小贯小绿叶蝉在5个茶树品种(系)上的蜜露排泄量与茶树叶片结构比较
2020-10-16刘丰静冉伟李喜旺汪素琴孙晓玲
刘丰静,冉伟,李喜旺,汪素琴,孙晓玲*
小贯小绿叶蝉在5个茶树品种(系)上的蜜露排泄量与茶树叶片结构比较
刘丰静1,冉伟2,3,李喜旺2,3,汪素琴4,孙晓玲2,3*
1. 福建省农业科学院茶叶研究所,福建 福州 350013;2. 中国农业科学院茶叶研究所,浙江 杭州 310008;3. 农业部茶树生物学与资源利用重点实验室,浙江 杭州 310008;4. 绍兴市经济作物技术推广中心,浙江 绍兴 312000
为明确茶树叶片组织结构与小贯小绿叶蝉抗性之间的相关性,在确定小贯小绿叶蝉不同龄期和虫态蜜露排泄量的基础上,比较研究了小贯小绿叶蝉雌成虫在不同茶树品种(系)上取食24 h后蜜露排泄量的差异,进而通过比较不同茶树品种(系)叶片的组织结构和色泽,分析了5个茶树品种(系)叶片的组织结构与茶树对小贯小绿叶蝉抗性能力强弱的关系。研究结果表明,小贯小绿叶蝉4龄若虫的蜜露排泄量显著高于1~3龄;雌、雄成虫的蜜露排泄量之间不具显著差异;取食中选10号的雌成虫蜜露排泄量显著低于取食其他品种;不同品种(系)茶树的叶面积、叶片总厚度、上表皮厚度、下表皮厚度以及色差等指标与雌成虫的蜜露排泄量之间无显著相关性,而叶片茸毛密度与雌成虫蜜露排泄量呈显著负相关,叶片茸毛长度与蜜露排泄量呈显著正相关。
小贯小绿叶蝉;茶树品种(系);蜜露排泄量;叶片结构
长期以来,我国茶园中小绿叶蝉发生的优势种类一直被认为是假眼小绿叶蝉[(Goëthe)][1-3]。然而,Fu等[4]利用和16 S RNA作为分子标记发现我国茶园中叶蝉发生的优势种应与东亚地区的叶蝉为同一种类。直到Qin等[5]通过与模式标本的形态学比对,才将我国茶区小绿叶蝉的种名订正为小贯小绿叶蝉(Matsuda)。小贯小绿叶蝉以成、若虫刺吸茶树[(L.) O. Ktze]芽、叶及嫩梢为害,严重影响茶叶的产量和品质。雌成虫陆续孕卵和分批产卵的习性导致叶蝉在田间世代重叠严重,难于防治。据统计,小贯小绿叶蝉大暴发可引起30%~50%的茶叶产量损失[3,6]。长期以来,对小贯小绿叶蝉的有效防治主要依赖于化学农药。然而,化学农药的大量使用常导致环境污染、农药残留、害虫抗药性以及次生性害虫猖獗发生等问题。王念武等[7]的研究结果表明,福建福安茶场的茶小绿叶蝉对吡虫啉、啶虫脒和联苯菊酯的抗性指数已达到中抗至高抗的水平。因此,如何在科学控制小贯小绿叶蝉为害的前提下,减少化学农药的施用量和使用次数是科研工作者及一线生产者非常关心的问题。众所周知,抗虫品种的选育是害虫综合治理系统中的一个基本且重要的方面,寄主植物抗性则被认为是最有效和经济的抗虫策略之一。遗憾的是,目前茶树抗小贯小绿叶蝉品种的筛选与鉴定仅见少量报道[8-10],其研究的深度与广度显著滞后于水稻、小麦和玉米等其他作物。
