三江源区退化高寒草甸蒸散特征及冻融变化对其的影响
2020-09-17田晓晖张立锋陈之光唐艳鸿
田晓晖,张立锋,张 翔,陈之光,赵 亮,李 奇,唐艳鸿,古 松,*
1 南开大学生命科学学院,天津 300071 2 中南林业科技大学生命科学与技术学院,长沙 410004 3 中国科学院西北高原生物研究所,西宁 810008 4 北京大学城市与环境学院,北京 100871
陆地生态系统蒸散(ET)是全球水量平衡的主要分量,也是生态系统水分消耗的主要方式,主要由土壤蒸发和植被蒸腾组成[1]。由于ET在土壤—植被—大气系统中的重要纽带作用,在全球变化科学研究中受到高度关注[2]。有研究表明,地表降水大约有70%通过蒸散返回大气,在干旱区甚至达到95%以上[3]。ET变化受辐射、降水、温度、植被等因子影响;反之,ET变化又影响生态系统植被生长状况和水分收支等。水热运动相互耦合,蒸散影响水分收支的同时,也会影响生态系统的能量(潜热和显热能等)分配,从而对区域气候产生一定影响[4]。草地是陆地生态系统中分布最广泛的植被类型之一,约占陆地生态系统总面积的1/3[5]。然而,草地蒸散的年际之间存在较大的变化,近年来受到了科学界的广泛关注[4,6]。因此准确掌握草地生态系统的蒸散变化及环境因子对其的影响,对于更好地理解全球水分收支动态及气候变化具有重要意义。
青藏高原平均海拔超过4000 m,面积约250万km2,被誉为“世界第三极”,由于其独特的地理和气候条件,成为全球气候变化的敏感区和放大区[7]。草地是其最主要的植被,占青藏高原面积的70%,其中高寒草甸分布最广,支撑着青藏高原地区的畜牧业发展[8]。由于青藏高原特殊的地理位置,高原上广泛分布着多年冻土和季节冻土,近地表层土壤的季节冻融循环影响着高原地表能量和水分交换[9]。近几十年来,在全球气候变暖的背景下,高原的温度呈明显的上升趋势,有研究表明,1960年以来,青藏高原气温以0.2—0.3℃/10 a的速度增加,而冬季增温的幅度(0.3—0.5℃/10 a)远远高于全年平均增温幅度[10]。不断加剧的气候变暖正改变冻土冻融发生过程和状态,总体上呈现最大冻土深度降低,冻结起始日期推迟,融化起始日期提前及冻结持续期缩短等的趋势[11-12]。全球气候变暖改变了青藏高原土壤冻融状态,这必将引起生态系统水分交换的变化。
三江源区位于青藏高原腹地,是长江、黄河、澜沧江的发源地,素有“中华水塔”之称,在水源涵养、维持生物多样性以及气候调节等方面发挥着重要作用,成为我国乃至世界上影响力最大的生态调节区之一[13]。然而,在气候变化和超载放牧的双重压力下,三江源区高寒草甸出现了严重的退化现象,部分地区形成了“黑土滩”,极大地降低了区域的水源涵养功能,进而严重影响到三江源区生态系统的结构和功能,改变了区域的水分收支状况,同时对三江源区的畜牧业乃至长江、黄河中下游地区的社会经济发展构成巨大的威胁[13-14]。虽然关于青藏高原高寒草地的蒸散研究也有一些报道[6-7,15],但由于高原面积广阔,地理环境差异较大,结果不尽相同。三江源区是我国重要的水源涵养地,也是典型的季节冻土区,然而对该区域高寒草甸蒸散变化的详尽研究仍然相对匮乏,更缺少冻融作用对其生态系统蒸散影响的相关报道。因此,定量研究三江源区退化高寒草甸蒸散变化及其冻融作用对其的影响对深入研究三江源区水分收支变化具有重要现实意义。
涡度相关技术是通过测定和计算CO2和H2O等物理量脉动与垂直风速脉动的协方差求算湍流通量的方法,目前已成为直接测定地表下垫面与大气间气体通量交换的理想方法,广泛应用于陆地生态系统的蒸散研究[1]。本研究利用涡度相关技术和微气象系统对三江源区退化高寒草甸的水汽通量及相关环境因子进行了连续观测,对2016年和2017年获取的观测数据进行统计分析,其主要目的是:(1)揭示三江源区退化高寒草甸生态系统蒸散的季节变化特征及年际差异;(2)探讨分析土壤冻融交替变化对蒸散的影响;(3)阐明蒸散变化对环境因子的响应。以期为全面探讨三江源区退化草甸蒸散对气候变化的响应及其生态系统水分收支变化提供参考依据。
1 研究地概况和研究方法
1.1 研究地概况
