黄土高原中部针叶树与灌木径向生长对气候的响应差异
2020-09-17李晓琴张凌楠曾小敏王可逸王雅波路强强刘晓宏
李晓琴,张凌楠,曾小敏,王可逸,王雅波,路强强,2,刘晓宏,3,*
1 陕西师范大学地理科学与旅游学院, 西安 710119 2 陕西省西安植物园,陕西省植物研究所, 西安 710061 3 中国科学院西北生态环境资源研究院冰冻圈科学国家重点实验室, 兰州 730000
全球气候持续变暖导致严重干旱等极端气候事件发生的频率和强度急剧增加[1-3],将对森林生态系统的健康发展产生不利影响。干旱程度增加,会导致树木生长率下降,死亡率普遍上升[4]。了解并掌握干旱胁迫下树木径向生长特征是研究气候变化影响森林生态系统结构与生产力的基础。树木年轮是研究干旱事件对树木径向生生影响的良好代用资料[5-7]。研究表明,油松在黄土高原树木年轮气候学研究中具有重要意义。Cai等利用陕西中部黄龙山油松年轮宽度重建平均气温[8];Koretsune等利用安塞纸坊沟油松年轮宽度及δ13C分析季节性降雨量对水分利用效率及树木径向生长的影响[9];Song等利用黄土高原西部石门山油松树轮样品重建平均气温[10]。在干旱、半干旱区,灌木是贫瘠土壤上生长的主要植物。灌木能在长期土壤侵蚀形成的破碎地形和黄土峭壁等严酷的生态环境下生长,并形成茂密的单一植物群落[11-12],涵养土壤水分,减少水土流失,具有重要的生态作用。狼牙刺和黄刺玫是黄土高原中部主要灌木树种[13-14],灌木生长对环境的适应性特征研究,可为当地进行生态屏障科学建设提供有价值的信息[15]。近年来,黄土高原地区灌木树轮研究已逐步发展,如黄土高原西部红砂(Reaumuriasongarica)、柠条(Caraganakorshinskii)等的气候与生态学研究[16-17],人工物种柠条(Caraganakorshinskii)和自然物种荆条(Vitexnegundovar.heterophylla)径向生长对气候变化的响应差异[18],但对狼牙刺(Sophoraviciifolia)和黄刺玫(Rosaxanthina)径向生长对气候响应的研究尚有所欠缺。
黄土高原地区干旱导致的植物生长缺水是最突出的生态问题。本文研究区位于黄土丘陵区,是中国生态环境最脆弱的地区之一。该地光照强,降水量少,蒸散量大,年降水量与蒸散量极不平衡,水分亏缺严重,影响植物光合作用过程,制约植物生长与水分利用,从而限制形成层活动,使树木生长受到严重的水分胁迫[19-20]。
