四川石渠县牦牛体表蜱携带无形体和疏螺旋体的分子检测
2020-10-14唐天才刘城成袁东波阳爱国杨晓农郝力力
唐天才,刘城成,魏 巍,袁东波,郭 莉,侯 巍,莫 茜,阳爱国,杨晓农,郝力力*
(1.西南民族大学 生命科学与技术学院,四川 成都 610041;2.中国动物疫病预防控制中心,北京100125;3.四川省动物疫病预防控制中心,四川 成都 610041)
目前,无形体科(Anaplasmataceae)可分为无形体属(Anaplasma)、埃立克体属(Ehrlichia)、埃及小体属(Eegyptianalla)、沃尔巴克体属(Wolbachia)、新立克次体属、(Neorickettsia)考德里体属(Cowdria)和客鲍体属(Xenohalioti)7个属,其中无形体属和埃立克体属成员是一类致病性较强病原体[1]。莱姆病(Lyme disease)是由伯氏疏螺旋体引起的一种经蜱传播的人兽共患病,据报道,现已有全世界五大洲70多个国家有莱姆病的存在,我国多省区有人感染莱姆病的报道,并且发病区域与发病率呈扩大和上升的趋势,已被世界卫生组织(WHO)列为重点防治研究对象[2-3]。蜱作为一种体表吸血寄生虫,是无形体和疏螺旋体等多种人兽共患病原体的主要传播媒介,给人类健康和畜牧业发展带来严重的危害。近年来,国内相继有关于不同地区不同蜱种传播无形体和疏螺旋体的报道,并且多呈现复合感染。如韩茜等[4]从江西赣州市的长角血蜱、嗜群血蜱、具角血蜱和微小扇头蜱中检测到立克次体和埃立克体;李红雨等[5]从新疆北疆地区的亚洲离眼蜱、刻点血蜱、草原革蜱和边缘革蜱中检测到无形体和疏螺旋体;韩蓉等[6]从青海的青海血蜱、阿坝革蜱、草原革蜱、森林革蜱、丹氏血蜱检测到无形体、疏螺旋体、立克次体以及梨形虫。
石渠县是四川省海拔最高面积最大的纯牧业县,境内平均海拔4 200 m,牦牛是当地最重要和数量最多的家畜,整体养殖方式粗放,驱虫意识淡薄,更重要的是牧民与牦牛接触极其紧密,极易被寄生于牦牛体表的蜱叮咬继而感染相关蜱媒病。此前本实验室研究显示石渠县存在青海血蜱与西藏革蜱,并对蜱传巴尔通体和斑点热群立克次体进行检测与分析,但其他相关蜱媒病的资料仍是一片空白。因此,本研究对蜱传无形体和疏螺旋体进行调查,以期为该地今后蜱媒病的防控提供数据支持。
1 材料与方法
1.1 样本采集于2018年3—8月从石渠县麻甲乡、德荣马乡、阿日扎乡及长须干玛乡的牦牛体表采集蜱,装入收集管,塞入潮湿脱脂棉,带回实验室。根据《中国畜禽寄生虫形态分类图谱》[7]及《中国经济昆虫志》[8],在体式显微镜下对蜱进行形态鉴定,再放入75%酒精溶液中,并置于4℃冰箱保存待检。