本试验地位于青海省果洛州玛沁县大武镇东南部的退化高寒草甸(34° 21′ N, 100° 29′ E),海拔3958 m。研究区地势平坦,草地分布均匀,该研究站代表了三江源区高寒草地典型植被类型[16]。该区属于典型的高原大陆性气候,无四季之分,仅有冷暖季之别,冷季漫长,干燥而寒冷,暖季短暂,湿润而凉爽;温度年差较小而日差较大,研究地年平均气温为-0.2℃,最冷月1月平均气温为-12.3℃,最热月7月的平均气温为10.1℃,全年无绝对无霜期。该区日照充足,太阳辐射强烈,年总辐射量为5500—6800 MJ/m2;年降水量为420—560 mm,85%集中在5—9月。该试验地土壤类型以高山草甸土为主,建群种为矮蒿草(Kobresiahumilis),主要伴生种为小蒿草(Kobresiapygmaea)、垂穗披碱草(Elymusnutans)、早熟禾(Poaannua)、细叶亚菊(Ajaniatenuifolia)等[17]。根据之前对该研究地植被类型、盖度等方面调查研究证实其处于退化状态[18]。退化高寒草甸于4月底或5月初返青;地上生物量从5月开始增加,7月底至8月初达到最大,10月植物开始衰老。
1.2 观测方法
涡度相关和微气象观测系统安装在地势平坦、视野开阔的退化高寒草甸上,“风浪区”(半径大于300 m)满足涡度相关观测要求。观测要素主要包括辐射通量、显热和潜热通量、土壤热通量、空气温湿度、不同深度土壤温度、不同深度土壤含水量、降水量等,观测仪器及安装高度见表1。涡度相关系统采样频率为10 Hz,所有观测数据每15 min输出一组平均值,并储存在CR5000和CR23X (CSI, USA) 数据记录仪。
本研究于2016年6—8月和2017年6、7、9月每月中旬对植被地上部生物量(above-ground biomass, AGB)和叶面积指数(LAI)进行调查。生物量测定采取收割法,随机选择5个大小为50 cm×50 cm的样方,将每一样方内绿色植物齐地面剪下后放进取样袋内并编号。之后将样品带回实验室,放入65℃恒温烘箱中72 h(烘干至恒重)称重(g/m2)。同时选取5个大小为25 cm×25 cm的样方,齐地面剪下地上部植物,用叶面积仪(Li- 3000, Li-Cor)测定植物叶面积。上述5个重复的平均值用于本研究的数据分析。
1.3 数据分析
首先对涡度相关系统观测的原始数据进行坐标轴旋转,温度变换和WPL校正,此外,剔除由于降水、仪器故障等引起的错误数据以及摩擦风较弱的数据(摩擦风速u*<0.1 m/s)。由地表反照率的变化可知,该研究地偶有降雪发生(主要发生在11月至翌年3月),但降雪频次和降雪量均较少且很快消融,由于蒸散量主要集中在生长季,因此对研究结果不会产生实质性影响,本研究并未进行特别分析。对于缺失及剔除的异常数据,按以下方法进行插补,当缺失的通量数据时间小于2 h时,使用线性内插法进行插补,而大于2 h的数据缺口则使用非线性回归法进行插补[19-20]。最后采用Excel和SPSS等软件进行统计分析。

表1 观测要素及其仪器
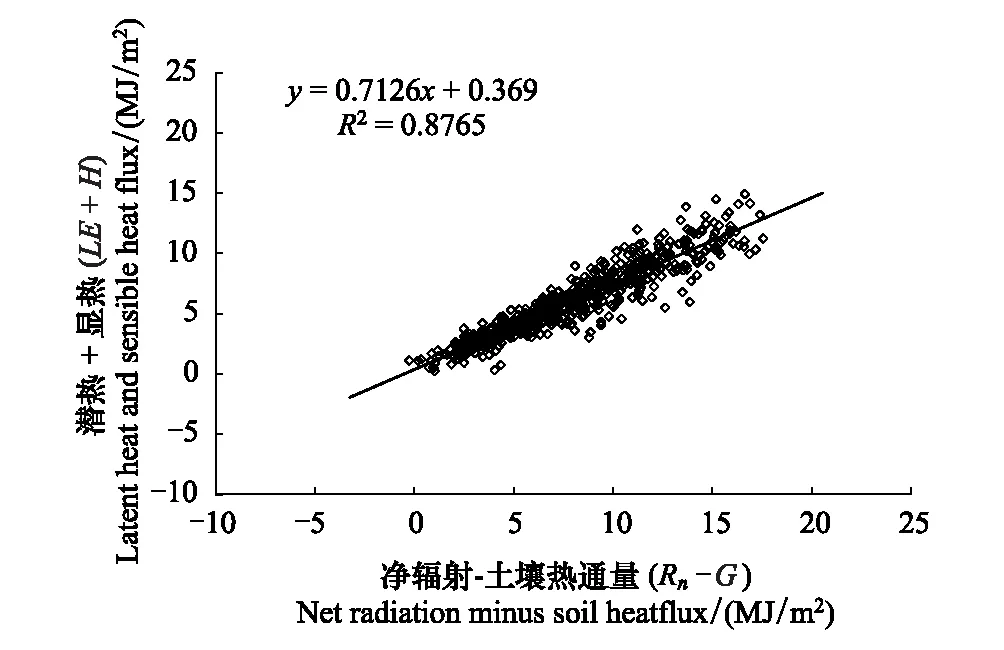
图1 退化高寒草甸涡度相关测定的显热与潜热通量之和(H + LE)与能量平衡法测定的有效能(Rn - G)的关系 Fig.1 The relationship between the sum of sensible heat and latent heat fluxes (H + LE) by eddy covariance and the available energy (Rn - G) by energy balance method in degraded meadow
生态系统能量平衡通常用下列公式来表示[20]:
Rn-G=H+LE
式中,Rn为净辐射通量(W/m2);H为显热通量(W/m2);LE为潜热通量(W/m2);G为土壤热通量(W/m2)。
能量闭合度是评价涡度相关技术的方法之一,通常用(H+LE)与(Rn-G)进行线性回归,利用其斜率和截距来分析能量的闭合度[20]。由图1可知,本研究能量闭合度约为0.71,说明存在能量不闭合现象,但该值在已报道的0.55-0.99范围内[21]。能量不闭合现象在涡度相关观测中普遍存在,由于导致能量不闭合的原因很复杂,详细原因有待于深入研究。
波文比(β)定义为显热通量与潜热通量的比值[22],即:
β=H/LE
冠层导度(gc)计算公式为[23]:
1/gc=ρCPVPD/(γLE)+(βΔ/γ-1)/ga
解耦系数(Ω)计算公式为[24]:
Ω=(Δ+γ)/(Δ+γ(1+ga/gc))
式中,ρ为空气密度(kg/m3),Cp为空气定压比热(MJ/kg℃),VPD为饱和水汽压差(kPa),Δ饱和水汽压—空气温度曲线的斜率,γ干湿表常数,ga为空气导度(mm/s),ga按下列公式计算[23]:
1/ga=μ/μ*2+6.2μ*-0.67
式中,u为风速,u*为摩擦风速(m/s)
平衡蒸散(ETeq)由下式计算得出[25]:
ETeq=Δ(Rn-G)/L(Δ+γ)
式中,L为水的汽化潜热系数(2.501 MJ/kg)。
2 结果与分析
2.1 环境因子季节变化
2016与2017年的Rn季节变化趋势相同(图2),最高值分别为12.81(6月)和11.22 MJ m-2d-1(7月),最低值出现在12月左右。但各月的Rn值略有差异,生长季中,2016年生态系统接收的Rn略高于2017年,分别为1654.88 MJ/m2和1546.71 MJ/m2,均超过全年Rn的60%。
两年气温(Ta)和5 cm深度的土壤温度(Ts)变化趋势与Rn相似(图2),最高值出现在7—8月,最低值均出现在1月。2016和2017年Ta的平均值分别为-0.35℃和-0.41℃,月均最高值分别出现在8月(11.0℃)和7月(9.7℃),而生长季中,2016年Ta的平均值为7.2℃,高于2017年平均值6.8℃。2016和2017年Ts的平均值分别为2.5℃和3.8℃,月均最高值分别出现在8月(12.9℃)和7月(13.4℃),而生长季中,2016年Ts的平均值为9.4℃,低于2017年的10.6℃。总体而言,两年的Ts高于Ta,且2016年的Ta略高于2017年,而2017年Ts高于2016年。
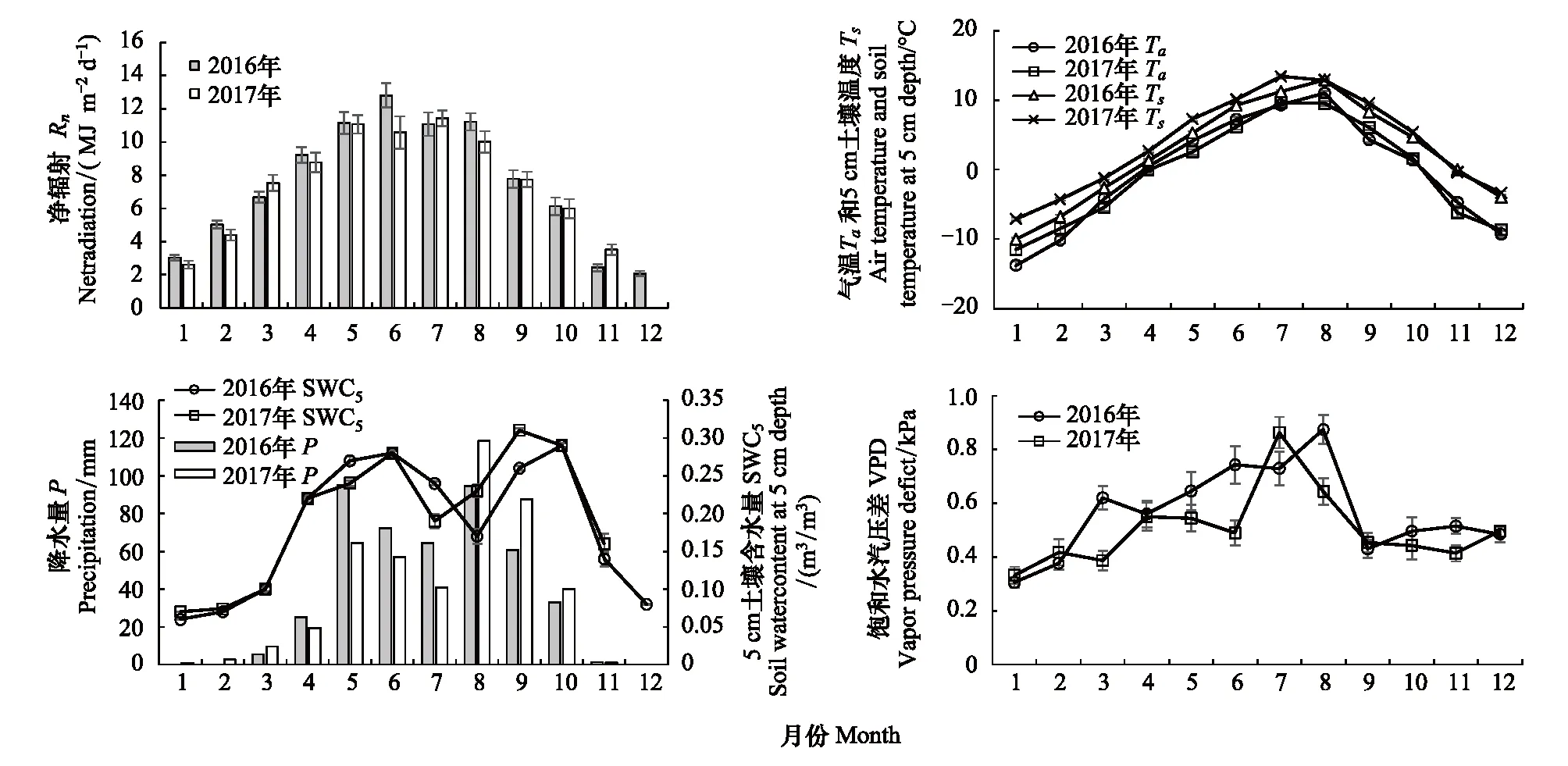
图2 2016和2017年退化高寒草甸的净辐射(Rn)、降水量(P)、5 cm土壤含水量(SWC5)、空气温度(Ta)、5 cm土壤温度(Ts)和饱和水汽压差(VPD)年变化Fig.2 Annual variations of net radiation (Rn), precipitation (P), soil water content at 5 cm depth (SWC5), air temperature (Ta), soil temperature at 5 cm depth (Ts) and vapor pressure deficit (VPD) in degraded meadow for 2016 and 2017
2016和2017年降水量相近(图2),分别为451.8 mm和442.3 mm,且降水主要集中在生长季5—9月,前者为386.7 mm,略高于后者的368.4 mm,分别占全年降水量的85.6%和83.3%。然而,两年生长季中月降水量分配存在较大年际差异,2016年降水量的最高值出现在5月(94.9 mm),其次为8月(94.7 mm),6、7、9月的降水量在60—70 mm之间;而2017年降水主要集中在8月(118.4 mm)和9月(87.7 mm),5—7月降水量相对较少,生长旺季的7月降水量仅为40.9 mm。
5 cm深度土壤含水量(SWC5)的季节变化与降水量密切相关(图2),最高值出现在降水较多的生长季,然而,由于该时期的ET较高,进而导致SWC5在降水相对较多的7—8月有所下降,类似结果在青藏高原其他草地研究中也有报道[5,28]。生长季中,2016和2017年SWC5的月最低值分别出现在生长旺季的8月(0.17 m3/m3)和7月(0.19 m3/m3),最高值分别出现在6月(0.28 m3/m3)和9月(0.31 m3/m3),且2016年生长季的SWC5平均值(0.24 m3/m3)略低于2017年(0.25 m3/m3)。总体而言,2016年5—7月的SWC5高于2017年同期,而8—9月却低于2017年同期的SWC5,这与两年的降水量变化基本一致。