研究表明,即使树木能在特定的干旱时期存活,对干旱事件的敏感性也会发生变化,这种变化将导致干旱后持续数年的滞后效应[21-23],可能会推迟树木生长恢复,并增加树木面对未来干旱事件的脆弱性[24-26]。Anderegg等在全球范围的研究表明干旱后森林生长恢复需要2—4年时间[27]。若干旱事件发生频率高于恢复所需时间,将对森林生态系统发展及可持续产生重要影响[28]。Lloret等提出表征树木对极端干旱响应能力的指标:抵抗力(Resistance,Rt)、恢复力(Recovery,Rc)及恢复弹力(Resilience,Rs)[29],常用于比较不同树种应对干旱的能力。Zang等对比分析3种常见的欧洲温带森林树种挪威云杉(Piceaabies)、银杉(Abiesalba)和欧洲山毛榉(Fagussylvatica)的耐旱性差异[30];Pretzsch等对比分析德国南部欧洲山毛榉(Fagussylvatica)和无梗橡树(QuercuseaMatt)纯林和混交林干旱胁迫的抗性差异[31];Merlin等分别从林分组成和树木大小方面对比分析无柄橡树(Quercuspetraea)和苏格兰松(Pinussylvestris)抗旱能力的差异[32];Serra-Maluquer等对比分析伊比利亚半岛东北部三种针叶树黑松(Pinusnigra)、地中海白松(Pinushalepensis)及欧洲赤松(Pinussylvestris)在连续三次极端干旱事件中抗旱性差异[23]。
本文以采自黄土高原中部的油松、狼牙刺及黄刺玫年轮样品为研究对象,通过测定树轮宽度,结合气候资料,分析气候变化对树木径向生长的影响。基于帕默尔干旱指数(Palmer Drought Severity Index,PDSI)和年轮宽度,计算极端干旱条件下不同树种对干旱的抵抗及恢复能力,旨在揭示黄土高原中部不同树种在气候变化背景下应对干旱能力的差异。
1 材料与方法
1.1 研究区概况
研究区位于黄土高原中部陕西省延安市任家台林场(36.01°N—36.28°N,109.17°E—109.39°E)(图1),采样点海拔范围为1146—1267 m。研究区属大陆性暖温带季风气候,光照充足,四季分明,年平均气温10℃,最热月(7月)平均气温23℃,最冷月(1月)平均气温-5℃。1982—2017年平均年总降水量为532 mm,其中6—9月降水占全年65%以上。近40年来,研究区年均气温呈先上升后下降趋势,但总体上升趋势明显;年降水和相对湿度均有下降趋势,但并不明显(图2)。
