长江中游宜昌江段鱼类早期资源现状及水文影响条件
2020-10-15黎明政刘焕章段中华曹文宣
陈 诚 黎明政 高 欣 姜 伟 刘焕章 段中华 曹文宣
(1. 中国科学院水生生物研究所水生生物多样性与保护重点实验室, 武汉 430072; 2. 中国科学院大学, 北京 100049; 3. 中国长江三峡集团公司中华鲟研究所, 宜昌 443100; 4. 三峡工程鱼类资源保护湖北省重点实验室, 宜昌 443100)
鱼类早期资源指精卵结合至早期稚鱼阶段, 是鱼类个体生活史的开端[1]。通过对鱼类早期资源的研究, 可以了解鱼类的繁殖活动、产卵场位置分布和繁殖时间等早期生活史特征[1]。同时, 早期资源也是影响种群数量变化的主要原因之一[2]。然而,捕捞、水利工程建设和江湖阻隔等人类活动造成了鱼类早期资源的急剧减少[3,4]。例如, Sanches等[5]研究结果显示, 水利工程导致巴拉那河的鱼苗密度有显著变化。过度捕捞造成地中海沿海区域鱼类的产卵活动明显减少[6]。
长江中游是多种鱼类的产卵场和栖息地。刘明典等[7]2014—2015年在宜昌江段调查到27种鱼类卵苗。李世健等[8]在监利江段调查到51种鱼类的鱼卵和仔稚鱼。黎明政等[9]在武穴江段调查到27种鱼类的卵苗。同时, 长江中游也是重要经济鱼类四大家鱼的主要产卵场之一。20世纪60年代, 长江中游四大家鱼的产卵场有25处, 产卵量占长江总量的四分之三。其中, 宜昌江段是四大家鱼产卵规模最大的产卵场,平均鱼卵径流量可达119.48×108粒, 占长江干流总量的9.7%[10]。20世纪80年代, 在葛洲坝修建之后, 长江中游宜昌至城陵矶江段仍有11处四大家鱼产卵场, 产卵规模为11.14×108粒, 占长江干流总规模的25.8%[11]。但是, 在三峡水库蓄水之后, 长江中游四大家鱼的繁殖活动受到了不利的影响[12]。同时, 过度捕捞, 尤其是对仔幼鱼的捕捞, 造成了鱼类资源的急剧减少[13]。此外, 三峡水库蓄水之后造成长江中游宜昌江段四大家鱼产卵时间推迟约20d, 监利江段宜昌江段四大家鱼的产卵规模减少约75%[14]。为了有效地开展长江鱼类资源的保护工作, 有必要掌握长江中游鱼类早期资源的现状及其变化规律。
本研究通过在宜都断面开展产漂流性卵鱼类早期资源调查, 了解宜昌江段产漂流性卵鱼类的繁殖情况; 结合历史资料, 分析四大家鱼及其他鱼类的繁殖规模和时间变化; 分析水文因子对产漂流性卵鱼类优势种类繁殖的影响; 为长江中游鱼类早期资源保护工作的开展提供科学依据。
1 材料与方法
1.1 样本的采集