饱和水汽压差(VPD)是表征空气湿度的物理量,其变化受降水、辐射和气温等因子影响。由VPD的季节变化可知(图2),总体上生长季的VPD高于非生长季,但各月的VPD值出现明显的年际差异。2016年的月均最高值出现在8月(0.87 kPa),而2017年月均最高值出现在7月(0.86 kPa)。生长季中,2016年VPD的平均值为0.69 kPa,明显高于2017的0.62 kPa,而非生长季中的平均VPD分别为0.48和0.42 kPa,总体上2016年VPD高于2017年。
2.2 蒸散季节变化
2016和2017年日蒸散量的年变化趋势基本一致,最高值均出现在植物生长旺季的7—8月,最低值出现在土壤冻结的冬季(图3)。土壤蒸发从3月末或4月初的冻融交替开始逐渐增加,之后随着土壤的解冻、降水的增加和植物的生长,蒸散(ET)迅速增加。2016和2017年月蒸散总量最大值分别为71.8 mm(8月)和92.5 mm(7月),之后随着辐射和温度的降低、植物生长的结束,ET逐渐降低(图3)。2016和2017年的ET分别为428.6 mm和485.6 mm,而生长季中的ET分别为314.4 mm和350.7 mm,分别占到全年总蒸散量的73%和72%,2017年蒸散量高于2016年,尤其是2017年7—10月ET明显高于2016年同期。
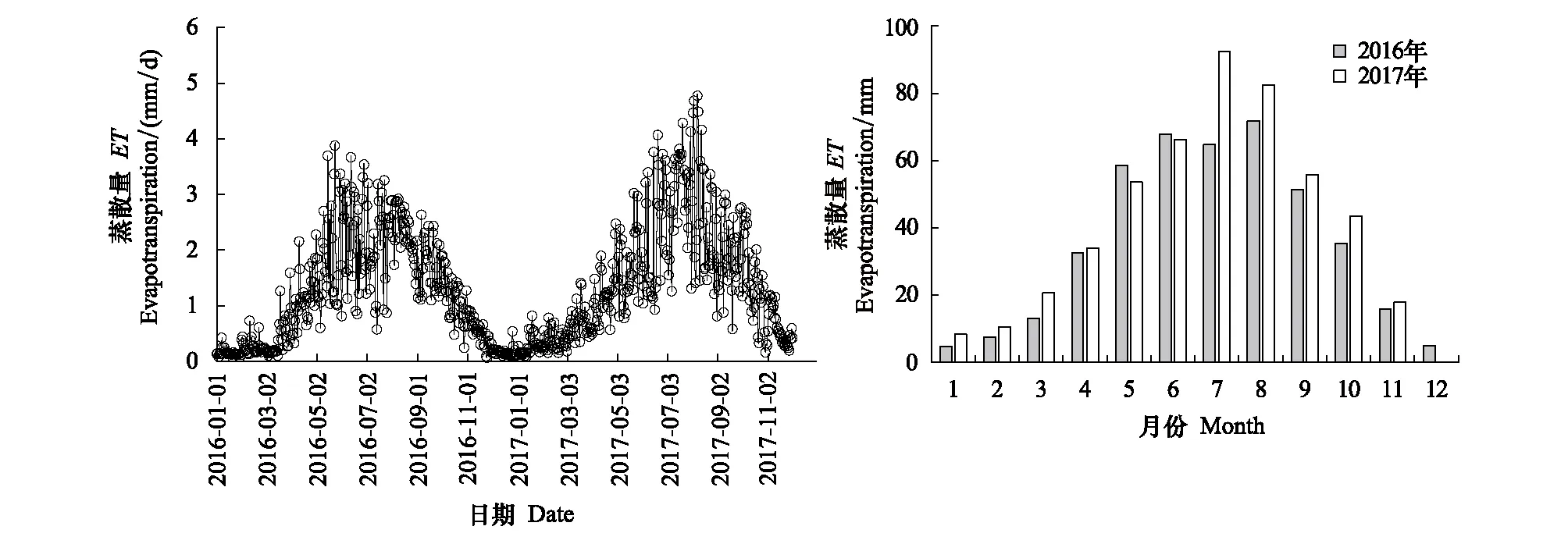
图3 2016—2017年退化草甸日蒸散量和月蒸散量的年变化Fig.3 Annual variation of daily and monthly evapotranspiration (ET) in degraded meadow for 2016 and 2017
2.3 土壤冻融与蒸散2.3.1 冻融时段的划分
参考Guo等[26]对冻融阶段的划分方法,本研究利用5 cm深土壤温度日最高和最低值变化,将全年划分为冻结期、冻融交替期、消融期3个时段:(1)土壤冻结期:土壤温度日最高值小于0℃;(2)冻融交替期:土壤温度日最高值大于0℃,而日最低值小于0℃;(3)土壤消融期:土壤温度日最低值大于0℃。为避免随机天气过程对土壤冻融阶段转变的影响,只有当连续三天满足下一阶段条件时,这三天中的第一天才作为下一阶段起始日期。为了研究植被对蒸散的影响,进一步把5—9月作为植物生长季独立划分出来。由3个时段及生长季的划分(表2)可知,2016年的冻结期和冻融交替期分别比2017年长8 d,而2017年的消融期比2016年多15 d。
2.3.2不同冻融时段蒸散量和蒸散速率
为阐明2016和2017年不同时段的ET差异,对上述划分的3个时段及生长季(表2)的ET量和ET速率进行了统计(表3)。由表可知,2016年仅冻融交替期的ET量高于2017年,而其他3个时段均低于2017年,尤其在消融期和生长季,2017年比2016年分别增加了63.1 mm和36.3 mm。

表2 2016和2017年土壤的冻结期、冻融交替期、消融期及生长季的划分
2016和2017年ET速率均在生长季达到最高值,分别为2.05 mm/d和2.29 mm/d;最低值出现在冻结期,分别为0.27 mm/d和0.33 mm/d。2016年冻融交替期的ET速率为0.97 mm/d,高于2017年的0.73 mm/d,且2016年冻融交替期的时间比2017年长8 d,因此该时段2016年ET量高于2017年,而在其他3个时段,2016年ET速率均低于2017年。

表3 2016和2017年3个时段及生长季蒸散量和蒸散速率
2.3.3不同冻融时段蒸散日变化
2016和2017年各时段的蒸散均呈明显的日变化(图4),ET的日最高值出现在13:00 h左右,而夜间维持在零附近。生长季的蒸散速率明显高于其他3个时段,而冻融交替期的蒸散速率介于消融期和冻结期之间。生长季期间,2017年蒸散速率的日最高值为0.30 mm/h,高于2016年同期的0.28 mm/h,而在土壤冻融交替时期,2017年蒸散速率的日最高值为0.11 mm/h,低于2016年同期的0.13 mm/h,在冻结期和消融期,2016年的蒸散速率均低于2017年。
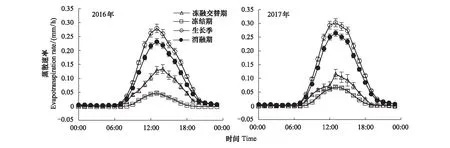
图4 2016和2017年生长季、冻结期、冻融交替期、消融期的蒸散日变化Fig.4 Diurnal variations of evapotranspiration (ET) in growing season, frozen, freeze-thaw and thawed periods for 2016 and 2017