研究区森林植被主要为退耕还林后恢复的天然次生林,主要树种有油松(Pinustabulaeformis)、辽东栎(Quercuswutaishansea)、山杨(Populusdavidiana)及白桦(Betulaplatyphylla),灌木主要有狼牙刺(Sophoraviciifolia)、黄刺玫(Rosaxanthina)、胡枝子(Lespedezabicolor)、沙棘(Hippophaerhamnoides)、柠条(Caraganakorshinskii)及铁杆蒿(TripoliumvulgareNees)等。

图1 树轮采样点示意图Fig.1 Tree ring sampling sites map
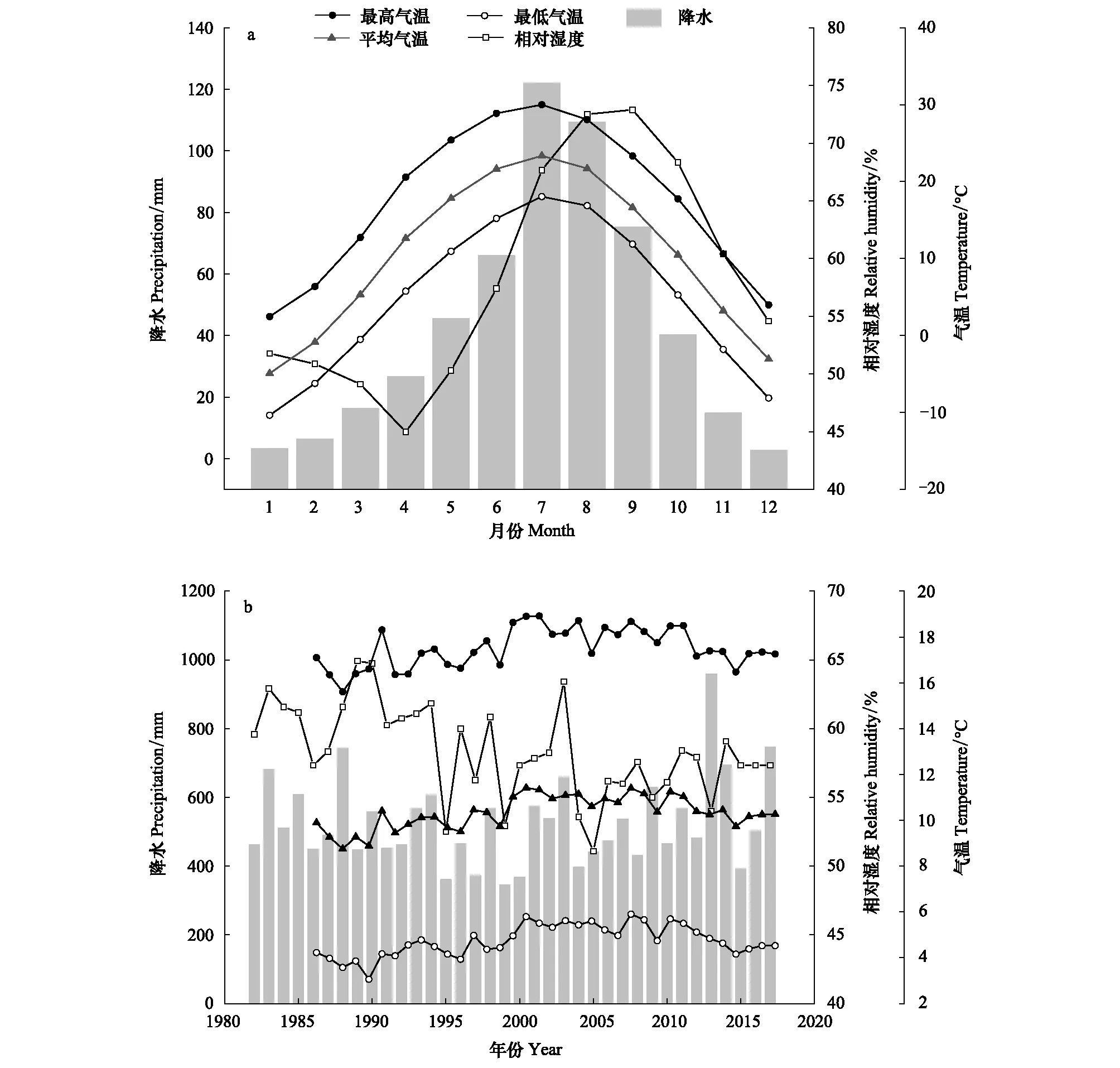
图2 延安气象站气象资料(1982—2017年)Fig.2 Meteorological data of Yan′an meteorological station (1982—2017)
1.2 样品采集及年表建立
2018年6月在延安任家台林场采集油松、狼牙刺及黄刺玫样品(图1和表1)。油松取样时,选择长势良好,树龄较大的优势木,用内径为5.15 mm的生长锥在胸径(1.3 m)处钻取完整的树轮样芯,每棵树钻取两个方向,共钻取51棵树,获得树芯102根。将钻取的树芯编号后装入纸管封存。选取生长健康粗壮的灌木单枝或主枝,沿茎基部截取树盘[33],每株灌木截取1—2个树盘,狼牙刺获得69个树盘,黄刺玫获得37个树盘。样品带回实验室后,油松树芯固定、风干,灌木用切割机修整成2—3 cm厚度平整的圆盘。分别用不同粗细的砂纸逐步打磨抛光,直至树芯和圆盘表面光滑,年轮界限明显,导管和薄壁细胞在显微镜下清晰可见。
为保证定年准确性,首先在显微镜下目视交叉定年,灌木样品沿4个方向对同一树盘进行初步定年[34]。利用0.01 mm精度的LINTAB 6.0轮宽分析仪测量树轮宽度,用COFECHA程序对测量和交叉定年结果进行校验[35],剔除不能正常交叉定年的序列,最后保留年轮清晰且序列较长的油松树芯64根,狼牙刺树盘45个,黄刺玫树盘28个(表1)。利用ARSTAN程序,对原始年轮序列采用适于干旱与半干旱区的负指数函数法拟合树木生长趋势[36],用双权重平均法合并后得到标准年表(STD)。采用子样本信号强度SSS>0.85的样本量作为年表起始点,高于样本量的年表序列具有可靠性[37],油松、狼牙刺和黄刺玫年表的可靠区间分别为1982—2017年、1986—2017年和2002—2017年。根据以下公式计算树轮断面积增量(BAI):
式中,d表示树木半径(未标准化的原始宽度),n表示年份[38]。
1.3 气候资料及数据分析