植物叶片的表皮毛数量、角质层厚度或蜡质层厚度等指标是植物物理抗性的重要组成部分[11-12],相关研究在茄子(L.)、秋葵[(L.) Moench]和大豆[(L.) Merr]等植物上已有报道[13-15]。有研究发现茶树叶片的组织结构与茶树抗虫性之间具有一定的相关性[16-18]。例如,白鸡冠等12个茶树品种对黑刺粉虱[(Quaintance)]的抗性与茶树叶片组织结构之间的相关性研究结果显示,叶片气孔密度与黑刺粉虱的产卵量呈显著正相关,而叶片下表皮、角质层和栅栏组织的厚度与抗黑刺粉虱的能力呈显著正相关[16];金珊[18]通过调查小绿叶蝉在8个不同茶树品种上的发生量和每雌产若量,明确指出所选的茶树品种对小绿叶蝉的抗性存在差异,进而发现抗性茶树品种叶片的茸毛长度、密度和蜡质含量显著高于感性品种。然而,上述考察指标是否可用于小贯小绿叶蝉抗性茶树品种的早期鉴定尚需进一步研究。众所周知,田间虫口发生密度、每雌产若量和生命表是调查作物对害虫抗感能力的直观指标。但是,上述指标的采集不仅费时、费力,而且极易受当年害虫发生情况和气候因素的影响,具有很大的局限性。蜜露排泄量是衡量刺吸式口器害虫取食量的代表性指标,而取食量的差异又直接反映了害虫在植物被害组织上生长适合度的差异,可作为考察植物抗虫性的重要指标之一[19-20]。鉴于此,本文在确定小贯小绿叶蝉最大蜜露排泄量虫态的基础上,以5个茶树品种(系)为研究对象,研究了小贯小绿叶蝉在不同品种上取食24 h后蜜露排泄量的差异,进而通过比较不同茶树品种(系)叶片的组织结构和色泽的差异,明确了茶树叶片组织结构与茶树对小贯小绿叶蝉抗性强弱之间的关系。
1 材料与方法
1.1 供试茶树与小贯小绿叶蝉
中选2号、中选6号、中选7号、中选10号及福鼎大白茶茶树品种(系)均种植于中国农业科学院茶叶研究所南门品系比较区(北纬30°10′53′′,东经120°5′19′′,海拔30 m),5年实生苗,外观特征详见表1。采用自然鉴定法,在田间发现小贯小绿叶蝉在上述品种(系)上的为害程度不同。上述品种单行双株种植,行距1.5 m,株距0.33 m,树高0.50~0.70 m,冠层宽度0.80~0.90 m,茶园常规管理。选择长势良好,无病虫害发生的茶树嫩梢用于生测。
供试小贯小绿叶蝉若虫采集于中国农业科学院茶叶研究所龙井43无性系茶园,以龙井43嫩枝饲养于多功能人工气候室[温度(25±2)℃,相对湿度(75±5)%,光照时间L∶D=14∶10]。室内繁殖一代后,小贯小绿叶蝉雌成虫用于蜜露排泄量测定。
