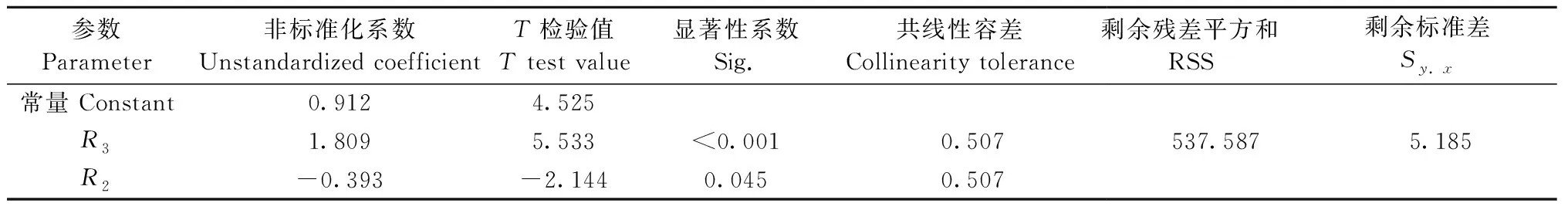
表5 模型回归系数Tab.5 Regression coefficients of the model

图3 实测值与模型预测值的相关性Fig.3 The correlation between measured value and model predicted value

图4 实测值-模型预测值标准残差Fig.4 Standardized residuals of measured value-model predicted value
3 讨论
3.1 极化分解分量与冠层生物量的关系 Kobayashi等(2012)采用Yamaguchi四分量分解估算热带森林蓄积,极化相干矩阵经正交变换后,表面散射、体散射与蓄积量的相关性变得稍好,表面散射与蓄积量为负相关关系,体散射与蓄积量为正相关关系,二次散射与蓄积量无相关性。Chowdhury等(2013)同样采用Yamaguchi四分量分解估算寒带森林蓄积,极化相干矩阵经正交变换后,二次散射与蓄积量的相关性显著提高,但对表面散射和体散射几乎没有影响。在Goncalves等(2011)的研究中,采用Freeman三分量分解估算热带森林蓄积,蓄积量与3个分解分量之间均呈显著正相关关系。
本研究采用3种极化分解方法提取分解分量,极化分解分量与冠层生物量均存在较为显著的相关关系,与Freeman三分量分解相比,Freeman二分量分解、Yamaguchi三分量分解的极化分解分量与冠层生物量相关性更好,二者对体散射模型进行了改进,均在体散射模型中加入可变系数,说明可变系数加入可提高森林区域极化分解效果,能够更准确地获取森林垂直结构信息。同时,Yamaguchi三分量分解效果稍好于Freeman二分量分解,Yamaguchi三分量分解在极化分解前对极化相干矩阵进行正交变换,改善了极化分解效果,从而提高了与冠层生物量的相关性,与Kobayashi等(2012)和Chowdhury等(2013)的研究结果在理论上一致。不同的是,本研究所得极化分解分量与冠层生物量均为负相关关系,这可能是因为二次散射和表面散射为SAR信号透过冠层的后向散射功率,冠层生物量越大,枝叶分布越密集,二次散射和表面散射功率越低; 体散射为SAR信号在冠层中的散射过程,C波段SAR穿透性较弱,随着冠层生物量增加,枝叶对SAR信号衰减作用增强,使得体散射功率降低。此外,分解分量特征受含水率、介电常数、灌木层、地形等多种因素影响, 森林结构、树叶及树干的形状和大小对生物量估算也存在一定影响(Santoroetal., 2009; Hoekmanetal., 2002; Rowlandetal., 2008)。
3.2 冠层生物量估算效果 Goncalves等(2011)应用多元线性回归建立了包括Freeman三分量分解在内的SAR参数与森林蓄积间的关系; Chowdhury等(2013)构建体散射分量、二次散射分量之积与表面散射分量的比值,与单独使用分解分量相比显著提高了与森林蓄积间的相关性。类似地,本研究构建各极化分解方法体散射分量与二次散射分量、表面散射分量之积的比值,应用多元线性回归建立SAR提取参数与森林冠层生物量回归模型,得到了较好结果(R2=0.658,RMSE=4.943 t·hm-2),相比于单一分解分量,冠-地散射比参数定量化描述了森林冠层散射与冠层下部散射间的关系,对森林的极化分解信息有效结合,当比值较大时,冠层生物量较高; 当比值较小时,冠层生物量较低。
由于数据限制,本研究仅采用22块样地数据进行建模,在统计学中属于小样本,虽然具有一定说服力,但还需要更多的样地观测数据进行验证(Garestieretal., 2009; Neumannetal., 2012; Gamaetal., 2010; Goncalvesetal., 2011)。同时,本研究仅对3种极化分解方法进行分析,更多分解方法对冠层生物量的估算效果有待进一步研究。
4 结论
1) 3种极化分解方法获得的极化分解分量均与冠层生物量具有显著相关关系,采用极化分解手段估算冠层生物量具有可行性。
2) Freeman二分量分解、Yamaguchi三分量分解的极化分解分量与冠层生物量相关性更高,说明极化相干矩阵旋转变换、体散射模型优化可有效提高极化分解效果,能够获取更丰富的森林垂直结构信息。
3) 相比于各极化分解分量,冠-地散射比参数对冠层生物量的敏感性更高,多种SAR极化分解参数共同使用能够较好估算冠层生物量,且未出现明显的饱和点。
应该指出的是,由于C波段SAR数据穿透性较弱,且本研究仅对Freeman三分量分解、Freeman二分量分解和Yamaguchi三分量分解3种极化分解方法估算冠层生物量的效果进行了探讨,应进一步研究其他分解方法估算冠层生物量是否与本研究获得相同的结论,验证本研究方法对估算地上生物量是否具有适用性。