1982—2017年延安气象站(36.36°E,109.32°N,海拔1090 m,图1)气候数据来自中国气象数据网(http://data.cma.cn/),包括最高气温、平均气温、最低气温、降水量及相对湿度。帕默尔干旱指数(PDSI)数据来自荷兰皇家气象研究所数据共享网(https://climexp.knmi.nl),分辨率0.5°×0.5°,范围为35.75°—36.25°N,108.75°—109.25°E,计算范围内PDSI算术平均值用于研究。
采用不同树种年表与气候的最大公共时段进行分析,油松为1982—2017年,狼牙刺为1986—2017年,黄刺玫为2002—2017年。考虑到树木生长对气候因子响应的“滞后现象”[39],选择前一年6月到当年10月的气候因子与标准年表进行Pearson相关分析,揭示限制3种树木径向生长的主要气候因子。
1.4 树木径向生长对极端干旱的抵抗力、恢复力及恢复弹力分析
计算树木径向生长对极端干旱响应的3个指标:抵抗力(Rt)、恢复力(Rc)和恢复弹力(Rs)[30]。Rt为干旱与干旱发生前树木生长的差异,表征树木在干旱胁迫后保持生长的能力;Rc为干旱与干旱发生后树木生长的差异,表征树木在干旱胁迫后恢复生长的能力;Rs为干旱发生前后树木生长的差异,表征树木在干旱发生后恢复到干旱发生前生长水平的能力[40]。3个指标计算公式如下:
Rt=Gd/Gprev
(1)
Rc=Gpost/Gd
(2)
Rs=Gpost/Gprev
(3)
式中,Gd表示干旱事件发生年的树轮宽度,Gprev和Gpost分别表示干旱事件发生前3年和后3年的树轮宽度的算术平均值[41]。
Rt量化干旱前期至干旱期树木生长的差异,Rt=1表示完全抵抗,干旱胁迫未影响树木生长,Rt>1表征树木承受干旱胁迫后加速生长,Rt<1表征树木遭受干旱胁迫后生长下降。Rc描述干旱期后恢复生长的反应,Rc=1表明干旱期后低生长水平仍存在,Rc<1表明生长水平进一步下降,Rc>1表明干旱后生长水平恢复。Rs表示干旱前后的差异,Rs<1表示生长下降和低恢复率,Rs>1表示生长增加和高恢复率[31]。
为分析极端干旱对树木径向生长的影响,将上年10月至当年9月平均PDSI<-2的年份设定为干旱年[42]。干旱有多种特征,如干旱频率和干旱强度[43-44]。干旱频率为年内干旱月数所占比率,干旱强度为等于或低于干旱阈值的累积与年内连续干旱月数的比值。
2 结果与分析
2.1 树轮年表统计特征