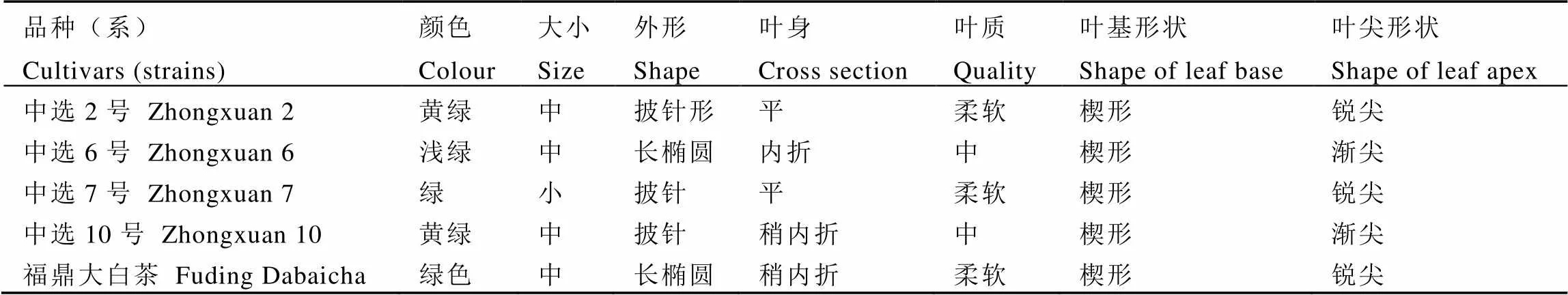
表1 茶树5个品种叶片的外观特征
1.2 试验方法
1.2.1 蜜露收集与排泄量测定
利用10 mL塑料指形管进行蜜露收集,采用万分之一电子天平(奥豪斯仪器上海有限公司)对蜜露进行称重。具体方法如下:试验前将10 mL指形管洗净并于60℃下烘干(4 h)后冷却至常温称重,记作1。从茶园中剪取一芽四叶的健康茶树嫩梢,剔除叶片后放入指形管。同时,放入1只饥饿2 h的小贯小绿叶蝉,底部用Parafilm M密封,茶枝插在浸过水的花泥上维持嫩梢新鲜。24 h后取出嫩梢和叶蝉并对指形管称重,重量记作2。蜜露排泄量的计算公式如下:蜜露排泄量=2-1。每个处理30个生物学重复。
蜕皮12 h以内的1~4龄小贯小绿叶蝉若虫和羽化12 h内的雌、雄成虫用于测试。比较小贯小绿叶蝉若虫不同龄期和成虫不同性别的蜜露排泄量时,统一采用单一品种福鼎大白茶。选择蜜露排泄量最高的虫态用于不同品种的测试。
1.2.2 叶片组织结构观察
分别采集5个茶树品种(系)健康植株上的芽下第二叶,在叶背近中脉处薄切下1 cm×0.5 cm表皮,(1)置于滴加有0.01%中性红溶液的载玻片上,在4×10倍光学显微镜下观察测量茸毛密度和长度;(2)置于FAA(福尔马林∶乙酸∶50%乙醇=1∶1∶18)中固定,番红整染后制成常规石蜡切片,观察并测量叶片总厚度和上下表皮厚度。另采集5个茶树品种(系)健康植株上的芽下第二叶,用色差计(CR2331C,香港美能达有限公司)测定叶片的亮度与色度。采集芽下第三叶测量叶面积。每个处理20个生物学重复。
1.2.3 统计分析
数据分析采用SPSS Statistics 21.0进行分析。小贯小绿叶蝉雌、雄成虫之间的蜜露排泄量差异显著性采用Student’s test进行比较;小贯小绿叶蝉若虫不同龄期之间、取食不同品种(系)的小贯小绿叶蝉雌成虫之间的蜜露排泄量差异显著性,以及各品种(系)间叶片组织结构特征和色泽的差异显著性分析均采用单因素方差分析(ANOVA),如果方差分析差异显著(˂0.05),则采用Turkey检验进行多重比较。
2 结果与分析
2.1 小贯小绿叶蝉若虫不同龄期和成虫不同性别的蜜露排泄量比较
取食24 h后,小贯小绿叶蝉4龄若虫的蜜露排泄量显著高于1~3龄。雌、雄成虫的蜜露排泄量之间不具显著差异。各龄若虫蜜露排泄量由大到小依次为4龄若虫˃3龄若虫˃2龄若虫˃1龄若虫(图1)。
2.2 取食不同茶树品种(系)的叶蝉雌虫蜜露排泄量比较
取食不同茶树品种(系)叶蝉的蜜露排泄量结果表明,取食中选10号的雌成虫的蜜露排泄量显著低于取食其他4个品种(系),但取食中选2号、6号、7号和福鼎大白茶的蜜露排泄量之间无显著差异(图2)。