2017年5月12日至7月25日和2018年5月10日至7月24日, 在葛洲坝下游宜都市清江河口上约1.5—3.0 km处设置采样断面(112°26′E, 30°24′N)(图 1)。采用产漂流性卵鱼类早期资源的调查方法。采样网具包括弶网和圆锥网, 采集方法参照曹文宣等[1]和黎明政等[14]的方法。弶网用于每日采样, 记录种类和数量。圆锥网用于采集断面数据,计算断面系数, 每年采集一次。弶网网口呈半圆形,网口直径1 m, 网目尺寸500 μm, 网口面积0.393 m2,网长2 m。网后部连接集苗箱, 集苗箱长40 cm, 宽和高均为30 cm, 上部敞口, 用于收集鱼卵和仔鱼。弶网采集在每日上午和下午各一次, 采集时间为7:30—9:00和16:30—18:00, 每次采集持续30—60min。同时, 测量采样点水温、透明度、流速等环境参数,水位和流量数据来自于全国水雨情网站(http://xxfb.mwr.cn/)。圆锥网用于断面定量采样。网口呈圆形, 网口直径0.5 m, 网口面积0.196 m2, 网长2 m, 网目尺寸500 μm。圆锥网设置在断面左岸近岸点、左岸至江中心1/2处、江中心、江中心至右岸1/2处以及右岸近岸处5个采样点, 每个采样点设置表层、中层和底层3个采样水层(表层、中层和底层距水面的距离分别占水深的20%、50%和80%), 每个水层采集15min。
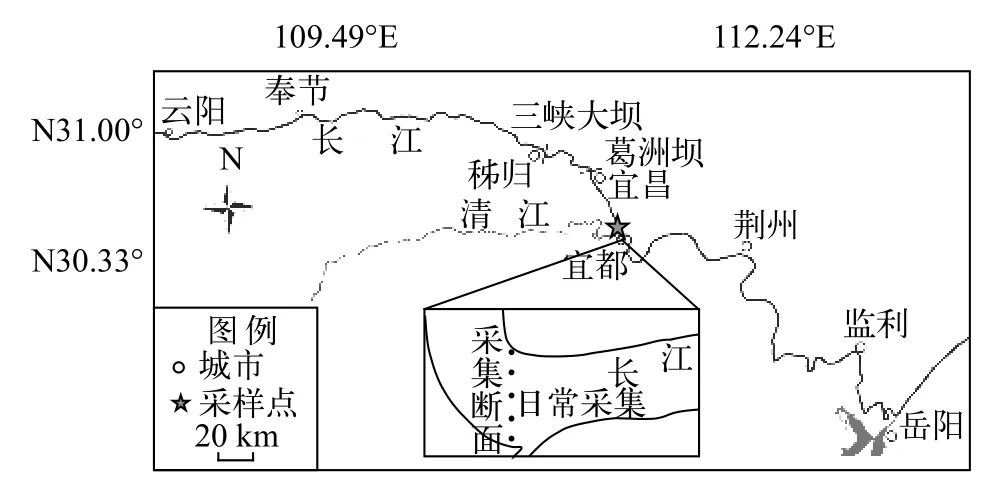
图1 2017年和2018年长江中游宜都断面鱼类早期资源采样图Fig. 1 Samling map of fish eggs and larvae in the Yidu section of the Yangtze River in 2017 and 2018
1.2 种类鉴定
鱼卵按照卵膜性质(漂流性、黏性和浮性)分类, 并依据卵径大小、胚体长等标准将大小不同的鱼卵区分培养, 培养水温维持在20—25℃, 直至孵出发育至分类特征明显的阶段, 按照形态、色素分布、肌节数目等进行种类鉴定, 具体方法参照曹文宣等[1]; 仔鱼通过显微镜观察进行种类鉴定, 之后样本用95%的乙醇保存。形态观测不能确定种类的样本用95%的乙醇固定保存在-20℃冰箱, 带回分子实验室, 通过线粒体DNA细胞色素b基因进行种类鉴定, 具体实验方法参照Xiao等[15]的方法进行。
1.3 数据处理
产卵规模计算鱼卵和仔鱼通过断面径流量的估算方法参照易伯鲁等[10]、黎明政等[14]和母红霞等[16]的方法进行, 具体计算公式如下:
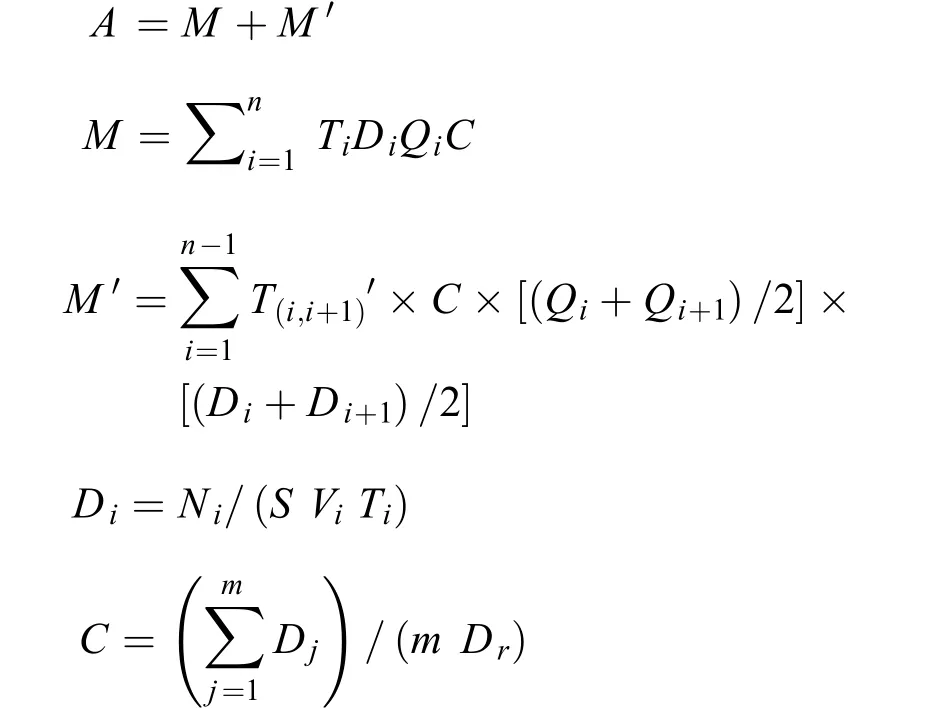