1.2 样本DNA提取取出用75%酒精保存的样品,无菌水洗涤晾干后延纵向中轴线切开,分别装入EP管,半只于-80℃冰箱冻存,另外半只加入500 μL 无菌水于Bertin Precellys 24研磨器充分研磨,参数如下:每管加入陶瓷珠(直径0.5 mm的20珠和2 mm的5珠),5 500 r/min研磨2次;4 000 r/min研磨1次。取研磨液200 μL,按照EasyPure Genomic DNA Kit试剂盒的(北京全式金公司)说明书提取DNA,于-20℃保存备用。
1.3 PCR检测蜱的分子鉴定按照本实验室前期所采用的方法进行[9],上游引物LCO1490:GGTCAACAAATCATAAAGATATTGG,下游引物HCO2198:TAAACTTCAGGGTGACCAAAAAA-TCA。而无形体与疏螺旋体的检测分别采用DU等[10]、ELLEN等[11]和JUNG等[12]报道的方法(表1)。所有引物扩增均采用25 μL的反应体系,2×Easy Tag PCR Super Mix 12.5 μL、上下游引物各1 μL(10 μmol/L)、模板1 μL、ddH2O 9.5 μL混匀。PCR扩增条件如下:94℃预变性4 min;94℃变性15 s,55℃退火35 s,72℃延伸,40个循环;72℃延伸8 min;16℃保温(表1)。取2 μL PCR扩增产物于1.2%琼脂糖凝胶中100 V电泳30 min,凝胶成像仪中观察结果。

表1 引物相关信息
1.4 序列分析及统计分析阳性产物送生工测序,所得序列用DNAStar软件进行拼接分析,于NCBI中BLAST进行比对,再应用MEGA6.0软件以Neighbor-Joining法(bootstrap值1 000)构建系统进化树,进行聚类分析,并用SPSS软件对数据进行统计学分析。
2 结果
2.1 蜱鉴定结果共捕获818只蜱,经形态学鉴定出2种蜱,即青海血蜱与西藏革蜱,其中青海血蜱172只,占总数的21.03%;西藏革蜱646只,占总数的78.97%,为优势种。
青海血蜱雄蜱与雌蜱形态特征大致相同,假头基呈矩形位于躯体前端,躯体窄长,饱血后约8.2 mm×4.5 mm,呈卵圆形,须肢粗短,躯体背面覆盖有几丁质盾板,肛沟围绕肛门之后;无珐琅斑无眼,足基节只有1个距,各基节均有粗短内距,末端钝,略微超出该节后缘,各附节较粗短,口下板齿式5/5,压舌板形;雄蜱气门板呈长逗点形,雌蜱气门板呈椭圆形。