2.3 茶树品种(系)叶片组织结构的差异比较
所选5个不同茶树品种(系)的叶片组织结构存在显著差异(表2)。中选10号的叶面积显著小于中选6号和7号,与中选2号和福鼎大白茶之间不具显著差异;中选10号的茸毛长度与中选6号之间无显著差异,却显著低于中选2号、7号和福鼎大白茶;但是,中选10号的茸毛密度显著高于其他4个品种(系);并且,中选10号上、下表皮的厚度显著高于中选2号、6号和7号,但与福鼎大白茶之间不具显著差异;福鼎大白茶的叶片总厚度显著高于中选2号、6号和7号,但与中选10号之间不具显著差异;色差测定结果表明,中选2号的色差值显著低于其他4个品种(系)。
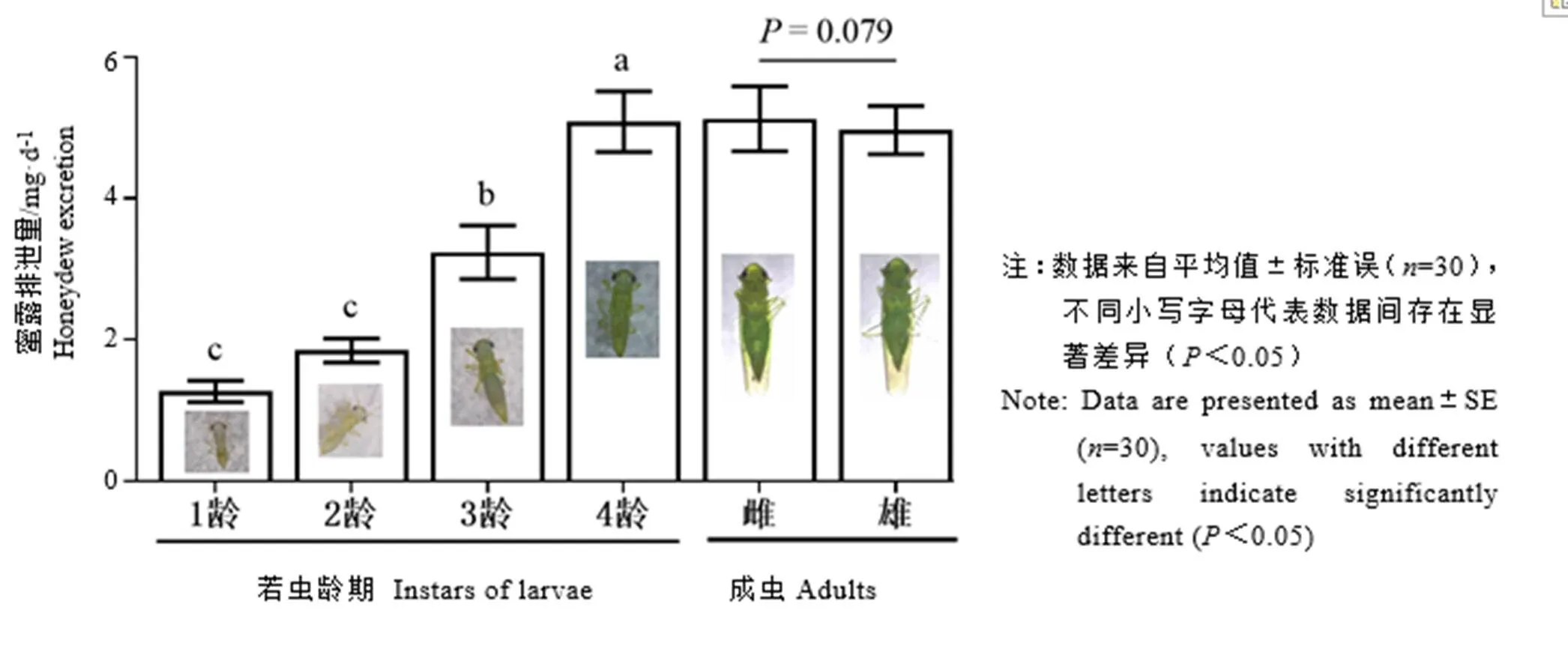
图1 小贯小绿叶蝉若虫不同龄期和成虫不同性别的蜜露排泄量比较

图2 取食不同品种(系)小贯小绿叶蝉雌成虫的蜜露排泄量比较

表2 5个茶树品种(系)叶片组织结构的差异比较
注:A、L、D、T、Tupper、Tlower和C分别指叶片面积、茸毛长度、茸毛密度、叶总厚度、上表皮厚度、下表皮厚度和色差值;数据来自平均值±标准误(n=20),同一列的不同字母表示该指标在不同茶树品种(系)之间存在显著性差异(<0.05)
Note: A, L, D, T, Tupper, Tlowerand C represent leaf area, hair length, hair density, total thickness, thickness of upper epicuticle, thickness lower epicuticle and value of chromatism, respectively. Data are presented as mean±SE (n=20), the same column followed by different letters meant significant difference in the different tea cultivars (strains) (<0.05)
3 讨论
小贯小绿叶蝉在我国各大茶区均有发生,且以成、若虫刺吸茶树汁液为害,常给茶叶生产带来严重的经济损失。雌成虫具有陆续孕卵和在嫩茎中分批产卵的习性,是对茶叶安全生产影响最大的虫态。本研究结果显示,在测试的所有虫龄和虫态中,小贯小绿叶蝉雌成虫的取食量最大。因此,在小贯小绿叶蝉若虫发生高峰期进行有效防治,将会极大减轻小贯小绿叶蝉对茶叶生产造成的经济损失。蜜露排泄量测定结果表明,随着虫龄的增长,小贯小绿叶蝉若虫蜜露的排泄量从1龄到4龄逐渐增多,此结果符合害虫生长发育对营养物质需求量逐渐增大的规律[21]。