式中,A为整个采样期内流经采集点江段面的鱼卵、仔鱼总流量;M为所有定时采集时间内鱼卵、仔鱼流量之和;M'为前后两次相邻采集之间非采集时间内计算出的鱼卵、仔鱼流量之和;i为某一特定的常规采样;i+1为第i次采样之后的一次采样;Qi为第i次采样时流经采样断面的水流量(m3/s);Di为第i次采样时流经采样断面的仔鱼和鱼卵密度(ind./m3);C为采样断面上的鱼卵和仔鱼的分布系数;T(i,i+1)为第i次采样和第i+1次采样之间的时间间隔;Ni为第i次采样中采集到的鱼卵和仔鱼数量;S为网口面积(m2);Vi为第i次采样中网口的水流速(m/s);Ti为第i次采样的持续时间(s);Dj为断面采样每个采样点的鱼卵和仔鱼密度(ind./m3);j为第j个断面采样位点;m为断面采样的采样点数量;Dr为断面采样中流经靠近右岸的采样点表层的鱼卵和仔鱼密度(ind./m3)。
确定产漂流性卵鱼类优势种类物种优势度利用Pinkas相对重要性指数(IRI, Index of relative abundance)[17]确定, 计算公式如下:
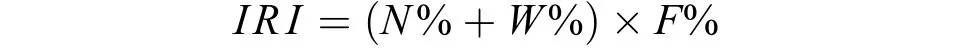
式中,N为某一种类鱼卵数量占采获鱼卵总数量的百分比;W为某种鱼卵生物量比例;F为某一种类出现的次数占调查总次数的百分比。由于鱼卵、鱼苗很小, 生物量难以确定, 因此只考虑数量这一因素。将IRI大于1000的种类定义为优势种。
四大家鱼产卵场位置推算四大家鱼产卵场位置的计算公式如下:
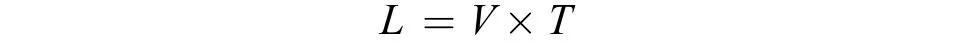
式中,L为四大家鱼卵、苗的漂流距离(m);V为调查江段水流速度(m/s);T为胚胎发育所需要的时间(s)。其中, 四大家鱼胚胎发育所需要的时间参照易伯鲁等[18]。
繁殖活动的水文条件分析根据相关研究文献资料, 近些年一段断面所采集的仔鱼大多来源于三峡大坝以上江段, 鱼卵来自三峡大坝下游产卵场[14,19]。因此,鱼类繁殖水文因子影响分析仅针对通过鱼卵鉴定的产漂流性卵鱼类的优势种类。首先, 对物种的密度及水文因子数据进行除趋势对应分析(Detrended correspondence analysis, DCA), 根据每个轴的梯度长度(Lengths of gradient, LGA)选择适合的分析方法。当LGA<3时, 采用冗余分析(Redundancy Analysis, RDA); 当LGA>4时, 采用典型对应分析(Canonical Correlation Analysis, CCA);当4>LGA>3时, 两种分析方法皆可[20]。DCA分析结果轴梯度长度为1.8, 故本研究利用冗余分析(Redundancy Analysis,RDA)分析鱼卵密度与环境因子(水温、流量、流量日变化、透明度)之间的相关关系。