2.4 平衡蒸散量
平衡蒸散量(ETeq),是指在没有水分限制的条件下,生态系统理论上ET的最大值[25]。为了进一步评价生态系统蒸散对水分收支的影响,我们对ETeq,ET和降水量(P)的年累计值年变化进行了比较(图5)。由图可知,2016与2017年3个变量的变化趋势相同,均在生长季出现快速增加趋势,而在其他时期相对缓慢,且ETeq的累计值高于ET和P。2016年ETeq的年累计值为626.9 mm,其中1—11月累计量为612.3 mm,高于2017年同期的573.7 mm。在植物生长季之前,2016年的ET累计值高于P,当进入生长季后,P的累计值高于ET,而2017年的ET累计值始终高于P。
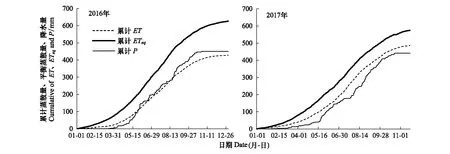
图5 2016与2017年蒸散量(ET)、降水量(P)和平衡蒸散量(ETeq)的累计值Fig.5 Cumulative evapotranspiration (ET), precipitation (P) and equilibrium evapotranspiration (ETeq) for 2016 and 2017
3 讨论
3.1 植被对蒸散的影响
蒸散包括土壤蒸发和植被蒸腾,生长季中的植被生长状况直接影响植被蒸腾和土壤蒸发的变化[4]。由植物生长旺季(7月)的地上生物量(above-ground biomass,AGB)和叶面积指数(LAI)观测数据可知,2016年的AGB为(192.7±33.1) g/m2,明显高于2017年同期的(114.6±18.5) g/m2;且2016年LAI为(1.2±0.1) m2/m2,同样高于2017年同期的(0.8±0.1) m2/m2。然而,2017年生长季中的ET却明显高于2016年,说明地上植被量的降低反而增加了该退化高寒草甸生态系统的蒸散,分析其可能的原因之一:Hu等[27]曾在研究中发现,在植被LAI较低情况下,ET主要由土壤蒸发控制。因此在该退化草甸中,地上生物量减少使植被盖度明显降低,土壤裸露面积增加,提高了土壤蒸发量,从而使生态系统ET增加,这在一些研究中也得到证实[28-29]。Zhang等[28]利用模型对三江源区退化草甸和人工草地能量分配的研究结果发现,退化草甸消耗的潜热高于人工草地,主要原因是退化草甸的土壤蒸发远高于人工草地。
ET/P是描述水分收支的重要参数之一[7]。通过与不同植被类型生态系统的蒸散结果比较可知(表4),本研究退化高寒草甸生态系统两年ET/P分别为0.95和1.09,该值在已报道的草地生态系统ET/P的范围(40%—150%)之间[34],但明显高于Gu等报道的未退化海北高寒草甸的0.60[6]和Li等报道的日本温带草地的0.66[31],但低于内蒙古退化草地的1.4[30]。本结果意味着植被退化可能会加剧该高寒草甸的水分散失,降低生态系统的水源涵养能力。
3.2 环境因子对蒸散的影响3.2.1 辐射与温度对蒸散的影响
太阳辐射能(Rs)是驱动陆地生态系统蒸散的主要动力[6],由于青藏高原海拔高,因此接收的太阳总辐射远高于低海拔地区。净辐射(Rn)是供给生态系统蒸散的有效能量,在不受水分限制条件下,ET主要受Rn控制[28]。2016和2017年ET随Rn的变化趋势相似(图6),当Rn<4 MJ m-2d-1时,由于土壤处于冻结状态,ET维持在相对较低水平,且基本不受Rn变化的影响;当Rn≥ 4 MJ m-2d-1时,随着土壤解冻的开始,ET随Rn的增加而呈现直线上升的趋势,该结果与张立锋等[28]报道的相一致。但在相同Rn条件下,2017年ET值高于2016年,尤其在生长季Rn> 10 MJ m-2d-1后,两年差异更明显,且2017年拟合直线的斜率高于2016年,说明2017年ET随Rn上升而增加的速率更快,即ET对Rn变化的响应更敏感。
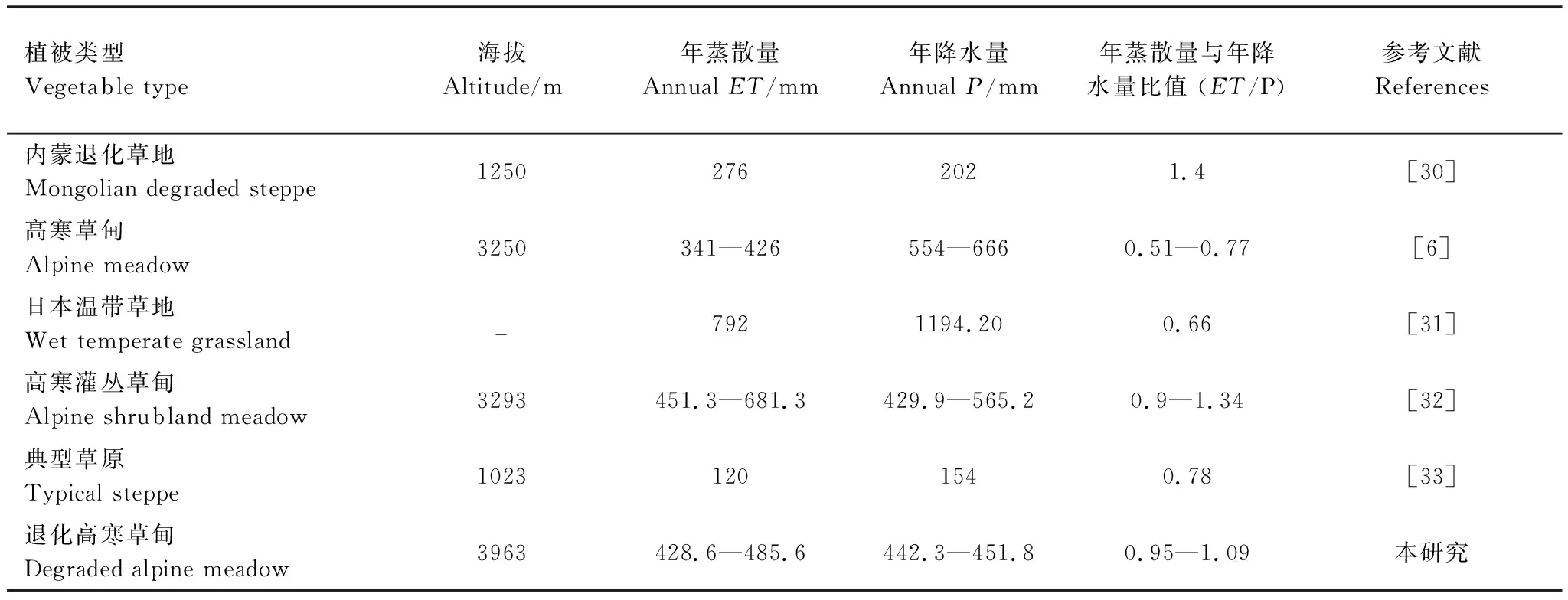
表4 不同草地类型蒸散量和降水量的比较