利用寄主植物的组成抗性进行害虫防治,是害虫综合治理体系的一个重要方面。叶片的物理结构是影响植物抗虫性强弱的重要因素之一。有研究发现,在进化过程中植物可以通过改变叶片的形态来抵御植食性昆虫的为害[22-24]。大豆叶片茸毛的着生角度越小,大豆植株对豆卷叶螟(Fabricius)的抗性则越强[13]。此外,大量研究结果表明,植物各组织部位的颜色、形状、大小、茸毛、刺突、角质层和蜡质层等指标都可能与植物抵御刺吸式口器害虫的为害有关[14-15, 25-26]。前期研究发现,中选10号对黑刺粉虱的抗性与其他4个品种之间不具显著差异[27]。本文的研究结果表明,取食中选10号的小贯小绿叶蝉雌成虫蜜露排泄量显著低于取食其他品种(系)的叶蝉。可见,与其他4个茶树品种相比,中选10号对小贯小绿叶蝉的抗性可能较强。进一步的分析发现,5个不同茶树品种(系)芽下第二叶的叶面积、叶片总厚度、上表皮厚度、下表皮厚度或色差等指标与叶蝉雌成虫蜜露排泄量的多少不具明显的相关性;但是叶片茸毛密度与雌成虫蜜露排泄量呈显著负相关(=–0.85),叶片茸毛长度与蜜露排泄量呈显著正相关(=0.84)。由此说明,茶树叶片上茸毛密度越大则茶树抗叶蝉的能力越强,反之茸毛长度越长则茶树抗叶蝉的能力越弱。这一研究结果与前人的研究结果基本相符。例如,德清、举岩、长兴紫笋和建德4个抗性茶树品系的茸毛密度显著高于竹山一号、恩标、蓝天和斑竹园等4个感性品种[18];西花蓟马()通过诱导番茄()植株腺体毛密度的增加而提高为害植株对后续西花蓟马的抗性[28]。金珊[18]研究发现,德清和举岩等4个抗性品种的茸毛长度显著高于恩标和蓝天等4个感性品种,但是目前大部分的研究[29-30]则认为叶片的茸毛长度与植物的抗虫能力之间无显著相关性。由此推断,植物对植食性昆虫的组成抗性是由植物形态、组织结构和化学因子等多个方面组成,单一性状指标不能作为抗性品种筛选的唯一指标。
综上所述,与其他4个茶树品种(系)相比,中选10号对小贯小绿叶蝉的抗性能力较强,并发现茶树叶片的茸毛密度可能是影响茶树对叶蝉抗性能力的重要指标之一。
致谢:本文所用茶树品种由中国农业科学院茶叶研究所遗传育种团队王新超课题组提供,河南农业大学暑期实习生张孟可参与部分工作,特此感谢。
[1] 吕文明, 陈琇, 罗其忍. 小绿叶蝉发生规律及其防治研究[J]. 茶叶科学, 1964, 1(1): 45-55. Lyu W M, Chen X, Luo Q R. Research on ocurence and control of[J]. Journal of Tea Science, 1964, 1(1): 45-55.