DCA和RDA分析利用Canoco 5.0[21]软件完成,其余数据分析与作图用Excel 2010软件中完成。
2 结果
2.1 种类组成及优势种
2017年和2018年共采集鱼卵21120粒和仔鱼2123尾。物种鉴定结果显示, 鱼卵和仔鱼共5目9科37种。其中, 鲤科(Cyprinidae) 23种, 鳅科(Cobitidae) 5种, 鲿科(Bagridae)及科(Sisoridae)各2种, 银鱼科(Salangidae)、平鳍鳅科(Homalopteridae)、科(Hemiramphidae)、科(Serranidae)和虎鱼科(Gobiidae)各1种, 鲤科鱼类资源数量为20788粒, 占总数量的89.44%,其中鲌亚科为优势类群, 占鲤科鱼类总数量的39.45%。根据卵的性质, 漂流性卵24种, 黏性卵5种, 浮性卵1种, 沉性卵7种(表 1)。
2017年和2018年采集到鱼卵28种, 有青鱼(Mylopharyngodon piceus)、草鱼(Ctenopharyngodon idellus)、赤眼(Squaliobarbus curriculus)、贝氏(Hemiculter bleekeri)、(Hypophthalmichthys nobilis)、鲢(Hypophthalmichthys molitrix)和银(Squalidus argentatus)等种类。采集到仔鱼27种,有太湖新银鱼(Neosalanx taihuensis)、寡鳞飘鱼(Pseudolanbuca engraulis)、贝氏、间下(Hyporhamphus intermedius)和子陵吻鰕虎鱼(Rhinogobius giurinus)等种类。
2017年, 采集鱼卵10572粒, 仔鱼1221尾, 其中四大家鱼卵3172粒, 贝氏卵2725粒, 分别占卵总数量的30.0%、25.8%; 仔鱼以贝氏和寡鳞飘鱼为主, 分别占比54.3%、24.4%。2018年共采集鱼卵10548粒, 仔鱼902尾, 其中四大家鱼卵4249粒, 贝氏卵2448粒, 分别占卵总数量的40.3%和23.2%; 仔鱼以子陵吻虎鱼、寡鳞飘鱼和贝氏为主, 分别占比22.7%、19.3%和17.7%。根据2017年和2018年采集卵和仔鱼数量分析, 宜都断面鱼卵优势种为四大家鱼(IRI=1433)和贝氏(IRI=1319); 仔鱼优势种为贝氏(IRI=1259)和寡鳞飘鱼(IRI=1169)。
2.2 产卵时间