研究期间,该生态系统接收的太阳辐射年总量高达6594.32 MJ/m2,但由于高原上相对较低的大气逆辐射和相对较高的反照率,反而导致接收的Rn较低[35]。本研究两年Rn/Rs平均值0.43,接近于已报道的海北高寒草甸的0.44[6],明显低于全球平均值0.61和一些已经报道的草地系统[36]。因此该研究地驱动蒸散的有效能Rn反而比低海拔地区低[36],从而导致生态系统的蒸散对Rn的响应异常敏感。另外,由于2017年高寒草甸的AGB和LAI低于2016年,使土壤蒸发在蒸散中所占的比例增加,Zhang等[29]指出三江源区高寒草甸的退化加剧了生态系统的蒸散量,这可能是导致2017年ET对Rn变化的响应比2016年更为敏感的原因之一,此外相关环境因子的影响尚有待于进一步研究。
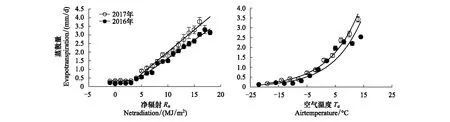
图6 2016和2017年退化草甸蒸散量(ET)与净辐射(Rn)和空气温度(Ta)的关系Fig.6 Relationships between evapotranspiration (ET) and net radiation (Rn), air temperature (Ta) in degraded meadow for 2016 and 2017 ET与 Rn: 2016年y=0.213x-0.49 (R2=0.99) (P<0.01); 2017年y=0.255x-0.53 (R2=0.98)(P<0.01); ET与Ta: 2016年 y=0.84e0.098x (R2=0.94) (P<0.01); 2017年y=1.09e0.094x (R2=0.98) (P<0.01)
温度不仅控制植物生长,也是影响生态系统蒸散的重要因子,气温的升高可增加边界层的水热交换,从而提高蒸发的速率[37]。本研究ET均随Ta升高而成指数上升趋势(图6),该结果与陈小平等[37]报道的草地生态系统相似。需要指出的是,许多平原地区的长期研究表明空气温度与蒸散相关度不高[38],而三江源区高寒草甸生态系统与之相反,主要是由于其长期处于低温环境,因此ET对温度的响应更为敏感,ET随温度的变化速率更快。另外,由于2017年平均气温为-0.41℃低于2016年的-0.35℃,尤其在生长季2017年平均气温为6.8℃,明显低于2016年的7.2℃(图2),这可能是导致2017年ET随温度升高而增加的速率略高于2016年的原因之一。
3.2.2饱和水气压差与土壤水分对蒸散的影响
饱和水汽压差(VPD)是反映空气对水分的需求能力[39],VPD通过影响冠层导度gc,从而影响生态系统的蒸散[31]。本研究ET均随VPD的增加而呈直线上升趋势(图7),这与已报道的一些研究结果类似[39-40]。然而,2017年拟合直线的斜率为2.64明显高于2016年的1.46,说明2017年ET随VPD的增加而上升的速率更快,进而导致在相同VPD条件下,2017年的蒸散量高于2016。低的VPD通常可降低大气蒸散需求,而高VPD表征空气干燥,可促进土壤蒸发和植被蒸腾[5]。本研究VPD变化范围在0.05-1.69 kPa之间,明显低于已报道的其他草地生态系统2-5 kPa的变动范围[4,41],这可能是导致本研究ET对VPD变化响应敏感的原因之一。

图7 2016和2017年退化草甸蒸散量(ET)与饱和水汽压差(VPD)和 5 cm土壤含水量(SWC5)的关系Fig.7 Relationships between evapotranspiration (ET) and vapor pressure deficit (VPD), soil water content at 5 cm depth (SWC5) in degraded meadow for 2016 and 2017 ET与VPD: 2016年: y=1.46x+0.36 (R2=0.82) (P<0.01); 2017年y=2.64x+0.09 (R2=0.92) (P<0.01); ET与SWC5: 2016年 y=31.73x2+18.06x-0.75 (R2=0.83) (P<0.01); 2017年y=-55.16x2+26.36x-1.03 (R2=0.84) (P<0.01)