[2] 葛钟麟, 张汉鹄. 中国茶叶蝉种类研究(一)[J]. 茶业通报, 1988(1): 15-18. Ge Z L, Zhang H G. Research on the Cicadelidae species damaging Chinese tea shrub (I) [J]. Journal of Tea Busines, 1988(1): 15-18.
[3] 赵冬香, 陈宗懋, 程家安. 茶小绿叶蝉优势种的归属[J]. 茶叶科学, 2000, 20(2): 101-104. Zhao D X, Chen Z M, Cheng J A. Belongingnes of tea leaf-hopper dominant species [J]. Journal of Tea Science, 2000, 20(2): 101-104.
[4] Fu J Y, Han B Y, Xiao Q. Mitochondrialand 16sRNA evidence for a single species hypothesis of, J. formosana andin East Asia [J]. Plos One, 2014, 9(12): e115259.doi: 10.1371/journal.pone.0115259.
[5] Qin D Z, Zhang L, Xiao Q, et al. Clarification of the identity of the tea green leafhopper based on morphological comparison between Chinese and Japanese Specimens [J]. Plos One, 2015, 10(9): e0139202. doi: 10.1371/journal.pone.0139202.
[6] 彭萍, 唐敏, 侯渝嘉, 等. 黄板诱杀茶园黑刺粉虱及假眼小绿叶蝉效果及特性研究[J]. 西南农业大学学报, 2010, 23(1): 87-90. Peng P, Tang M, Hou Y J, et al. Study on the effect and characteristics of yellow sticky trap stickingandGothe in tea garden [J]. Southwest China Journal of Agricultural Sciences, 2010, 23(1): 87-90.
[7] 王念武, 徐金汉, 陈峥, 等. 不同茶园假眼小绿叶蝉抗药性比较[J]. 福建农业大学学报, 2004, 33(2): 169-173. Wang N Y, Xu J H, Chen Z, et al. Resistance level of(Gothe) in different tea plantations [J]. Journal of Fujian Agriculture and Forestry University, 2004, 33(2): 169-173.
[8] 刘丽芳. 茶树不同品种和次生代谢物质对叶蝉取食行为的影响的DC-EPG研究[D]. 北京: 中国农业科学院, 2011. Liu L F. Studies on effect of various cultivars and secondary metabolites of tea plants to tea green leafhopper feeding by DC-EPG [D]. Beijing: Chinese Academy of Agricultural Sciences, 2011.
[9] 金珊, 孙晓玲, 陈宗懋, 等. 不同茶树品种对假眼小绿叶蝉的抗性[J]. 中国农业科学, 2012, 45(2): 255-265. Jin S, Sun X L, Chen Z M, et al. Resistance of different tea cultivars toGöthe [J].Scientia Agricultura Sinica, 2012, 45(2): 255-265.
[10] 王蔚, 吴满容, 张思校, 等. 茶小绿叶蝉在福建省茶树品种上的选择机制初探[J]. 河南农业科学, 2016, 45(4): 80-84. Wang W, Wu M R, Zhang S X, et al. Preliminary study on selection mechanism ofon tea varieties in Fujian Province [J].Journal of Henan Agricultural Sciences, 2016, 45(4): 80-84.
[11] 赵延霞. 沙棘组织结构及对沙棘木蠹蛾的抗虫性[D]. 北京: 北京林业大学, 2012. Zhao Y X.Tissue structure of sea buckthorn and insect Resistance to[D]. Beijing:Beijing Forestry University, 2012.