西藏革蜱雄蜱与雌蜱形态特征区别明显,雄蜱:体型较小,呈卵圆形,未吸血躯体长宽约4 mm×2 mm;假头短且呈矩形、须肢外缘呈弧形、整个盾板有混杂不均一的银白色花斑;气门板逗点状、雄蜱腹面无几丁质板、各足基节依次增大,4对足短而粗;肛沟围绕肛门之后。雌蜱气门板椭圆形、背突短钝,背缘无几丁质增厚部;盾板分两部分,前段端有珐琅彩,后端无银白相间的花斑;其他特征与雄虫一样。
分子生物学鉴定结果显示,本研究的青海血蜱与分离自甘肃的青海血蜱同源性达99.09%,而本研究的西藏革蜱与分离自西藏的西藏革蜱同源性达99.50%,表明形态学鉴定与分子生物学鉴定结果一致。
2.2 蜱媒病原PCR扩增结果以提取DNA为模板,分别扩增无形体和疏螺旋体的目的基因片段,阳性样本均出现与预期相符的特异性条带。
2.3 蜱携带病原检测结果无形体、疏螺旋体结果如表2所示,在818只蜱中,有132只检测到了无形体,总阳性率为16.14%(132/818),其中麻甲乡、德荣玛乡、阿日扎乡以及长须干玛乡的阳性率分别为8.93%、28.13%、14.28%和17.09%。经χ2检验,麻甲乡西藏革蜱与青海血蜱的无形体阳性率无显著差异(χ2=0.127,P>0.05);麻甲乡的无形体阳性率显著的低于德荣玛乡和长须干玛乡(P<0.05)。另外,只在青海血蜱中检测到牛无形体和埃立克体,其阳性率分别为6.98%和2.33%,经χ2检验,青海血蜱中牛无形体阳性率显著的高于埃立克体(χ2=4.195,P<0.05);只在西藏革蜱中检测到绵羊无形体,其总阳性率为17.96%。基于疏螺旋体flaB基因,有8只蜱检测到了疏螺旋体,阳性率为0.98%(8/818),再对这8份阳性样品进行16S rRNA基因扩增,结果均为阳性,其中西藏革蜱中未检测到疏螺旋体。此外,有3只青海血蜱混合感染了牛无形体与疏螺旋体。
2.4 核酸序列比对及遗传进化分析将所得序列拼接、两两比对后,得到5条不同的无形体序列,分别命名为M2-4、M2-13、A1-4、X1-42和M1-4,相似性为91.1%~99.7%。疏螺旋体序列只得到1条,命名为Borreliasp. Shiqu。将这些序列于GenBank中进行比对,选择对应相似性最高的序列2~3条以及不同种的序列作为参考序列,分别构建无形体16S rRNA基因、疏螺旋体flaB基因以及疏螺旋体16S rRNA基因的系统进化树(图1,2,3)。本研究中得到的绵羊无形体(A1-4和X1-42)16S rRNA 基因序列与西安(MG869525)、新疆(KJ410245)和日本(LC194134)分离到的绵羊无形体同源性均最高,分别达100%和99.74%。牛无形体(M2-4)16SrRNA基因序列与美国(AY144729)、俄罗斯远东地区(JX092092)分离到的牛无形体序列同源性最高,达99.74%;与重庆(MF289479)分离到的牛无形体序列同源性达99.22%。