土壤水分是土壤蒸发和植物蒸腾的直接来源,是影响陆地生态系统ET的关键因子[28]。高寒草甸的根系主要分布在0—10 cm深度的表层土壤[5],因此生态系统的ET与5 cm的土壤含水量(SWC5)密切相关。当SWC5低于0.25 m3/m3,两年ET随SWC5的升高而增加,当高于0.25 m3/m3时,ET随SWC5的升高呈平稳(2016年)或下降(2017年)趋势(图7),这与已报道的一些草地研究结果类似[28,42]。通常,当土壤水分受限时,ET随土壤湿度的升高而增加,而当土壤水分不受限时,ET随土壤水分升高的响应不敏感或呈下降趋势[28]。由于2017年SWC5最高值出现在9月(图2),而该时期驱动蒸散的净辐射和温度远低于7-8月的值(图2),可能是导致2017年的蒸散随SWC5的上升而降低的原因。
3.2.3环境因子综合作用对蒸散的影响
为深入探讨环境因子对ET的影响,对ET与主要环境因子(Rn、Ta、SWC5、VPD)进行逐步回归分析。由相关系数(表5)可知,该生态系统ET与上述环境因子均呈极显著相关,其中Rn和Ta相关性最高。2016和2017年回归方程分别为:ET=0.639Rn+0.291Ta+0.092SWC5-0.046 (R2=0.866,P=0.001);ET=0.532Rn+0.439Ta+0.353 (R2=0.787,P=0.000),表明2016年Rn对ET的影响最大,其次是Ta和SWC5;而2017年ET主要受Rn和Ta的影响。由此可见,该生态系统的ET变化主要受Rn和Ta的控制。
3.3 冠层导度gc和解耦系数Ω对蒸散的影响
冠层导度(gc)是影响蒸散的重要因子,其变化受LAI、SWC、VPD等环境因子的影响,通常ET随gc的升高而增加[43]。为评价gc对ET的影响,本研究计算了4个时段晴天条件下[30](晴空指数大于0.7)的gc日变化(图8),可以得出生长季中的gc最高,其次为消融期,冻融交替期,而冻结期的gc最低,该结果与已报道的加拿大的温带草地、海北高寒草甸等生态系统变化趋势一致[4,6],且与不同时段的蒸散速率变化相一致(图4),主要是由于高的ET一般与高的gc相关,生长季植被生长茂盛以及较高的土壤湿度,进而导致gc升高,蒸散速率加快[44]。然而,2017年生长季gc的平均值为10.3 mm/s,明显高于2016年的8.2 mm/s,这是导致2017年生长季ET高于2016年的原因之一。两年冻融交替期和冻结期gc的平均值小于3 mm/s,且2016年与2017年相差不大。

表5 2016和2017年退化草甸ET与主要环境因子的Pearson相关系数
两年的gc日变化趋势相同,生长季和消融期gc最高值出现在正午前后,而另外两个时段gc最高值出现在8:00—11:00之间,该结果与Wever等[4]和Zhang等[29]研究结果一致。在冻结期和冻融交替期,由于植物凋萎,地面有效辐射加强,温度和VPD较低,这可能是导致gc在上午出现高值的原因[28];而在降水丰沛的生长季和消融期,由于生长季水分不受限制,温度和VPD相对较高,日间gc随VPD的增大而增强,但当VPD过大时,导致气孔关闭,gc降低,因此gc的最大值出现在正午前后[4,44],这也和ET的最大值出现的时间相吻合。
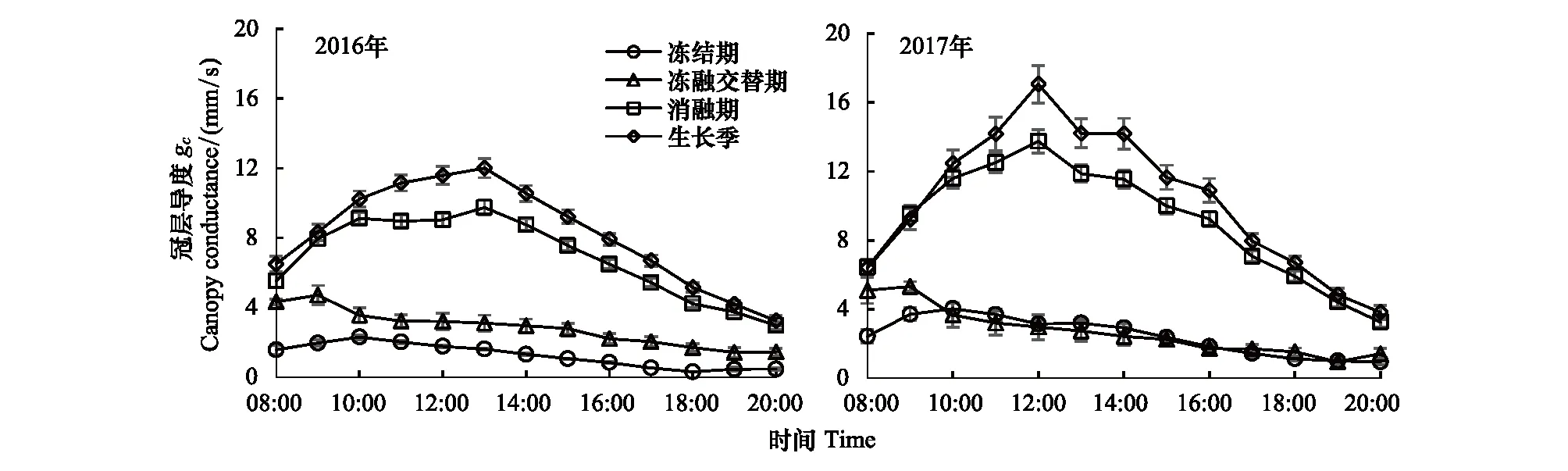
图8 2016年和2017年晴天条件下退化草甸冻结期、冻融交替期、消融期、生长季的冠层导度(gc)日变化Fig.8 Diurnal pattern of canopy conductance (gc) on clear days in growing season, frozen, freeze-thaw and thawed periods in degraded meadow for 2016 and 2017
解耦系数Ω是评价地表与大气之间水热交换耦合状况的参数[45],其值在0—1之间变化,当Ω越接近0时,表明ET主要受VPD控制;Ω越接近1,表明ET主要受Rn主导[4,40]。两年Ω的变化趋势相同,最高值出现在生长季和消融期(图9),2016和2017年生长季Ω的平均值分别为0.55和0.60,而冻结期和冻融交替期的Ω明显低于生长季和消融期,这与已报道的诸多研究结果相同[6,17,28]。该结果说明生长季和消融期的ET主要受Rn影响,而冻结期和冻融交替期的Rn并不是驱动ET的主要因子。另外,2017年生长季和消融期的Ω均高于2016年,可见Rn对2017年ET的影响大于2016年。