[12] 李雅菲. 菊花和除虫菊腺体形态结构和内含物与其抗蚜性的相关性研究[D]. 武汉: 华中农业大学, 2011. Li Y F.Comparative study on the morphology and inelusions of triehomes and aphids-resistance between florist's chrysanthemum cultivars and pyrethrum [D]. Wuhan: Huazhong Agricultural University, 2011.
[13] 邢光南, 谭连美, 刘泽稀楠, 等. 大豆地方品种叶片叶柄茸毛性状的形态变异及其与豆卷叶螟抗性的相关分析[J]. 大豆科学, 2012, 31(5): 691-696. Xing G N, Tan L M, Liu Z X N, et al. Morphological variation of pubescence on leaf blade and petiole and their correlation with resistance to Bean Pyralid (Fabricius) in soybean landraces [J].Soybean Science, 2012, 31(5): 691-696.
[14] Hasanuzzaman A T M, Islam M N, Zhang Y, et al. Leaf morphological characters can be a factor for intra-varietal preference of whitefly(Hemiptera: Aleyrodidae) among eggplant varieties [J]. Plos One, 2016, 11(4): e0153880.
[15] Halder J, Sanwal S K, Deb D, et al. Mechanisms of physical and biochemical basis of resistance against leafhopper () in different okra () genotypes [J]. Indian Journal of Agricultural Sciences, 2016, 86(4): 481-484.
[16] 王庆森, 黄建, 陈常颂, 等. 茶树种质叶片组织结构与其对黑刺粉虱抗虫性的关系[J]. 茶叶科学, 2009, 29(1): 60-66.Wang Q S, Huang J, Chen C S, et al. Relationship between leaf structure of tea germplasm and its resistance to(Quaintance) [J]. Journal of Tea Science, 2009, 29(1): 60-66.
[17] 叶火香, 韩善捷, 韩宝瑜. 假眼小绿叶蝉趋色性及茶梢信息物质强化其趋色性的效应[J]. 安徽农业大学学报, 2015, 42(6): 868-872. Ye H X, Han S J, Han B Y. Phototaxis of tea green leafhopper and strengthening effect of infochemicals from tea shoots on its phototaxis [J]. Journal of Anhui Agricultural University, 2015, 42(6): 868-872.
[18] 金珊. 不同茶树品种抗假眼小绿叶蝉机理研究[D]. 西安: 西北农林科技大学, 2012.
Jin S. Resistance Mechanisms of tea plant cultivars to tea green leafhopper [D]. Xi′an: Northwest A&F University, 2012.
[19] 郭贵明, 苏芝业, 杨伦伦, 等. 麦长管蚜蜜露分泌量与小麦品种的关系研究初报[J]. 山西大学学报, 1993, 16(2): 228-231. Guo G M, Su Z Y, Yang L L, et al. A preliminary report on the relationship between the(Fab.) honeydew secretion and wheat varieties [J]. Journal of Shanxi University, 1993, 16(2): 228-231.
[20] 周晓静, 白素芬, 李欣, 等. 蜜露检测法在转基因棉抗蚜性指标中的应用[J]. 河南农业大学学报, 2012, 46(1): 53-57.
Zhou X J, Bai S F, Li X, et al. Application of honeydew colormetry in determining indexes of aphid resistance of transgenic insect resistant cottons [J]. Journal of Henan Agricultural University, 2012, 46(1): 53-57.
[21] 钦俊德. 植食性昆虫食性的生理基础[J]. 昆虫学报, 1980, 23(1): 106-122. Qin J D. The physiological basis of herbivores [J]. Acta Entomologica, 1980, 23(1): 106-122.
[22] Mick E Hanley, Byron B L, Meredith M F, et al. Plant structural traits and their role in anti-herbivore defence [J]. Perspectives in Plant Ecology, Evolution and Systematics, 2007, 8(4): 157-178.
[23] Patricia G Z, Alfonso E, Josefa B. Costs of leaf reinforcement in response to winter cold in evergreen species [J]. Tree Physiology, 2016, 36(3): 273-286.
[24] Duy N, Ivo R, Celestina M. How plants handle multiple stresses: hormonal interactions underlying responses to abiotic stress and insect herbivory [J]. Plant Molecular Biology, 2016, 91: 727-740.