牛无形体(M2-13)与日本(LC012813)分离到的牛无形体序列同源性最高,达99.74%。埃立克体(M1-4)16Sr RNA基因序列与新疆(KU844280)、武汉(KX987325)、福建(DQ324547)分离到的Ehrlichiasp.同源性均为100%(图1)。本研究中得到的疏螺旋体flaB基因序列与分离自日本大刺血蜱中的Borreliasp.HM菌株序列(LC170031)的同源性最高,达98.94%(图2);疏螺旋体16S rRNA基因序列与分离自日本大刺血蜱中Borreliasp.HM菌株序列(LC170035)的同源性最高,达99.84%(图3)。
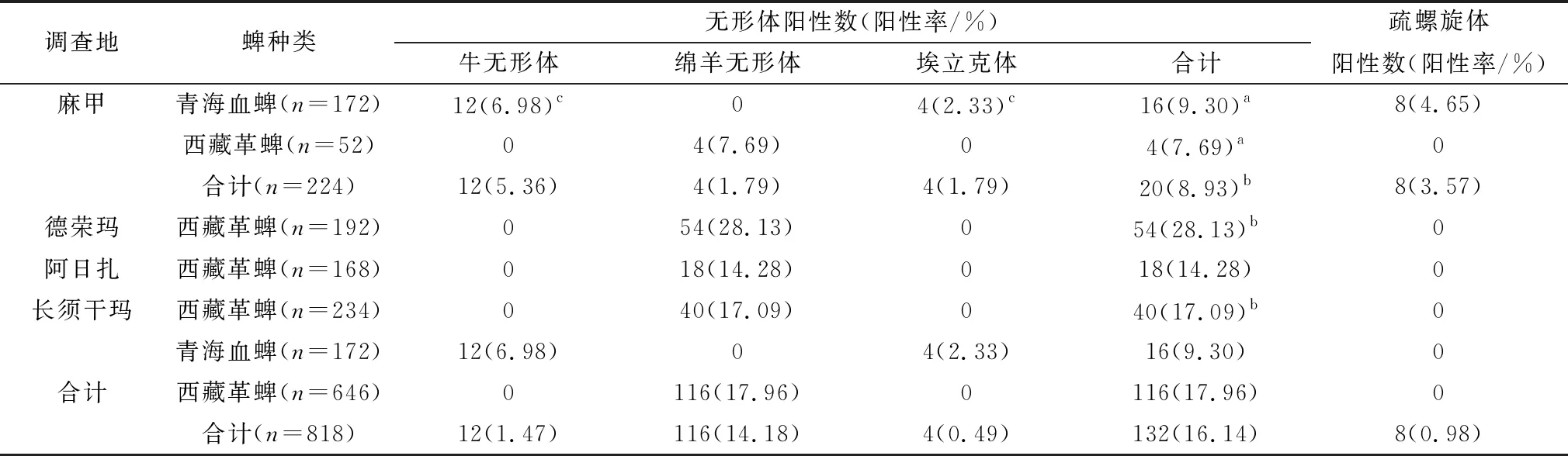
表2 石渠县蜱种类及携带无形体与疏螺旋体检测情况统计表
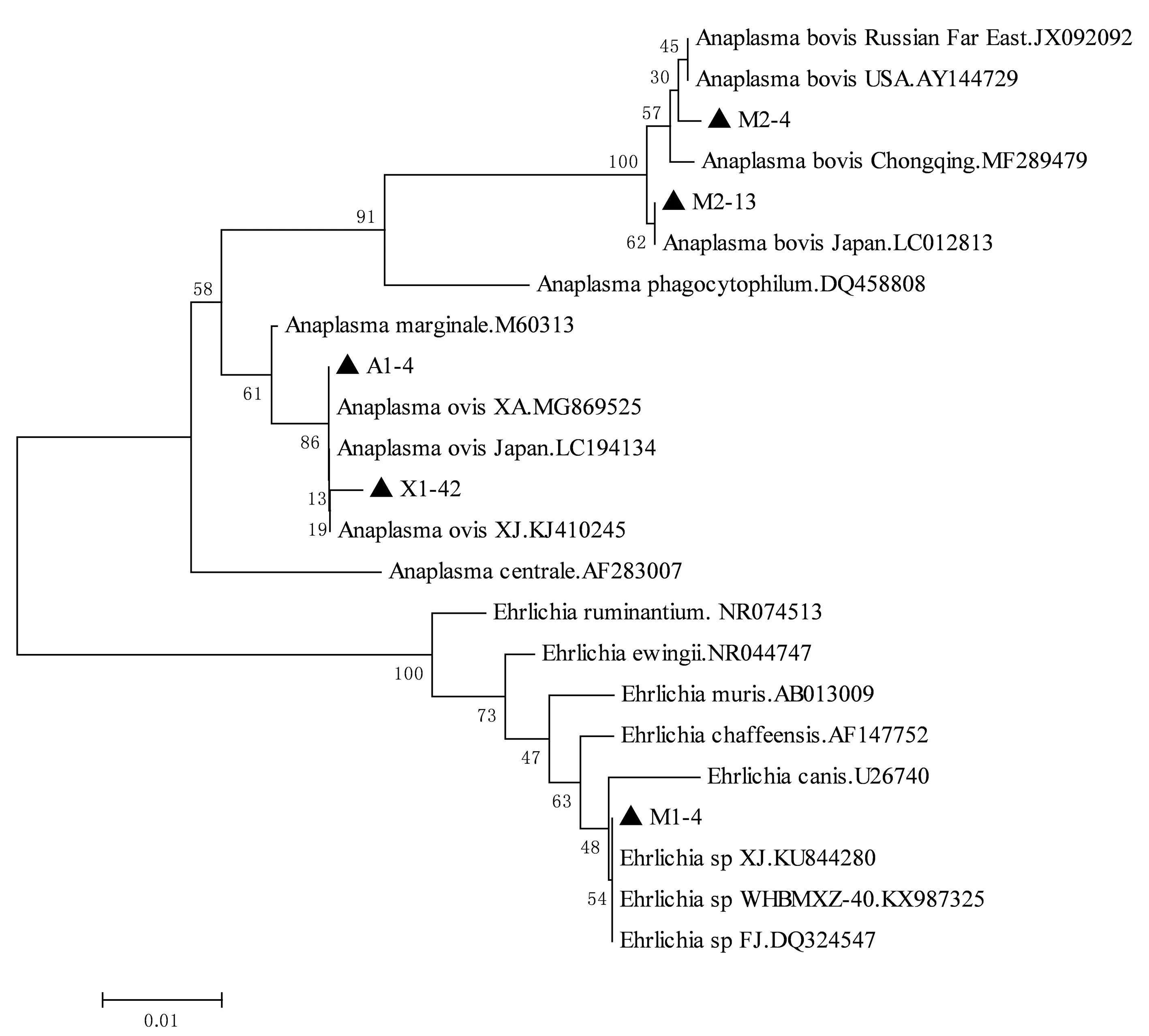
图1 基于无形体科16S rRNA基因片段构建系统进化树
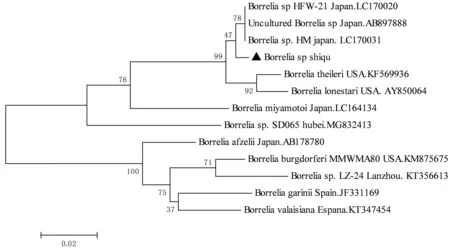
图2 基于疏螺旋体flaB基因片段构建系统进化树
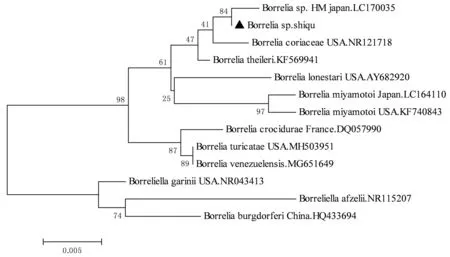
图3 基于疏螺旋体16SrRNA基因片段构建系统进化树
3 讨论
本研究首次对四川省石渠县牦牛体表寄生蜱进行无形体和疏螺旋体检测,结果显示检测到牛无形体、绵羊无形体、埃立克体和疏螺旋体。其中牛无形体和绵羊无形体是无形体属成员,另外无形体属还包括:边缘无形体(A.marginale)、吞噬细胞无形体(A.phagocytophilum)、中央无形体(A.centrale)、血小板无形体(A.platys)以及山羊无形体(A.capra)。在本研究中,经序列比对分析发现,青海血蜱中携带牛无形体和埃立克体,西藏革蜱中携带绵羊无形体,这可能与蜱的种类有关。许多研究表明不同蜱种能特异的携带不同无形体,如孟庆玲等[13]对古尔班通古特沙漠南缘硬蜱携带无形体进行了调查,结果表明不同蜱携带无形体的种类有所不同,残缘璃眼蜱中携带边缘无形体,长角血蜱中携带绵羊无形体;张群芝等[14]从豫南地区的长角血蜱中检测到了吞噬细胞无形体。另外,结果显示绵羊无形体处于优势地位,约占无形体总数的87%,这很可能是因为西藏革蜱是石渠县优势蜱。经报道,目前只有吞噬细胞无形体、山羊无形体和绵羊无形体变种可引起人无形体病[15-18],然而关于A.ovis能否感染人尚不明确。目前国内只有魏然等[19]进行绵羊无形体对人致病性研究,结果表明A.ovis可以感染人,提示石渠县牧民存在潜在感染A.ovis的风险,需要引起重视、并加强监测。此外,本研究选取16S rRNA 基因作为靶基因,该基因在国内外已被广泛的应用于无形体的鉴定与检测[4-5,20],并且能快速诊断多种无形体科病原,在本研究中检出牛无形、绵羊无形体和埃立克体,因此该基因可作为对未知无形体科病原大样本量初筛的首选基因。