仔鱼5月采集较少, 2017年5月仅采集仔鱼5尾,分别为间下、光泽黄颡鱼和光唇蛇, 2018年5月采集仔鱼16尾, 分别为太湖新银鱼、寡鳞飘鱼和贝氏。仔鱼采集数量主要在6月和7月。2017年和2018年采集结果显示, 仔鱼中贝氏和寡鳞飘鱼为优势种类(图 2)。
2.3 产卵规模
2017年, 通过宜都断面的鱼卵径流量为124.45×108粒, 鱼卵中四大家鱼径流量为36.13×108粒, 占总径流量的29.03%; 贝氏径流量为19.96×108粒, 占总径流量的16.04%。2018年通过宜都断面的鱼卵径流量为101.07×108粒, 鱼卵中四大家鱼径流量为35.20×108粒, 占总径流量的34.83%; 贝氏的鱼卵径流量为19.17×108粒, 占总径流量的18.97%。
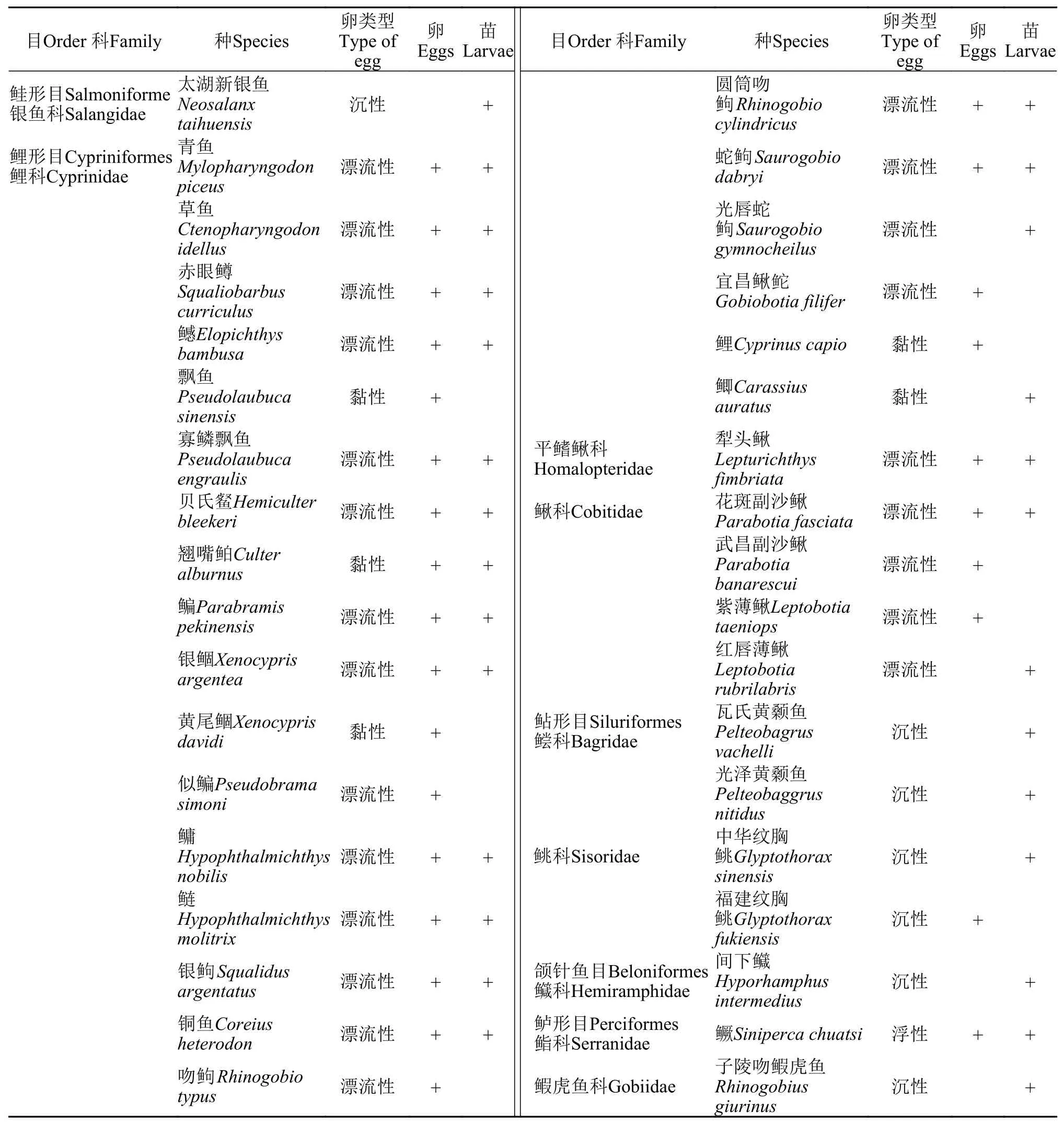
表1 2017年和2018年调查期间宜都断面鱼类早期资源种类组成Tab. 1 Species composition of eggs and larvae in the Yidu section in 2017 and 2018
2017年通过宜都断面的仔鱼径流量为16.43×108尾, 寡鳞飘鱼和贝氏径流量分别为5.89×108尾和2.91×108尾, 占仔鱼总径流量的35.87%和17.72%。四大家鱼的总径流量为4.12×108尾, 占仔鱼总径流量的25.07%。2018年通过宜都断面的仔鱼径流量为8.29×108尾, 寡鳞飘鱼和贝氏分别为0.97×108尾和1.22×108尾, 占仔鱼总径流量的11.72%和14.70%。四大家鱼的总径流量为0.11×108尾, 占仔鱼总径流量的1.32%。
2.4 四大家鱼的产卵场位置
通过对采集到的四大家鱼卵发育期的观测,0.14%处于128细胞期前, 95.31%处于桑椹期至眼囊期, 4.55%处于尾芽期至心脏搏动期, 距受精1—33h。其中, 65.76%的四大家鱼卵处于原肠早期至原肠晚期, 距受精7—13h。四大家鱼仔鱼均处于鳔雏形期和鳔一室期, 距受精98—146h。根据鱼卵和仔鱼的发育期以及流速分析, 四大家鱼鱼卵来源于宜都断面2.4 km以上的江段。产卵场位置主要分布在葛洲坝下至庙咀、胭脂坝至云池江段。四大家鱼仔鱼应来源于三峡大坝以上江段(图 3)。
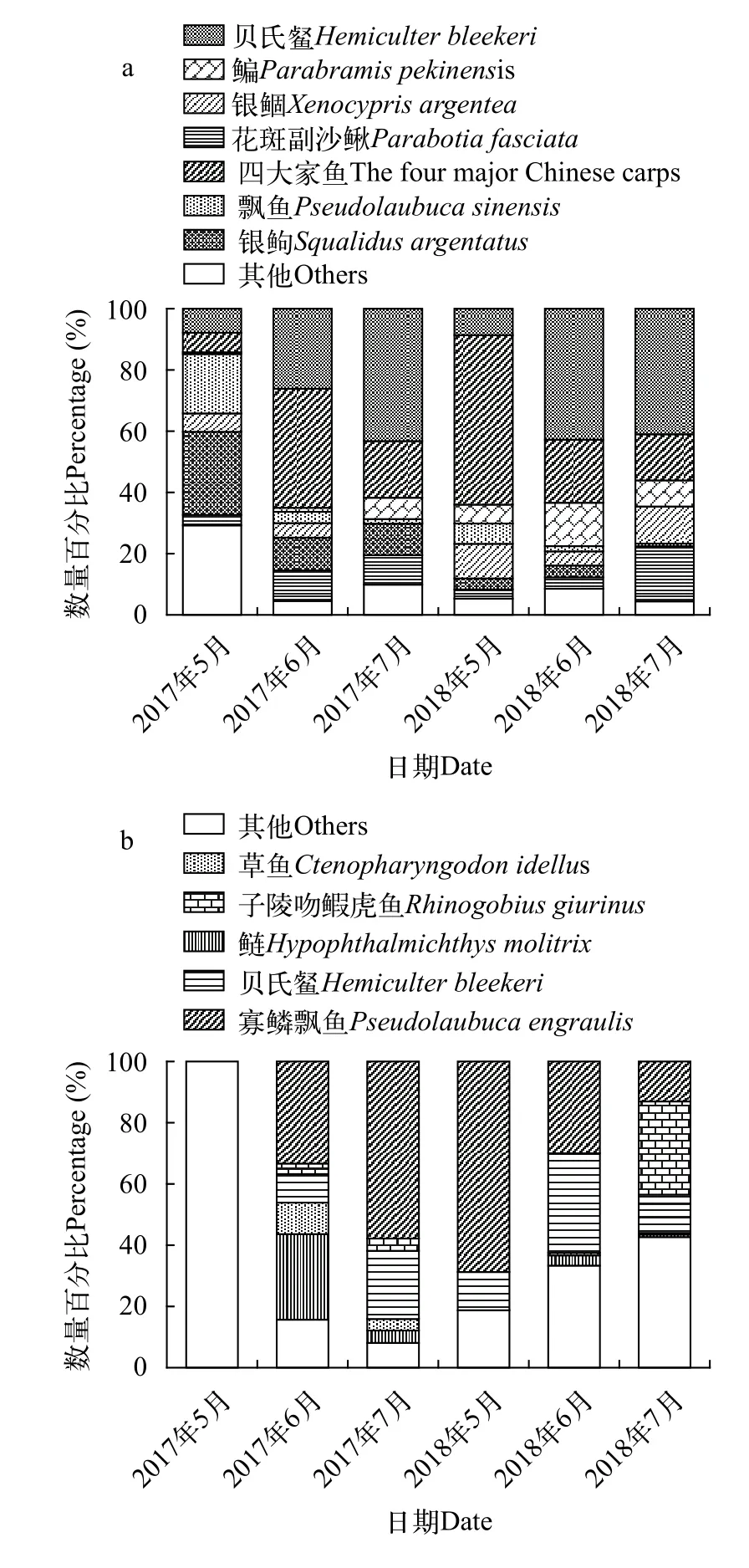
图2 2017年和2018年宜都断面主要类群鱼卵(a)、仔鱼(b)的相对丰度Fig. 2 Abundance percentage of eggs (a) and larvae (b) of dominant species in the Yidu section in 2017 and 2018