[25] 曹贺贺. 小麦对麦长管蚜的组成抗性和诱导抗性研究[D]. 杨凌: 西北农林科技大学, 2014. Cao H H. Constitutive and indecud defenses of wheat against[D]. Yangling:Northwest A&F University, 2014.
[26] 赵曼. 不同玉米品种(系)对玉米蚜抗性机制的初步研究[D]. 郑州: 河南农业大学, 2013. Zhao M. Prelimiary research on the resistance mechanism of different maize hybrids (inbreds) to(Fitch) [D]. Zhengzhou:Henan Agricultural University, 2013.
[27] 李喜旺, 辛肇军, 孙晓玲. 利用黄板监测不同茶树品种(品系)上黑刺粉虱成虫的种群消长及日节律变化[J]. 茶叶学报, 2015, 56(1): 51-55. Li X W, Xin Z J, Sun X L.Monitoring population dynamic and circadian rhythm ofadults on different tea varieties (strains) by using yellow sticky trap [J].Acta Tea Sinica, 2015, 56(1): 51-55.
[28] Escobar-Bravo R, Klinkhamer P G L, Leiss K A. Induction of jasmonic acid-associated defenses by thrips alters host suitability for conspecififics and correlates with increased trichome densities in tomato [J]. Plant and Cell Physiology, 2017, 58(3): 622-634.
[29] 林凤敏, 吴敌, 陆宴辉, 等. 棉花叶片茸毛性状与绿盲蝽抗性的关系[J]. 植物保护学报, 2010, 37(2): 165-171. Lin F M, Wu D, Lu Y H, et al. The relationship between trichome characteristics of cotton leaves and the resistance to[J]. Acta Phytophylacica Sinica, 2010, 37(2): 165-171.
[30] 桂连友, 龚信文, 孟国玲. 茄子叶片绒毛与侧多食跗线螨抗性的关系[J]. 应用生态学报, 2007, 18(1): 229-232. Gui L Y, Gong X W, Meng G L. Relationship between eggplant leaf pubescent and its resistance to[J]. Chinese Journal of Applied Ecology, 2007, 18(1): 229-232.
The Comparison of Leaf Structures andMatsuda Honeydew Excretion among Five Tea Cultivars (Strains)
LIU Fengjing1, RAN Wei2,3, LI Xiwang2,3, WANG Suqin4, SUN Xiaoling2,3*
1. Tea Research Institute of Fujian Academy of Agricultural Sciences, Fuzhou 350013, China; 2. Institute of Tea Research, Chinese Academy of Agricultural Sciences, Hangzhou 310008, China; 3. Key Laboratory of Tea Biology and Resources Utilization, Ministry of Agriculture, Hangzhou 310008, China; 4. Shaoxing Economic crop Technology Extension Center, Shaoxing 312000, China
The food consumptions offemale adults feeding on the leaves of five tea cultivars (strains) were compared by weighing the honeydew excretion after 24 h. The leaf structures and colors of five tea cultivars (strains) were also compared. Further analysis illustrated the relationship between plant resistance toand leaf structures. The results show that: (1) the food consumption of female adults feeding onZhongxuan 10 was significantly lower than those feeding on other cultivars (strains), and there was no significant difference between the male and female adults. (2) The leaf area, blade thickness, upper skin thickness, lower skin thickness and chromatic aberration of different tea cultivars (strains) had little effect on’s honeydew excretion. The density of leaf pubescence was negatively correlated to the honeydew excretion, but the length of leaf pubescence had a positive correlation with the honeydew excretion.
Matsuda, tea cultivars (strains), honeydew excretion, leaf structure
S571.1;S435.711
A
1000-369X(2020)05-625-07
10.1371/journal.pone.0153880.
2020-07-23
2020-08-24
国家茶叶产业技术体系咀嚼式害虫防控(CARS-19)
刘丰静,男,助理研究员,主要从事茶树害虫防控研究。*通信作者:xlsun1974@163.com