表1表明,狼牙刺、黄刺玫平均敏感度(MS)高于油松,两种灌木比油松对气候更敏感;狼牙刺标准差(SD)最大,其包含的气候信息最多;3个年表信噪比(SNR)较高,样本具有丰富的环境信息;样本总体代表性(EPS)>0.85,样本所含信号基本能代表总体特征。总体而言,研究区3个树种年表统计特征包含较多的气候信号,采集样品符合年轮学研究要求。标准年表显示(图3),油松、狼牙刺与黄刺玫在高低频变化上较为一致,特别是2000与2011年,3个树种均表现为明显的轮宽窄年。
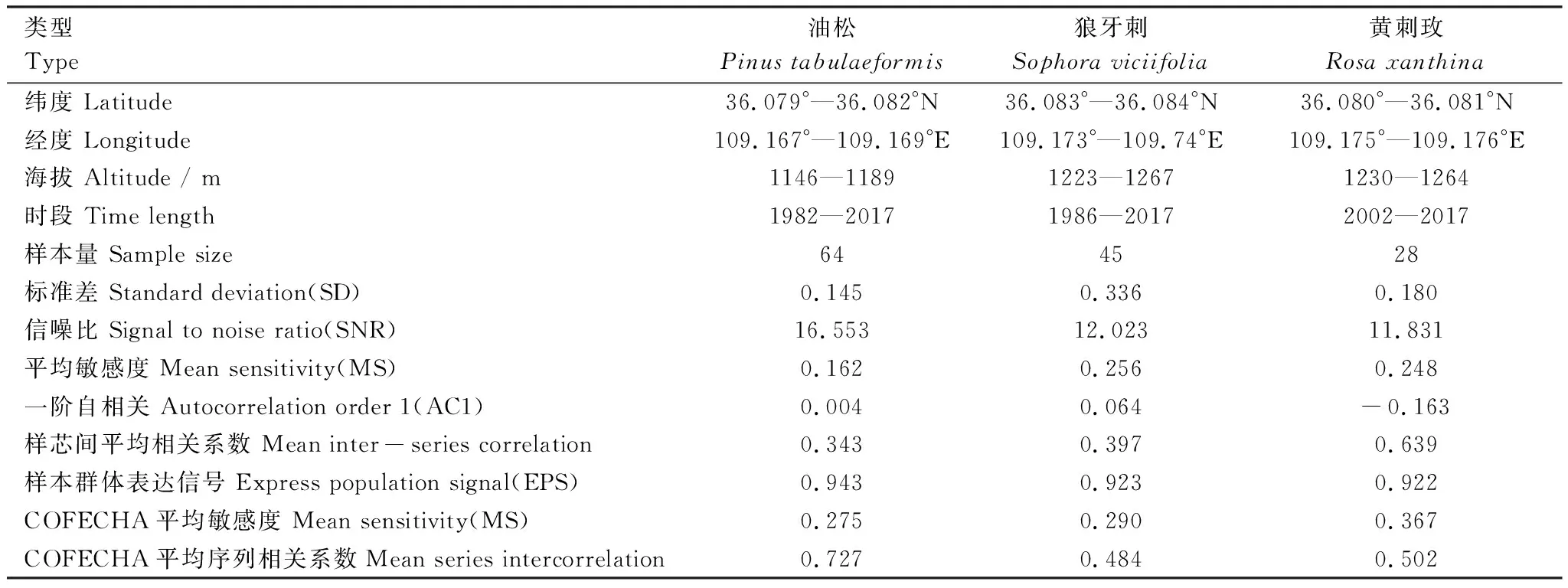
表1 样地信息及树轮宽度特征
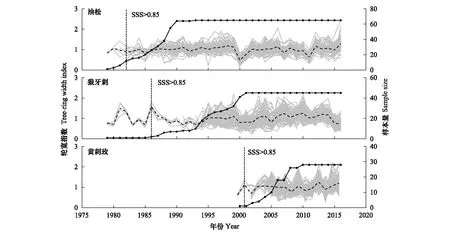
图3 3个树种标准年表(虚线),单根树芯的标准年表(灰线)及样本量(点线)Fig.3 Site standard chronologies (the dotted lines), single standard chronologies (the gray lines) and sample depths (the scattered lines) of three tree speciesSSS为子样本信号强度
2.2 树木径向生长对气候的响应
图4表明,油松与前一年9月平均最低气温,当年5月平均最高气温和平均气温显著负相关(r介于-0.417—-0.366,P<0.05),与当年9月平均最高气温显著正相关(r=0.332,P<0.05);狼牙刺与前一年9月平均气温,当年7月平均最高气温,8月平均气温和平均最高气温显著负相关(r介于-0.372—-0.358,P<0.05);黄刺玫与当年6月平均最低气温,7月平均气温和平均最高气温显著负相关(r介于-0.534—-0.486,P<0.05)。
油松与前一年11月降水和当年7月相对湿度呈显著正相关(r=0.329,r=0.38,P<0.05),与PDSI均正相关,且在当年4—7月达到显著水平(r介于0.348—0.373,P<0.05);狼牙刺与PDSI均正相关,且在前一年7—8月、12月和当年3—7月达到显著水平(r介于0.353—0.429,P<0.05);黄刺玫与当年7月PDSI正相关(r=0.501,P<0.05)。
油松和狼牙刺均受前一年和当年生长季气温限制,而黄刺玫主要受当年生长季气温限制,生长季相对湿润的环境有利于油松生长。3个树种均与PDSI正相关,油松受当年生长季干旱胁迫,狼牙刺受前一年和当年生长季干旱胁迫,黄刺玫只在当年7月表现出受到明显的干旱胁迫。
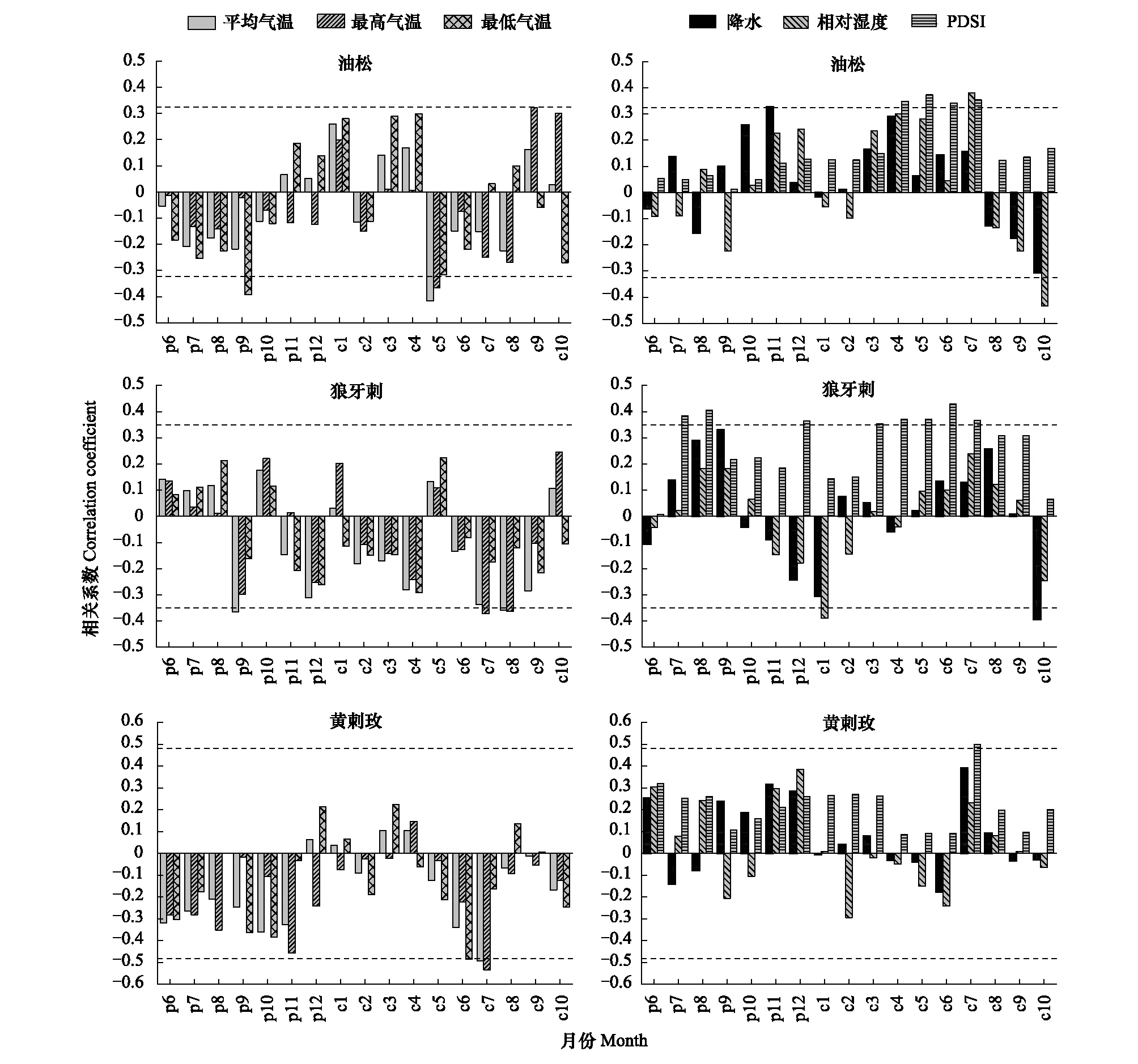