2.5 产漂流性卵鱼类及关键环境因子
2017年最小流量在7月4日, 为6920 m3/s, 最大流量在7月11日, 为30200 m3/s。整个调查期间, 水温介于18.5—26.0℃, 5月12日达到18℃以上, 7月20日达到24℃以上。2017年出现4次产卵高峰, 分别在6月10日、6月11日、6月18日和7月10日。高峰期产卵规模总计为60.7×108粒, 占2017年总产卵规模的48.7%。
2018年6月12日流量最小, 为10200 m3/s, 最大流量在7月14日, 为43800 m3/s。整个调查期间, 水温为18.5—25.2℃, 5月10日达到18℃以上, 7月2日达到24℃以上。2018年出现2次显著的产卵高峰,分别为5月19日和6月24日, 产卵规模共为35.9×108粒, 占2018年采集鱼卵总规模的35.5%(图 4)。
应用RDA分析方法, 对鱼卵漂流密度与水文因子的关系进行分析。在前2个排序轴上, 物种与水文因子的相关性为0.426和0.345, 第一排序轴和第二排序轴的累积贡献率为92.83%。RDA分析结果显示, 四大家鱼卵总密度和花斑副沙鳅卵密度受流量日变化和流量影响较大; 贝氏鱼卵密度受流量影响较大; 鳊鱼卵密度受水温影响较大; 银鱼卵密度主要受透明度影响(图 5)。
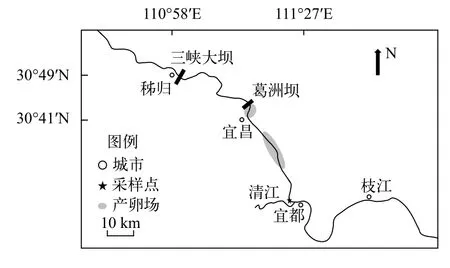
图3 长江中游宜昌江段四大家鱼产卵场位置Fig. 3 Spawning grounds of the four major Chinese carps in the Yichang reach in the middle Yangtze River
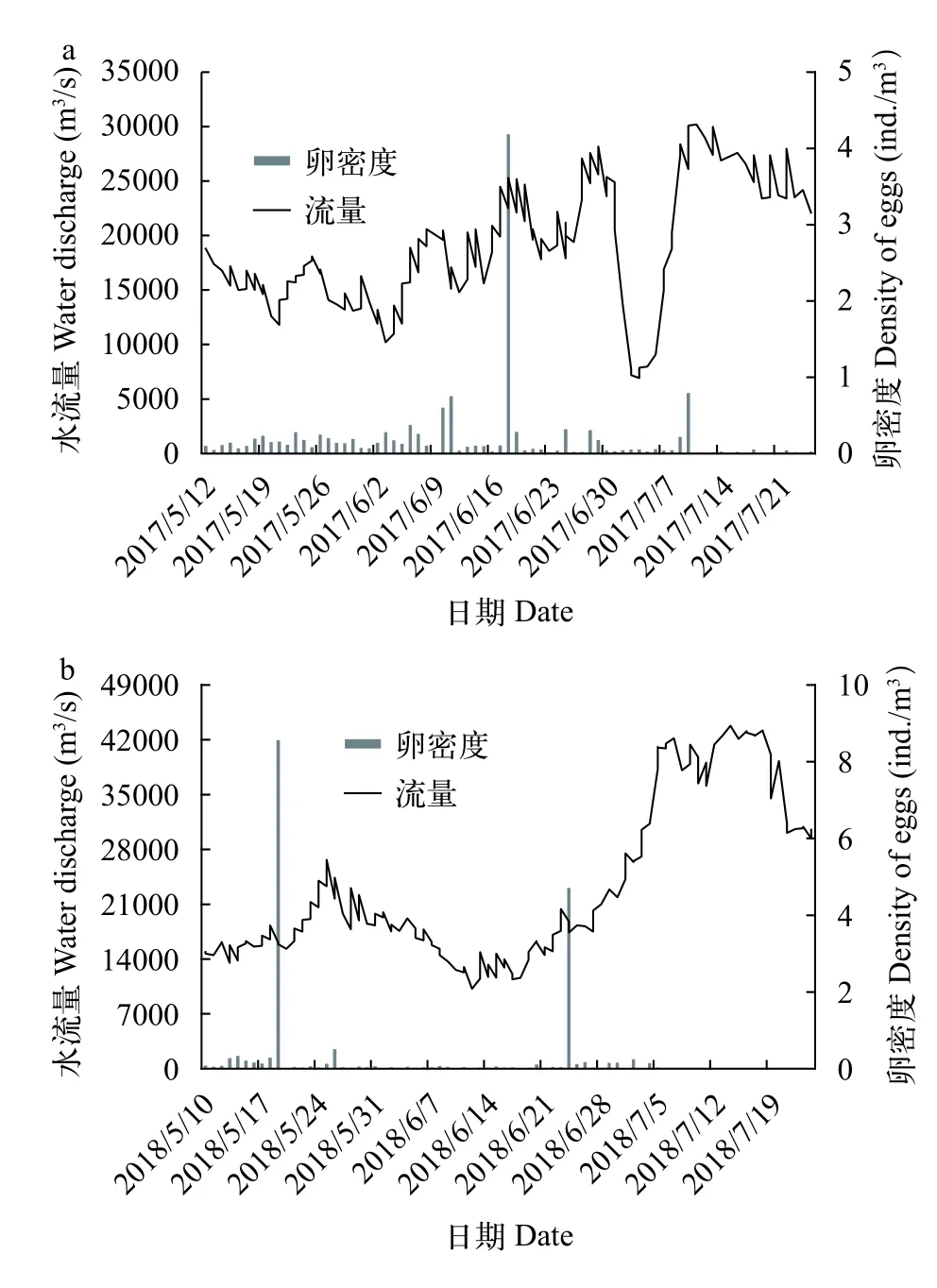
图4 2017年(a)和2018年(b)调查期间宜都断面鱼卵日密度变化图Fig. 4 Discharge and eggs density drifting through the Yidu section in 2017 (a) and 2018 (b)

图5 2017年和2018年宜都断面水文因子对漂流性卵密度的影响Fig. 5 Effects of hydrological factors on the density of the drifting eggs in the Yidu section in 2017 and 2018
3 讨论
3.1 长江中游鱼类繁殖现状及变化