目前,已证实4个基因型的疏螺旋体具有致病性,分别是埃氏疏螺旋体(B.afzelii)、伽氏疏螺旋体(B.garinii)、伯氏疏螺旋体狭义种(B.buredorferisensustricto)和法雷斯疏螺旋体(B.valaisana)[21]。另外,近年来在人的血清中检测到了宫本疏螺旋体(B.miyamotoi)[22],并且不断有关于宫本疏螺旋体感染人的病例报道[23]。本研究中,只在麻甲乡的青海血蜱中检测到疏螺旋体,这可能与蜱的种类有关,有研究表明,在大多数情况下,疏螺旋体属是由某些特定的蜱类传播[24],在我国北方全沟硬蜱是莱姆病的主要传播媒介,而南方林区粒形硬蜱和二棘血蜱是主要的传播媒介[21];也可能与调查点的生境有关,研究表明疏螺旋体主要分布在东北、华北和西北林区[21],而在本次调查的地点中只有麻甲乡境内有林区。基于疏螺旋体的flaB和16S rRNA基因分析表明,本研究获得的疏螺旋体与分离自日本大刺血蜱中Borreliasp.HM菌株最接近,与已知的疏螺旋体相比较,flaB片段与美国色勒疏螺旋体(Borreliatheileri)KAT株的同源性最高为96.45%,而16S rRNA片段与美国BorreliacoriaceaeCo53株的同源性最高为98.85%,并不能准确鉴定到种,这与KIWA等[24]在日本的研究结果相似,因此后期可基于多位点序列分型(MLST)对疏螺旋体进行进一步研究。
近年来,随着蜱传病对公共卫生的威胁越来越严重,不断有新的蜱传病复合感染的报道。如有无形体与巴贝斯虫混合感染、伯氏疏螺旋体与巴贝斯虫混合感染、森林脑炎病毒与伯氏疏螺旋体的混合感染、伯氏疏螺旋体与无形体的混合感染、伯氏疏螺旋体与无形体和巴贝斯虫3者之间的混合感染等[25]。同样在本研究中也存在混合感染的现象,其中有3只青海血蜱混合感染了牛无形体和疏螺旋体。另外,本实验室在2016年对同是高原地区的阿坝州牦牛和藏绵羊感染梨形虫的情况进行了调查,结果显示感染率较高,达49%[26]。因此,今后可进行蜱传梨形虫检测,从而进一步探讨其混合感染之间的关系。
综上所述,本研究首次从四川省石渠县牦牛体表的青海血蜱体内检测到牛无形体、埃立克体和疏螺旋体,西藏革蜱体内检测到绵羊无形体,这为了解此类病原体在当地的流行、传播以及制定科学的防控措施提供了数据支持。