图4 3个树种标准年表与气候因子的相关系数Fig.4 Correlation coefficients between the standard chronologies of three tree species and the climate factors虚线代表显著性水平(P<0.05),p表示前一年(previous),c表示当年(current),帕默尔干旱指数(Palmer Drought Severity Index, PDSI)
2.3 径向生长对极端干旱的响应
根据3个树种BAI,结合前一年10月至当年9月平均PDSI<-2选出1995年、1999—2001年、2005年和2011年4个干旱事件[29](图5)。

图5 3个树种BAI(树轮断面积增量),阴影表示标准差;前一年10月至当年9月平均PDSI(1982—2017年)Fig.5 BAI (Basal Area Increment) of three tree species, shadows denote standard deviation; mean PDSI from October of the previous year to September of the current year (1982—2017)
图6显示,1995年55%的油松及79%的狼牙刺样本Rt>1,干旱并未直接影响树木生长;1999—2001年干旱期,78%的油松及66%的狼牙刺样本Rt<1,严重干旱导致树木生长下降;除2005年外,狼牙刺Rt均值大于油松,平均水平上狼牙刺抵御干旱的能力强于油松。除2005年黄刺玫外,其余干旱事件3个树种Rc>1,干旱后树木生长均有一定程度恢复,特别是油松在2011年干旱后完全恢复生长。从1995年到2011年,油松和黄刺玫Rc均有上升趋势。1995年干旱后71%的油松及67%的狼牙刺样本、2005年干旱发生后70%的狼牙刺样本、2011年干旱后64%的油松及66%的黄刺玫样本Rs>1,干旱后树木生长增加并伴随高恢复率。油松和狼牙刺在1999—2001年干旱后Rs均值<1,树木未恢复到干旱前的生长水平[23]。油松和黄刺玫在2011年干旱后Rs均值>1,表明油松在干旱胁迫后,很大程度上能恢复到干旱前的生长水平,黄刺玫对干旱胁迫的恢复弹性较强。

图6 3个树种在不同干旱事件的抵抗力、恢复力和恢复弹力Fig.6 Resistance、Recovery and Resilience of three tree species in different drought events经配对t检验,小写字母差异具有统计学意义(P<0.05)