本研究结果显示, 2017年和2018年5—7月通过长江中游宜都断面的鱼类早期资源较丰富。已有的研究结果显示, 母红霞[22]发现2009年和2010年5—7月通过宜都断面鱼卵径流量分别为57.26×108粒和64.45×108粒, 年均60.86×108粒。与2009年和2010年同比, 2017年和2018年通过宜都断面的鱼卵径流量明显增加, 年均径流量增加了85.3%。其中, 宜昌江段四大家鱼的产卵规模明显增大。2005—2012年5—7月通过宜都断面四大家鱼鱼卵径流量为(0.03—5.85)×108粒, 年均2.49×108粒[14], 而2017年和2018年四大家鱼卵年均35.67×108粒, 同比增加了13倍, 达到了20世纪80年代的水平[10,11]。鱼卵径流量增加的主要原因可能有2个。第一,2009年起, 原农业部在长江中游开展较大规模的四大家鱼亲鱼放流, 增加了繁殖群体数量[7,23]。第二,2011年三峡工程开始生态调度试验, 在一定程度上促进了鱼类的繁殖活动。徐薇等[24]的研究结果显示, 2012年三峡水库试验性生态调度期间长江中游沙市江段鱼卵总径流量和家鱼卵径流量均出现高峰, 繁殖种类明显增加。周雪等[25]的研究结果显示,2013—2017年三峡水库生态调度造成长江中游监利江段涨水持续时间达到4d及以上, 期间四大家鱼的卵苗量较大, 占年总量的45%以上。
在三峡水库蓄水运行之后, 长江中游四大家鱼的产卵时间推迟。在三峡水库蓄水之前, 长江中游宜昌江段四大家鱼产卵时间从4月中下旬开始[10]。在蓄水后, 2005—2012年宜昌江段四大家鱼的产卵时间推迟到5月8日至5月28日, 平均推迟了21.8d, 导致产卵时间推迟的主要原因是下泄水温滞后, 到达18℃的时间推迟[14]。与2005—2012年相比, 2017年和2018年宜昌江段四大家鱼产卵时间没有明显变化, 产卵场位置也没有明显变化。因此, 本研究的结果表明, 在三峡水库蓄水运行之后, 长江中游四大家鱼产卵场的环境条件可能已经趋于稳定。如果亲本增加, 宜昌江段产卵场依然可以满足其完成繁殖活动, 产生较大规模的鱼卵和仔鱼。
本研究结果还显示, 2017年和2018年5—7月有一定规模的仔鱼通过长江中游宜都断面, 其中四大家鱼的仔鱼均来源于三峡大坝以上江段。《长江三峡水利枢纽环境影响报告书》认为, 在三峡水库蓄水之后, 长江上游涪陵以下的8个四大家鱼产卵场将消失, 产卵场将上移。根据母红霞[22]、王红丽等[26]、Li等[14]和马琴[27]在三峡水库库区的鱼类早期资源调查结果显示, 涪陵以下江段仍有产卵规模较小的四大家鱼的产卵场。2005—2012年, 通过宜都断面四大家鱼仔鱼径流量年均值为4.65×108尾[14]。2013年和2014年分别为1.52×108尾和1.27×108尾[27]。与2005—2014年相比, 2017年和2018年没有明显的变化。研究结果表明, 三峡水库库区内在一定水文条件下仍具备四大家鱼繁殖的条件, 且可以通过三峡大坝进入长江中游, 可以为长江中游鱼类资源提供一定的补充。