年Ω日变化均呈现上午高于下午的变化趋势,该变化与Wever等[4]和Hao等[40]报道的草地结果相同,说明该生态系统Rn对午前ET的影响相对较大,而VPD对午后ET的影响相对较大。

图9 2016年和2017年晴天条件下退化草甸冻结期、冻融交替期、消融期、生长季的解耦系数(Ω)日变化Fig.9 Diurnal pattern of decoupling coefficient (Ω) on clear days in growing season, frozen, freeze-thaw and thawed periods in degraded meadow for 2016 and 2017

图10 2016年和2017年退化草甸晴天条件下日均Priestley-Taylor系数(ET/ETeq)与冠层导度(gc)之间的关系Fig.10 Relationship between the daily average Priestley-Taylor coefficient (α=ET / ETeq) and canopy conductance (gc) on clear days in degraded meadow for 2016 and 20172016年:y=0.2696ln(x)+0.2007 (R2=0.87) (P<0.01); 2017年:y=0.294ln(x)+0.2554 (R2=0.87) (P<0.01)
Priestley-Taylor系数(α=ET/ETeq)为蒸散与平衡蒸散之比,其能够反映土壤中可利用水分对蒸散的影响,通常水分供应充足的生态系统ET/ETeq高于水分受限的生态系统[46]。晴天条件下,两年ET/ETeq与gc均呈现对数增长趋势(图10),这与之前报道的很多结果相一致[5,29,32], 这是由于生长季的gc远高于冻结期和冻融交替期(图8),而相对较高的gc导致生态系统更多的ET[6]。另外,2017年ET/ETeq值高于2016年,其中2016和2017年生长季的ET/ETeq分别为0.78和0.93,明显高于受水分限制的半干旱草地生态系统[30,32],但与水分充足的森林生态系统相当[47]。结果说明该生态系统ET受水分限制相对较小,且2017年生态系统受到的水分限制相对小于2016年,进而导致2016年ET低于2017年。
3.4 冻融变化对蒸散的影响
2016与2017年土壤冻融交替期、冻结期及消融期的长短不同(表2),冻融交替期分别为21 d和13 d,冻结期分别为140 d和132 d,而消融期分别为205 d和220 d。该结果说明2016年的冻结期和冻融交替期比2017年的时间长,而消融期则比2017年短15 d,进而导致2017年消融期(包括生长季)的ET比2016年高63.1 mm(表4);而2017年冻融交替期比2016年缩短8 d,其ET比 2016年低19.9 mm。
由以上结果可知,冻融时间的变化可引起地气之间能量和水分交换状况的改变,从而导致生态系统蒸散的变化[26]。通常土壤冻结可降低土壤蒸发,减少土壤水分的散失,当土壤消融后,相对较高的土壤含水量及相对较低的地表反照率(使土壤吸收更多的太阳辐射),以及辐射和温度的上升,进而加剧了土壤蒸发量[48]。有研究表明,温度(特别是冬天)的升高,可能导致青藏高原土壤消融期提前、冻结推迟以及高原冻结天数持续下降[49]。因此在全球气候变化背景下,三江源区土壤冻融时间将会发生改变,从而对蒸散变化产生深刻影响,而未来气候变暖可能会加剧该退化高寒草甸生态系统系统的蒸散量,降低其水源涵养能力。
4 结论
三江源区退化高寒草甸生态系统的年蒸散量可达降水量的95%以上,且存在明显的年际间差异,植被生物量相对较高的年份,其生态系统的蒸散量反而低于生物量相对较低的年份,由此可推测该区域高寒草甸的退化可能将导致其生态系统水源涵养功能的降低。同时,土壤冻融时间的改变对其生态系统的蒸散产生了重要影响,土壤冻结期的缩短将引起蒸散量的增加。由于影响该生态系统蒸散的最主要环境因子是净辐射和温度,因此在全球气候变暖的背景下,三江源区退化高寒草甸生态系统的蒸散可能出现增加趋势,进而对该区域的水分收支产生深刻的影响。