图7表明,月平均PDSI较小时,3个树种Rt较小,1999—2001年PDSI=-3.38,油松和狼牙刺Rt、Rs<1;1995年PDSI=-2.46,油松和狼牙刺Rt、Rs>1。月均PDSI减小,油松和黄刺玫Rc增强。干旱频率增加,油松Rc、Rs相对减弱。2005年干旱频率达到100%,黄刺玫Rc、Rs<1,狼牙刺Rt<1。干旱强度减小,3个树种Rt均较小,油松和黄刺玫Rc均较强,狼牙刺Rc较弱,干旱强度对3个树种Rs无明显影响。
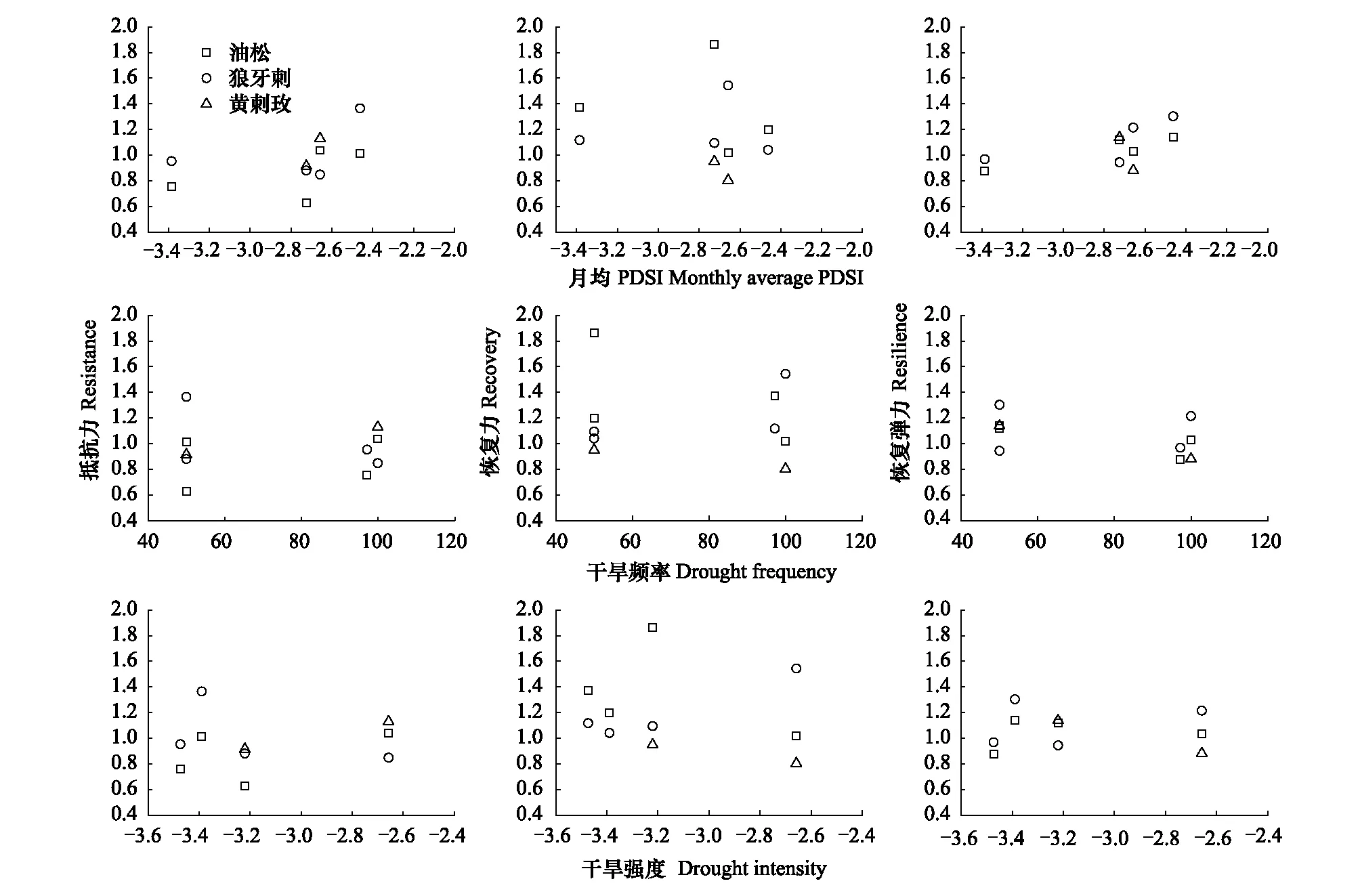
图7 3个树种抵抗力、恢复力及恢复弹力与月平均PDSI、干旱频率及干旱强度的关系Fig.7 Relationships between monthly mean PDSI, drought intensity, drought frequency and Resistance、Recovery、Resilience of three tree species
3 讨论3.1不同树种径向生长对气候响应的差异
研究表明,3个树种径向生长均受到生长季气温的显著影响。油松与前一年9月和当年5月气温显著负相关,与当年9月气温显著正相关;狼牙刺与前一年9月和当年7—8月气温显著负相关,黄刺玫与当年6—7月气温显著负相关。温度过高造成植物蒸腾作用和土壤蒸散发加强,土壤可利用水减少,叶面气孔部分关闭,光合作用减弱,进而合成有机物减少,但呼吸作用仍不断消耗供树木生长的营养物质,导致树木生长缓慢[45-47]。9月处于生长季末期,温度降低,树木尚未休眠,温度较高有利于光合产物积累[46],对油松当年和下一年生长都有促进作用。
3个树种标准年表与降水相关分析表明,上一年冬季较好的水分条件增加光合产物积累,促进油松下一年早材细胞生长。当年7月相对湿度较高,树木蒸腾作用减弱,土壤中充足的水分有利于油松生长。PDSI反映土壤水分的可利用程度[48],土壤可利用水直接影响形成层活动,进而影响木质部形成[49]。3个树种径向生长均与PDSI正相关,说明其均受到干旱制约,这与梁尔源[50],陈峰[51]和彭剑峰[52]等的结果一致。
3.2 不同树种径向生长对极端干旱的响应差异
持续的气候变化导致极端干旱频率和强度加剧,树木应对干旱的恢复能力下降[44]。在极端干旱期,3个树种均出现明显的生长下降,这与前人的研究结果一致[24,53]。对4个干旱事件逐月的情况进行对比,发现1995年4—9月干旱;1999、2000和2005年全年干旱,且强度较大;2001年除当年9月外均干旱;2011年3—8月干旱。1995年夏季干旱对油松和狼牙刺生长影响较大,夏季干旱阻碍油松针叶形成;狼牙刺作为环孔材树种,在遭遇干旱后易发生空穴和栓塞,限制水分传输[24]。1999—2001年、2005年强干旱抑制油松生长,干旱可能导致木质部管胞气穴化,影响油松抵御干旱的能力[32]。2005年全年干旱抑制黄刺玫生长,可能导致木质部栓塞,致使恢复能力减弱。2011年夏旱消退后,水分供给改善,油松针叶在秋冬季仍能保持光合作用,能在干旱后恢复生长[54]。