3.2 鱼类早期资源与环境因子的关系
本研究结果显示, 四大家鱼、贝氏䱗和花斑副沙鳅等种类的产卵规模与流量和流量日变化有着密切的关系。鱼类的繁殖活动受外界环境因子的影响[28]。已有的研究提出“最佳环境窗口”假说, 认为每个物种都有特定的环境条件需求, 以满足繁殖活动和保证产生最多的后代[29—31]。Li等[32]的研究结果显示, 四大家鱼的产卵活动需要18—24℃的水温和0.55 m/d的水位涨幅。Wang等[12]也发现, 四大家鱼的仔鱼丰度与流量增加持续时间显著相关。因此, 水温应该是四大家鱼等产漂流性卵鱼类的繁殖限制条件, 达到了一定水温鱼类才能开始繁殖活动[10,33]。流量等则是刺激鱼类繁殖和促进繁殖活动的重要环境条件[10,34]。
3.3 长江中游鱼类早期资源保护措施
鱼类早期资源减少导致种群补充量减少[35,36],是造成长江鱼类资源急剧减少的主要原因之一[1,37]。Ma等[38]的研究结果显示, 长江中游对于通江湖泊中鱼类早期资源的补充, 维持鱼类多样性有重要意义。为了长江鱼类资源保护, 2019年农业农村部、财政部和人力资源社会保障部联合印发《长江流域重点水域禁捕和建立补偿制度实施方案》, 长江禁渔已经逐步实施, 2021年长江将全面禁渔。这一措施将消除捕捞对鱼类资源的影响, 尤其是对湖区等仔幼鱼资源的影响, 能够显著提高鱼类早期生活史阶段的存活率, 增加种群的补充量。长江鱼类的早期生活史阶段还面临着栖息地减少的问题。长江中下游的湖泊和河漫滩原本是仔幼鱼的主要栖息地。但是, 长江中下游的通江湖泊仅剩石臼湖、洞庭湖和鄱阳湖, 其余湖泊都已经被阻隔, 仔幼鱼无法进入湖泊生长, 其栖息地功能已经完全丧失。Liu等[39]的研究结果显示, 长江中下游江湖连通性减少已经严重影响了鱼类多样性。同时, 港口建设、防洪护坡和城市景观等建设破坏了河漫滩的自然环境条件, 减少了仔幼鱼的栖息地[40,41]。因此,为了更加有效地开展鱼类资源保护, 结合本研究结果, 还建议通过恢复江湖连通、灌江纳苗, 以及维持河漫滩自然环境条件, 增加仔幼鱼的栖息地面积;开展重要经济鱼类亲本放流, 增加繁殖群体规模;开展生态调度, 促进繁殖活动。
致谢:
感谢中国科学院水生生物研究所胡华明、李文静、杨萍、张文武和张晨在样本采集、数据处理上给予的帮助!