水分是树木生长和存活的基础,水分传输在根部受到影响,水分向上传输的有效性降低,植物的水分平衡、光合作用、气孔调节等生理代谢过程将受到影响。狼牙刺具有发达的根系,根深可达到200cm以下的土层,为深根性树种[55]。黄刺玫基本没有垂直伸展的主根,但具有发达的水平根和大量斜生根[56]。有研究表明狼牙刺叶片比黄刺玫有更强的抗旱性[57]。
木本植物在持续干旱期的生存和恢复能力与其栓塞抗性密切相关[58-59],抵抗栓塞形成的能力与木质部结构有一定关联[60-61]。油松的木质部只有管胞,管胞分子小且有横壁,输水阻力大速度慢,但木质部脆弱性小,不易发生气穴和栓塞,在受到水分胁迫时仍能维持较高的导水率[62]。一般而言,抗旱性强的植物具有小的导管直径及大的木材密度[63]。导管直径和木材密度是影响黄土丘陵区典型灌木抵抗气穴化的重要因素[64],导管直径越大,抵抗气穴化的能力越弱[65],木材密度和抵抗栓塞形成能力呈显著正相关[66]。研究表明狼牙刺导管直径及木材密度均大于黄刺玫,且狼牙刺和黄刺玫抵抗气穴化和栓塞形成的能力较强[66],但两者的比较还需进一步探究。
在干旱发生后,油松恢复能力比狼牙刺、黄刺玫更强,这是因为油松有更耐旱的木质部,更复杂的群落生态系统和强大的根部系统。油松具有较好的郁闭度,腐殖质的积累增加了水的储存和营养物质的供应,从而提高用水效率[67]。两种阔叶灌木具有较大的叶片来获取资源,可以通过减少叶片的蒸腾作用来维持生长[68]。
4 结论
黄土高原中部油松、狼牙刺及黄刺玫径向生长均受到生长季温度制约,油松生长得益于前一年冬季充足的降水储备和生长季相对湿润的环境。3个树种均受到明显的干旱胁迫,随着干旱的频率和强度增加,干旱事件对树木生长的影响加剧,致使树木在干旱后恢复能力减弱。相比两种灌木,油松对干旱有更强的恢复能力。狼牙刺比黄刺玫有更耐旱的叶片及更有利的根部系统,抵御干旱的能力更强,但狼牙刺在遭受干旱后恢复弹力比黄刺玫弱,这可能跟两者抗栓塞能力的差异有关。进一步研究还需结合气候资料与植物生理调控特征对干旱胁迫的综合响应机制,阐明不同树种在气候变化背景下应对干旱的能力差异,为黄土高原中部森林的可持续发展提供科学依据。
致谢:感谢任家台林场工作人员在树轮样品采集中提供的帮助。
